

Optimized Method for the Identification of Candidate Genes and Molecular Maker Development Related to Drought Tolerance in Oil Palm (Elaeis guineensis Jacq.)
 ,
,  and
and 
Abstract
:1. Introduction
2. Results
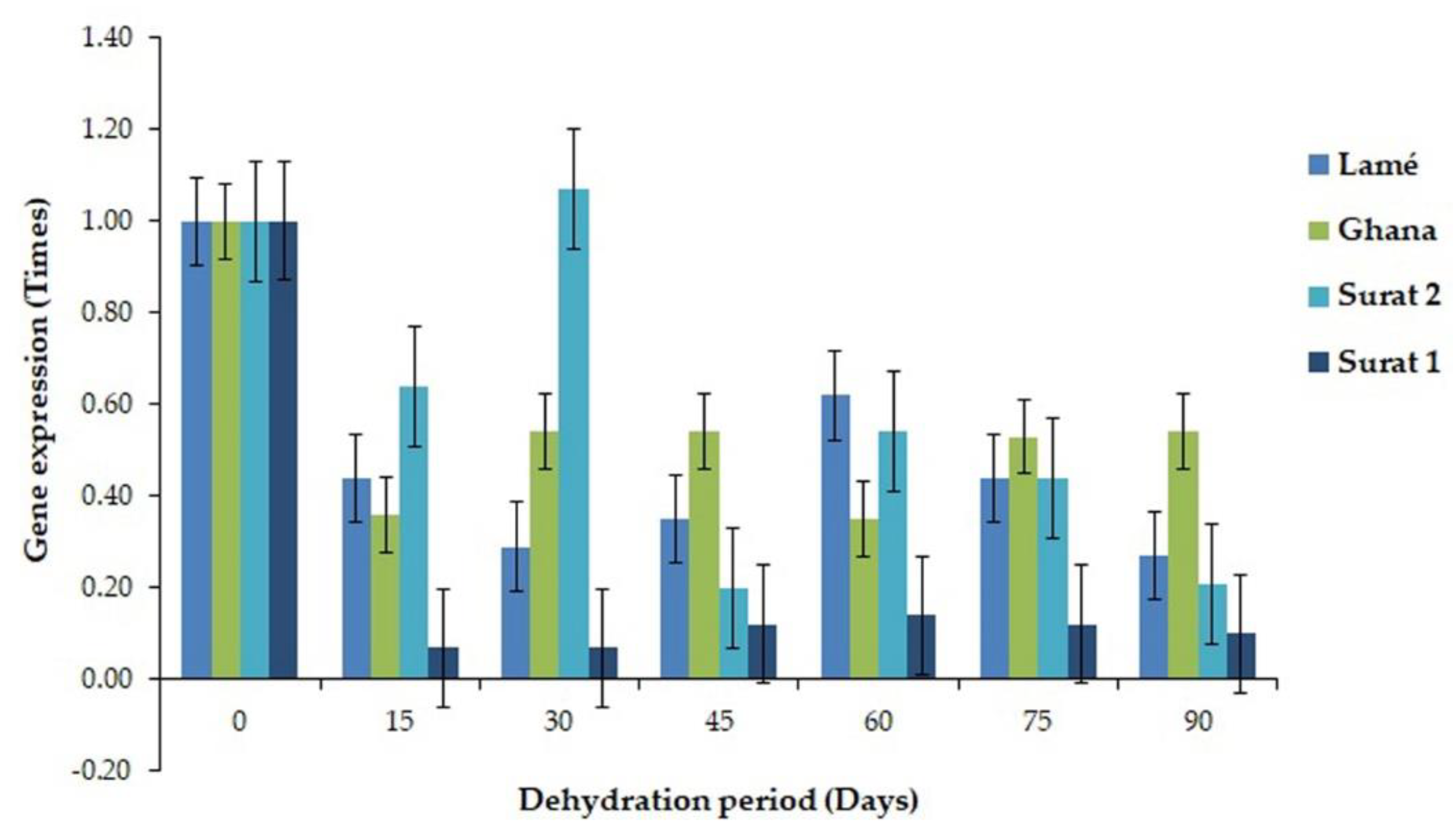
2.1. Differential Display Reverse Transcriptase PCR (DDRT-PCR) and RT-PCR
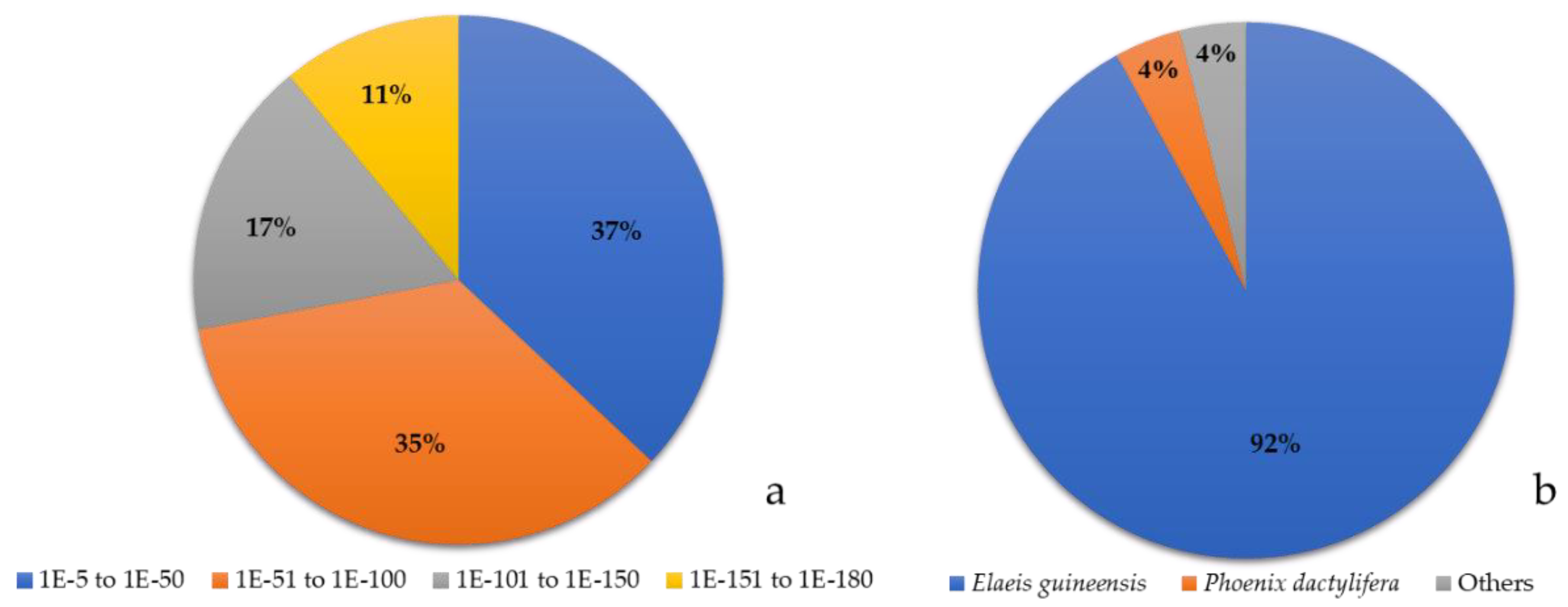
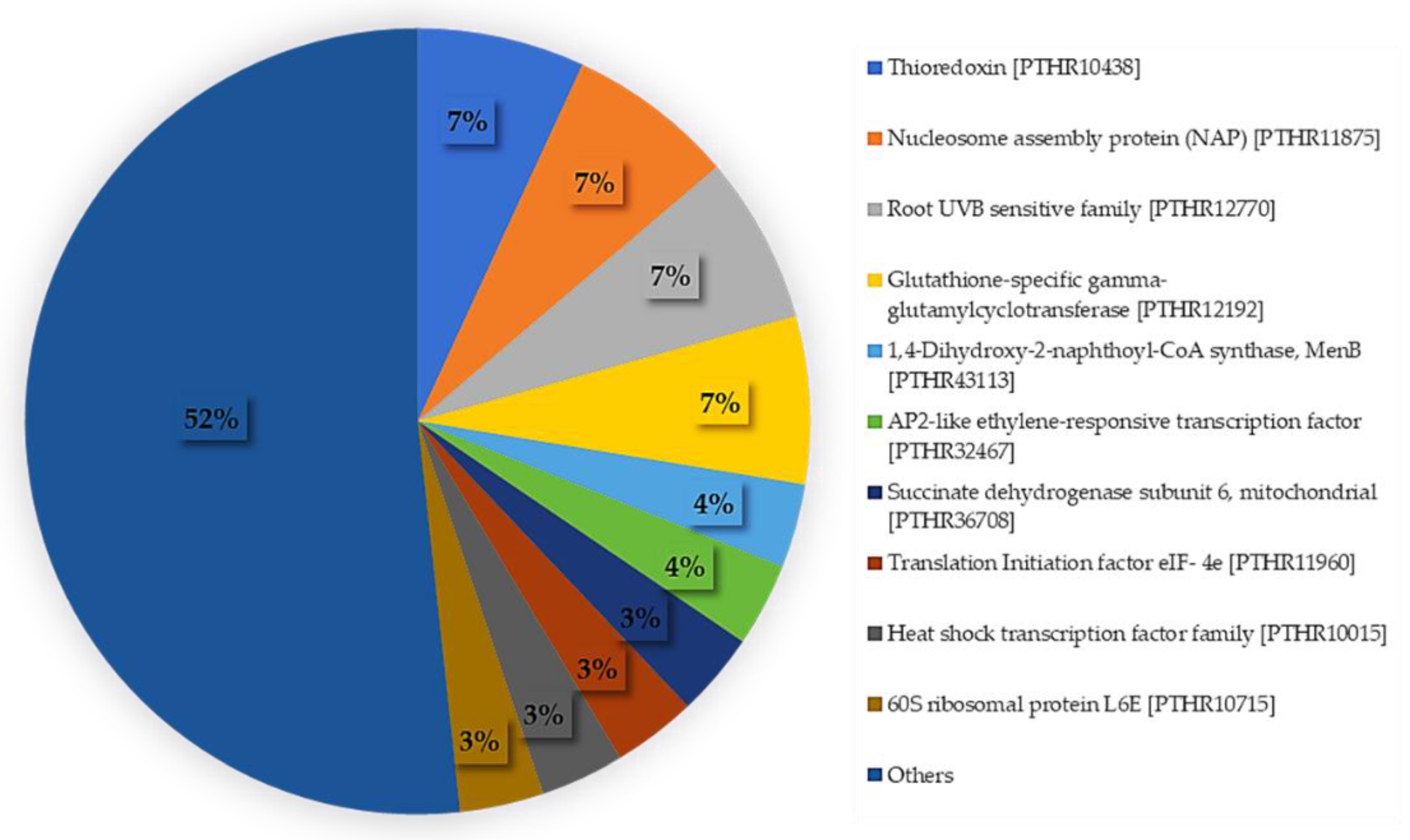
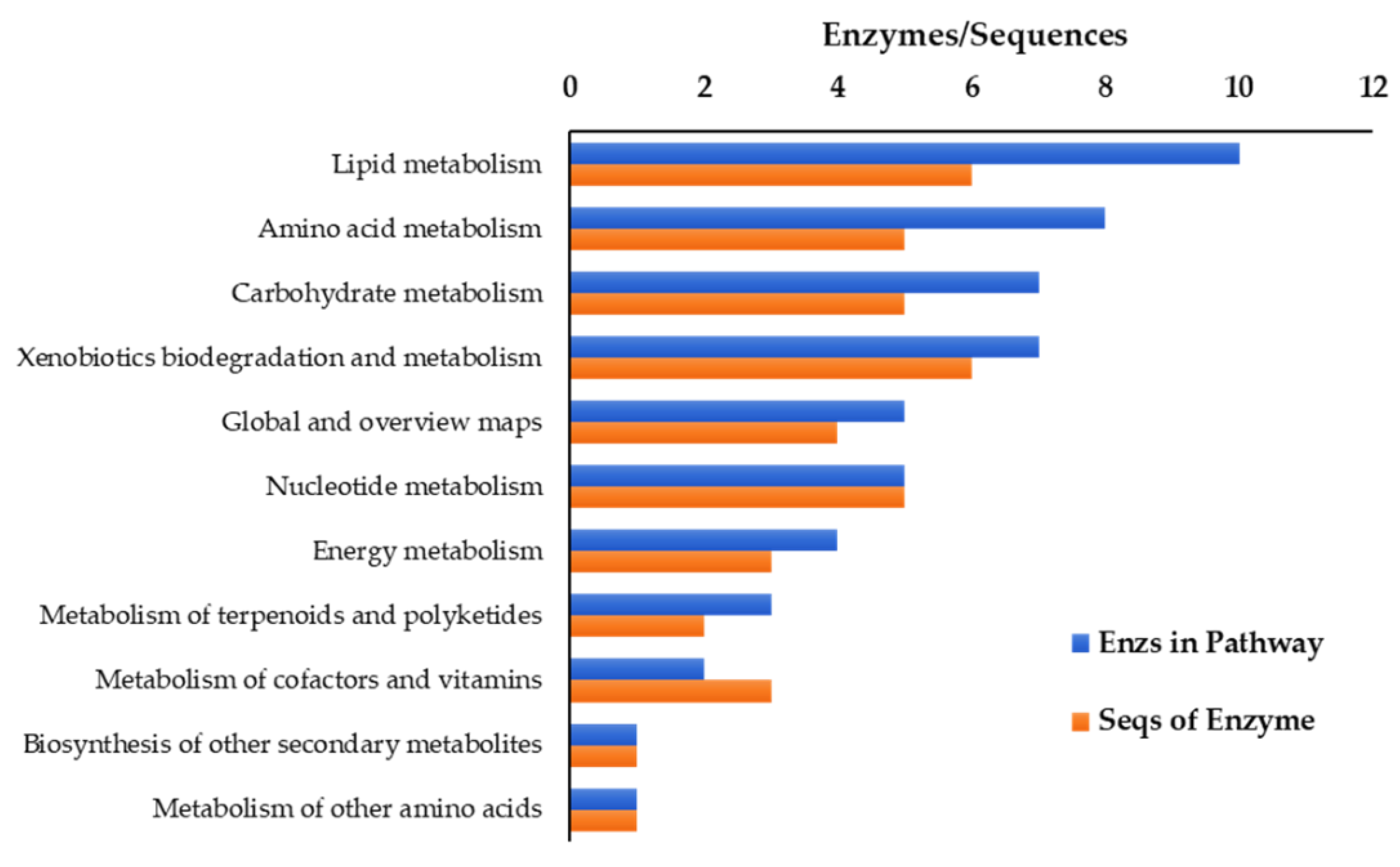
2.2. 454 Sequencings, Transcriptome Assembly, and Annotation
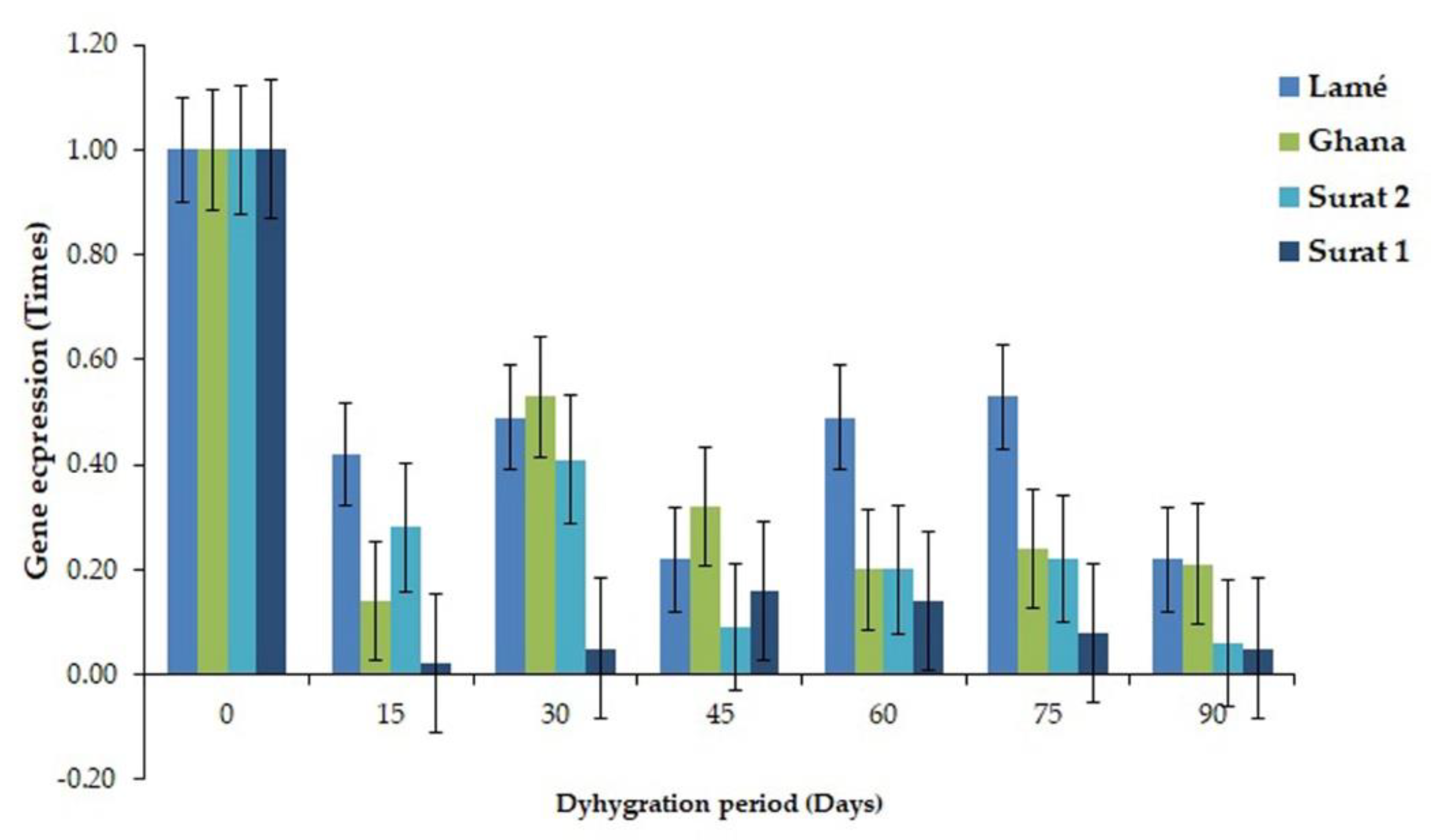
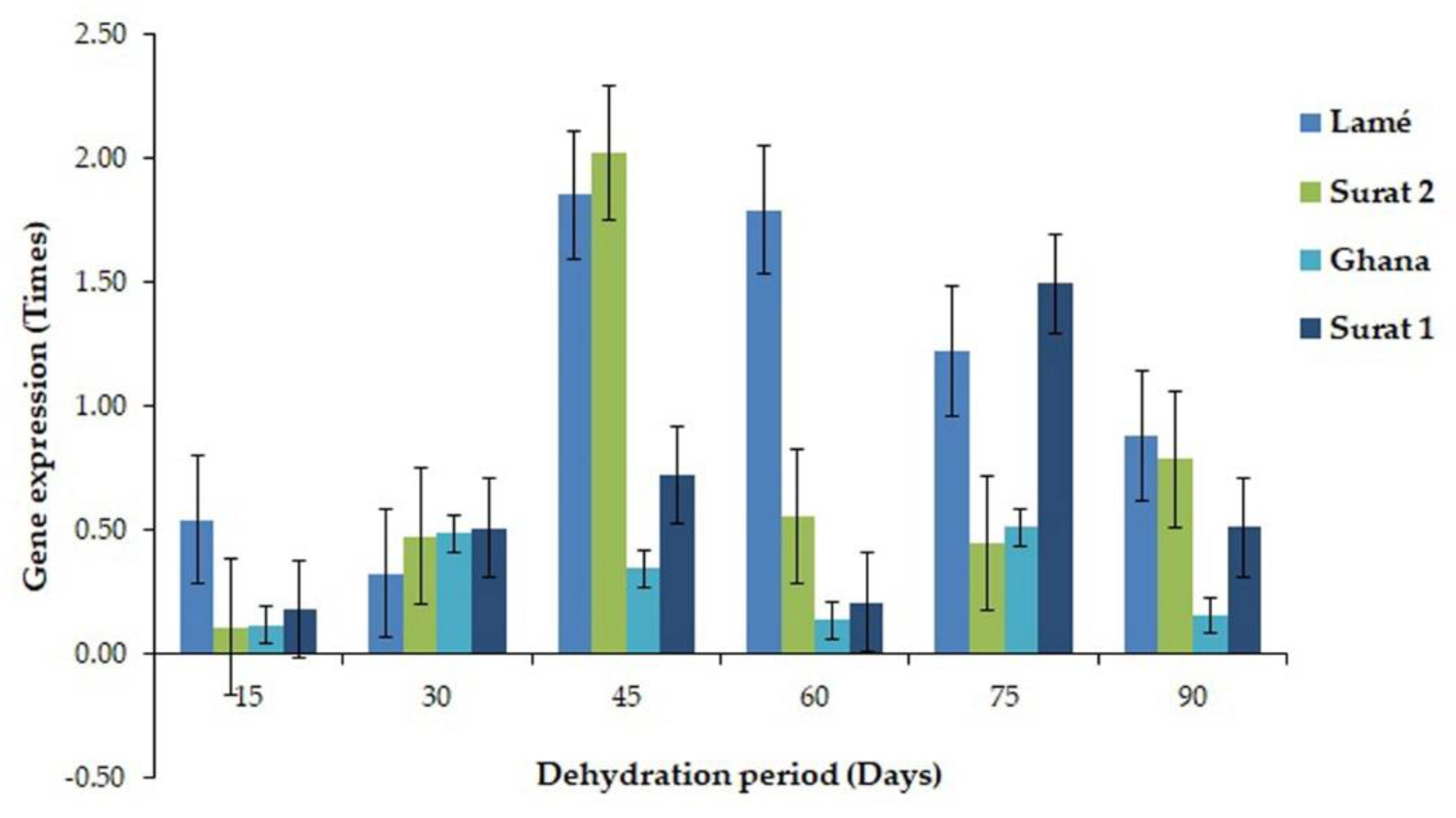
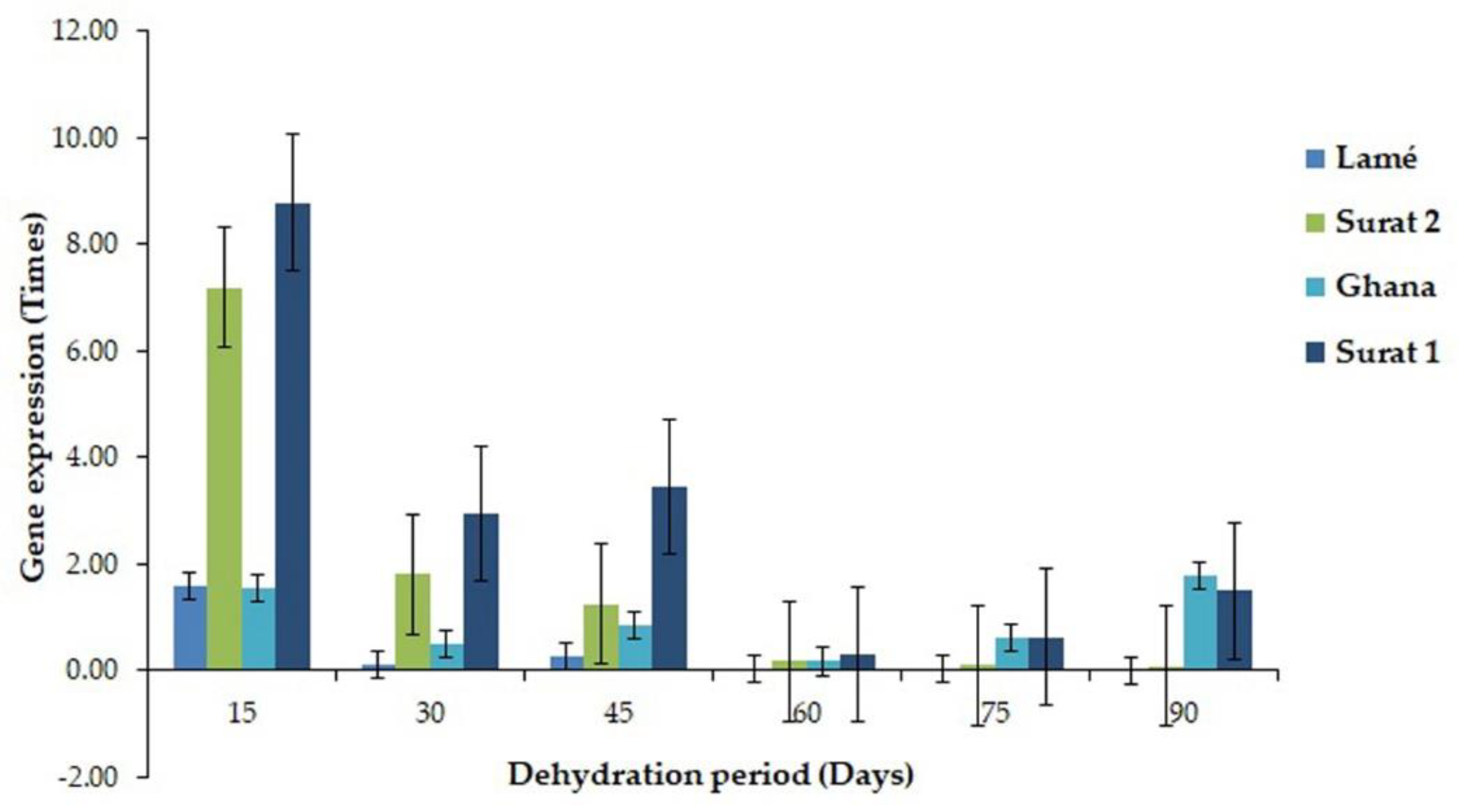
2.3. Comparison of the Expressed Gene Associated with the Drought Tolerance between DDRT-RT and Pyrosequencing and Real-Time PCR
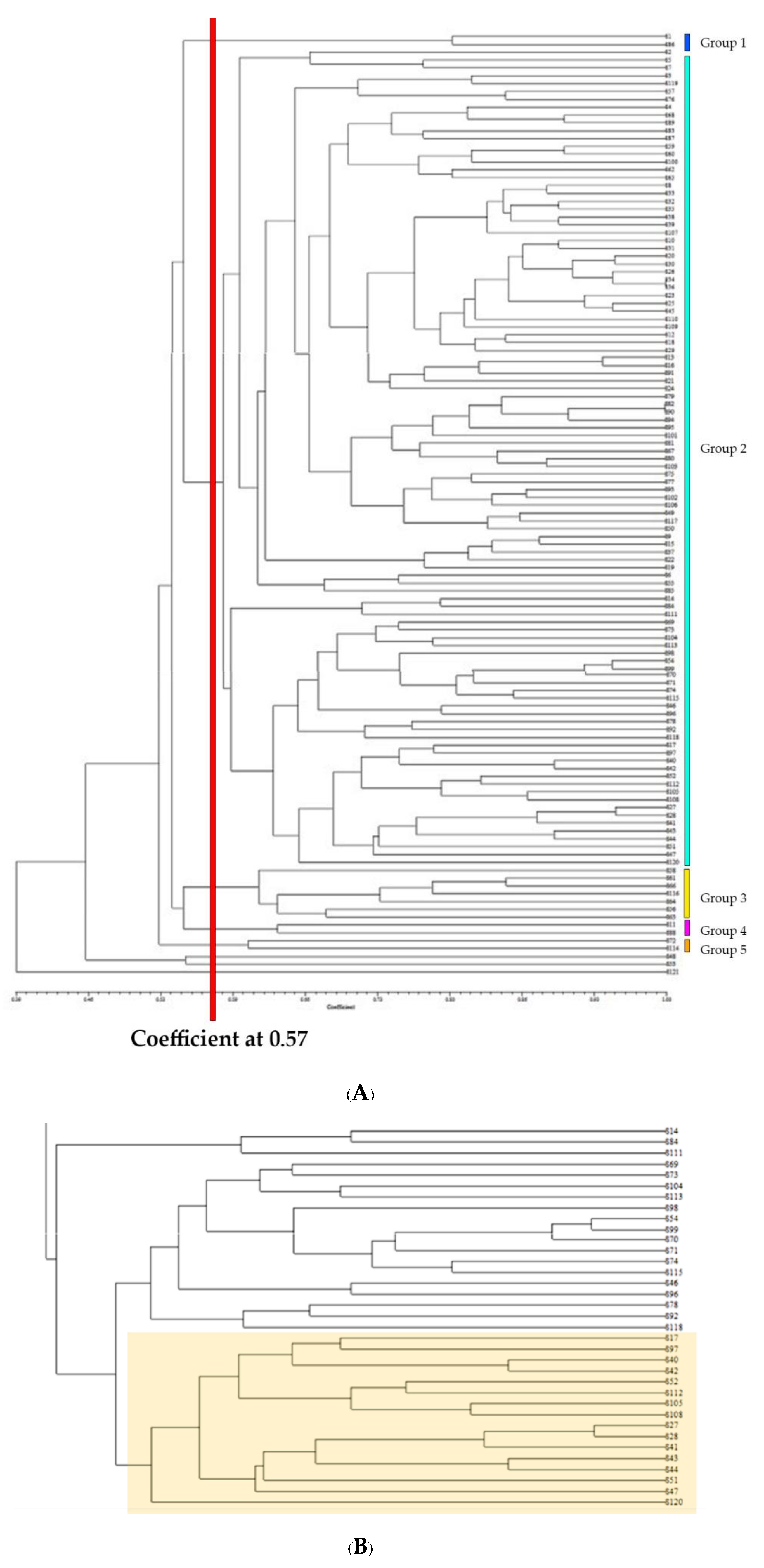
2.4. Molecular Marker Development
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Dehydrated Condition
4.2. Differential Display Reverse Transcription Polymerase Chain Reaction (DDRT-PCR)
4.3. RNA Isolation, Transcriptome Analysis, and Sequence Annotation
4.4. Comparison of the Expressed Gene Associated with Drought Tolerance from Transcriptome and DDRT-RT and Real-Time PCR
4.5. Single Sequence Repeat (SSR) Molecular Marker Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 4th ed.; Blackwell Science Ltd.: Oxford, UK, 2003; p. 10. [Google Scholar]
- Bray, E.A. Genes commonly regulated by water-deficit stress in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 2331–2341. [Google Scholar] [CrossRef] [PubMed]
- Caliman, J.P.; Southworth, A. Effect of drought and haze on the performance of oil palm. In Proceedings of the IOPRI International Oil Palm Conference: Commodity of the Past, Today and the Future, Sheraton Nusa Indah Hotel, Bali, Indonesia, 23–25 September 1998. [Google Scholar]
- Ahmadi, M.; Heravan, E.M.; Sadeghian, S.Y.; Mesbah, M.; Darvish, F. Drought tolerance variability in S1 pollinator lines developed from a sugar beet open population. Euphytica 2011, 178, 339–349. [Google Scholar] [CrossRef]
- Schafleitner, R.; Rosales, R.O.G.; Gaudin, A.; Aliaga, C.A.A.; Martinez, G.N.; Marca, L.R.T.; Bolivar, L.A.; Delgado, F.M.; Simon, R.; Bonierbale, M. Capturing candidate drought tolerance traits in two native Andean potato clones by transcription profiling of field grown plants under water stress. Plant Physiol. Biochem. 2007, 45, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Gohin, M.; Bobe, J.; Chesnel, F. Comparative transcriptomic analysis of follicle-enclosed oocyte maturational and developmental competence acquisition in two non-mammalian vertebrates. BMC Genom. 2010, 11, 18. [Google Scholar] [CrossRef]
- Emboule, L.; Daigle, F.; Meyer, D.; Mari, B.; Pinarello, V.; Sheikboudou, C.; Magnone, V.; Frutos, R.; Viari, A.; Barbry, P.; et al. Innovative approach for transcriptomic analysis of obligate intracellular pathogen: Selective capture of transcribed sequences of Ehrlichia ruminantium. BMC Mol. Biol. 2009, 10, 111. [Google Scholar] [CrossRef]
- Lu, F.H.; Cho, M.C.; Park, Y.J. Transcriptome profiling and molecular marker discovery in red pepper Capsicum annuum L. TF68. Mol. Biol. Rep. 2012, 39, 3327–3335. [Google Scholar] [CrossRef]
- Sun, C.; Li, Y.; Wu, Q.; Luo, H.; Sun, Y.; Song, J.; Lui, E.; Chen, S. De novo sequencing and analysis of the American ginseng root transcriptome using a GS FLX Titanium platform to discover putative genes involved in ginsenoside biosynthesis. BMC Genom. 2010, 11, 262. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, F.; Liu, R.; Feng, J.; Li, H. De novo sequencing and analysis of root transcriptome using 454 pyrosequencing to discover putative genes associated with drought tolerance in Ammopiptanthus mongolicus. BMC Genom. 2012, 13, 266. [Google Scholar] [CrossRef]
- Cheung, F.; Win, J.; Lang, J.; Hamilton, J.; Vuong, H.; Leach, J.; Kamoun, S.; Levesque, A.C.; Tisserat, N.; Buell, C.R. Analysis of the Pythium ultimum transcriptome using Sanger and pyrosequencing approaches. BMC Genom. 2008, 9, 542. [Google Scholar] [CrossRef]
- Alves, J.D.; VanToai, T.; Kaya, N. Differential display: A novel PCR—Based method for gene isolation and cloning. Rev. Bras. Fisiol. Vegetal. 1998, 10, 161–164. [Google Scholar]
- Rodríguez, M.; Canales, E.; Borrás-Hidalgo, O. Molecular aspects of abiotic stress in plants. Biotecnol. Apl. 2005, 22, 1–10. [Google Scholar]
- Hadi, S.M.; Ibrahim, K.M.; Yousif, S.I. Differential expression for genes in response to drought and salinity in Ruta graveolens Plantlets. Jordan J. Biol. Sci. 2019, 12, 203–207. [Google Scholar]
- Srivastava, S.; Gupta, S.M. Role of differentially expressed ripening related genes and promoters in banana fruit: Identified by mRNA DDRT-PCR. Res. Environ. Life Sci. 2008, 1, 81–90. [Google Scholar]
- Benito, E.P.; Prins, T.; Kan, J.A.V. Application of differential display RT-PCR to the analysis of gene expression in a plant-fungus interaction. Plant Mol. Biol. 1996, 32, 947–957. [Google Scholar] [CrossRef]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, X.; Guo, X.; Tang, G.; Sun, F.; Liu, S.; Xi, Y. Transcriptional and physiological data reveal the dehydration memory behavior in switchgrass (Panicum virgatum L.). Biotechnol. Biofuels 2018, 11, 91. [Google Scholar] [CrossRef]
- Zhang, P.; Fan, Y.; Sun, X.; Chen, L.; Terzaghi, W.; Bucher, E.; Li, L.; Dai, M. A large-scale circular RNA profiling reveals universal molecular mechanisms responsive to drought stress in maize and Arabidopsis. Plant J. 2019, 98, 697–713. [Google Scholar] [CrossRef]
- Wang, L.; Lee, M.; Ye, B.; Yue, G.H. Genes, pathways and networks responding to drought stress in oil palm roots. Sci. Rep. 2020, 10, 21303. [Google Scholar] [CrossRef]
- Lee, F.C.; Yeap, W.C.; Appleton, D.R.; Ho, C.L.; Kulaveerasingam, H. Identification of drought responsive Elaeis guineensis WRKY transcription factors with sensitivity to other abiotic stresses and hormone treatments. BMC Genom. 2022, 23, 2–17. [Google Scholar] [CrossRef]
- Xu, W.; Li, Y.; Cheng, Z.; Xia, G.; Wang, M. A wheat histone variant gene TaH2A.7 enhances drought tolerance and promotes stomatal closure in Arabidopsis. Plant Cell Rep. 2016, 35, 1853–1862. [Google Scholar]
- Simova-Stoilova, L.; Vaseva, I.; Grigorova, B.; Demirevska, K.; Feller, U. Proteolytic activity and cysteine protease expression in wheat leaves under severe soil drought and recovery. Plant Physiol. Biochem. 2010, 48, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zou, Y.; Hu, J.; Ding, Y. Genome-wide analysis of the rice PPR gene family and their expression profiles under different stress treatments. BMC Genom. 2018, 19, 720. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.C.; Mei, C.; Liang, S.; Yu, Y.T.; Lu, K.; Wu, Z.; Wang, X.F.; Zhang, D.F. Crucial roles of the pentatricopeptide repeat protein SOAR1 in Arabidopsis response to drought, salt and cold stresses. Plant Mol. Biol. 2015, 88, 369–385. [Google Scholar] [CrossRef]
- Su, H.G.; Li, B.; Song, X.Y.; Ma, J.; Chen, J.; Zhou, Y.B.; Min, D.H.; Xu, Z.S.; Ma, Y.Z. Genome-wide analysis of the DYW subgroup PPR gene family and identification of GmPPR4 responses to drought stress. Int. J. Mol. Sci. 2019, 20, 5667. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.; Prasad, A.M.; Srinivasan, R. Pentatricopeptide repeat proteins and their emerging roles in plants. Plant Physiol. Biochem. 2007, 45, 521–534. [Google Scholar] [CrossRef]
- Chary, S.N.; Hicks, G.R.; Choi, Y.G.; Carter, D.; Raikhel, N.V. Trehalose-6-phosphate synthase/phosphatase regulates cell shape and plant architecture in Arabidopsis. Plant Physiol. 2008, 146, 97–107. [Google Scholar] [CrossRef]
- Lin, Q.; Yang, J.; Wang, Q.; Zhu, H.; Chen, Z.; Dao, Y.; Wang, K. Overexpression of the trehalose-6-phosphate phosphatase family gene AtTPPF improves the drought tolerance of Arabidopsis thaliana. BMC Plant Biol. 2019, 19, 381. [Google Scholar] [CrossRef]
- Lin, Q.; Wang, S.; Dao, Y.; Wang, J.; Wang, K. Arabidopsis thaliana trehalose-6-phosphate phosphatase gene TPPI enhances drought tolerance by regulating stomatal apertures. J. Exp. Bot. 2020, 71, 4285–4297. [Google Scholar] [CrossRef]
- Bajsa, J.; Pan, Z.; Duke, S.O. Serine/threonine protein phosphatases multi-purpose enzymes in control of defense mechanisms. Plant Signal. Behav. 2011, 6, 1921–1925. [Google Scholar] [CrossRef]
- Castilhos, G.; Lazzarotto, F.; Spagnolo-Fonini, L.; Zanettini, B.M.H.; Margis-Pinheiro, M. Possible roles of basic helixloop-helix transcription factors in adaptation to drought. Plant Sci. 2014, 223, 1–7. [Google Scholar] [CrossRef]
- Nuanlaong, S. Gene Identification Involved in Drought Tolerance in Oil Palm by Diferent Display RT-PCR (DDRT-PCR). Master’s Thesis, Walailak University, Nakhonsithammarat, Thailand, 12 November 2014. [Google Scholar]
- Augustine, S.M.; Narayan, J.A.; Syamaladevi, D.P.; Appunu, C.; Chakravarthi, M.; Ravichandran, V.; Tuteja, N.; Subramonian, N. Overexpression of EaDREB2 and pyramiding of EaDREB2 with the pea DNA helicase gene (PDH45) enhance drought and salinity tolerance in sugarcane (Saccharum spp. hybrid). Plant Cell Rep. 2015, 34, 247–263. [Google Scholar] [CrossRef]
- Manjulatha, M.; Sreevathsa, R.; Kumar, A.M.; Sudhakar, C.; Prasad, T.G.; Tuteja, N.; Udayakuuumar, M. Overexpression of a pea DNA helicase (PDH45) in peanut (Arachis hypogaea L.) confers improvement of cellular level tolerance and productivity under drought stress. Mol. Biotechnol. 2014, 56, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Shivakumara, T.N.; Sreevathsa, R.; Dash, P.K.; Sheshshayee, M.S.; Papolu, P.K.; Rao, U.; Tuteja, N.; UdayaKumar, M. Overexpression of Pea DNA Helicase 45 (PDH45) imparts tolerance to multiple abiotic stresses in chili (Capsicum annuum L.). Sci. Rep. 2017, 7, 2760. [Google Scholar] [CrossRef] [PubMed]
- Umate, P.; Tuteja, R.; Tuteja, N. Genome wide analysis of helicase gene family from rice and Arabidopsis: A comparison with yeast and human. Plant Mol. Biol. 2010, 73, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.W.; Yang, S.H.; Shin, K.H.; Lee, S.C.; Kim, S.H. The AtLRK10L1.2, Arabidopsis ortholog of wheat LRK10, is involved in ABA mediated signaling and drought resistance. Plant Cell Rep. 2015, 34, 447–455. [Google Scholar] [CrossRef]
- Lim, J.; Lim, C.W.; Lee, C.S. Pepper novel serine-threonine kinase CaDIK1 regulates drought tolerance via modulating ABA sensitivity. Front. Plant Sci. 2020, 11, 1133. [Google Scholar] [CrossRef]
- Devaiah, K.M.; Bali, G.; Athmaram, T.N.; Basha, M.S. Identification of two new genes from drought tolerant peanut up-regulated in response to drought. Plant Growth Regul. 2007, 52, 249–258. [Google Scholar] [CrossRef]
- Medini, M.; Baum, M.; Hamza, S. Transcript accumulation of putative drought responsive genes in drought-stressed chickpea seedlings. Afr. J. Biotechnol. 2009, 8, 4441–4449. [Google Scholar]
- Li, F.H.; Fu, F.L.; Sha, L.N.; He, L.; Li, W.C. Differential expression of serine/threonine protein phosphatase type-2C under drought stress in maize. Plant Mol. Biol. Rep. 2009, 27, 29–37. [Google Scholar] [CrossRef]
- Saakre, M.; Baburao, T.M.; Salim, A.P.; Ffancies, R.M.; Achuthan, V.A.; Thomas, G.; Sivarajar, S.R. Identification and characterization of genes responsible for drought tolerance in rice mediated by Pseudomonas fluorescens. Rice Sci. 2017, 24, 291–298. [Google Scholar] [CrossRef]
- Ahn, Y.K.; Tripathi, S.; Kim, J.H.; Cho, Y.I.H.E.; Kim, D.S.; Woo, J.G.; Yoon, M.K. Microsatellite marker information from high-through put next-generation sequence data of Capsicum annuum varieties Mandarin and Blackcluster. Sci. Hortic. 2014, 170, 123–130. [Google Scholar] [CrossRef]
- Novaes, E.; Drost, D.R.; Farmerie, W.G.; Pappas, G.J.; Grattapaglia, D.; Sederoff, R.R.; Kirst, M. High-throughput gene and SNP discovery in Eucalyptus grandis, an uncharacterized genome. BMC Genom. 2008, 9, 312. [Google Scholar] [CrossRef]
- Pujni, D.; Chaudhary, A.; Rajam, M.V. Increased tolerance to salinity and drought in transgenic indica rice by mannitol accumulation. J. Plant Biochem. Biotechnol. 2007, 16, 1–7. [Google Scholar] [CrossRef]
- Farooq, M.A.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Ferreira, D.O.; Fraga, O.T.; Pimenta, M.R.; Caetano, H.D.N.; Machado, J.P.B.; Carpinetti, P.A.; Brustolini, O.J.B.; Quadros, I.P.S.; Reis, P.A.B.; Fontes, E.P.B. GmNAC81 inversely modulates leaf senescence and drought tolerance. Front. Genet. 2020, 11, 601876. [Google Scholar] [CrossRef]
- Furlan, A.L.; Bianucci, E.; Giordano, W.; Castro, S.; Becker, D.F. Proline metabolic dynamics and implications in drought tolerance of peanut plants. Plant Physiol. Biochem. 2020, 151, 566–578. [Google Scholar] [CrossRef]
- Ni, Z.; Liu, N.; Yu, Y.; Bi, C.; Chen, Q.; Qu, Y. The cotton 70-kDa heat shock protein GhHSP70-26 plays a positive role in the drought stress response. Environ. Exp. Bot. 2021, 191, 104628. [Google Scholar] [CrossRef]
- Aslam, M.M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of abscisic acid-mediated drought stress responses in plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhou, L.; Lei, X.; Cao, H.; Wang, Y.; Dou, Y.; Tang, W.; Xia, W. Genome-wide identification of WRKY genes and their expression profiles under different abiotic stresses in Elaeis guineensis. PLoS ONE 2017, 12, e0189224. [Google Scholar]
- Pang, T.; Guo, L.; Shim, D.; Cannon, N.; Tang, S.; Chen, J.; Xia, X.; Yin, W.; Carlson, J.E. Characterization of the transcriptome of the xerophyte Ammopiptanthus mongolicus leaves under drought stress by 454 pyrosequencing. PLoS ONE 2015, 10, e0136495. [Google Scholar]
- Dong, S.; Liu, Y.; Niu, J.; Ning, Y.; Lin, S.; Zhang, Z. De novo transcriptome analysis of the Siberian apricot (Prunus sibirica L.) and search for potential SSR markers by 454 pyrosequencing. Gene 2014, 544, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Pandey, N.; Lakhwani, D.; Dubey, N.K.; Pathre, U.V.; Sawant, S.V. Comparative transcriptomic analysis of roots of contrasting Gossypium herbaceum genotypes revealing adaptation to drought. BMC Genom. 2012, 13, 680. [Google Scholar]
- Wang, X.; Li, Y.; Wang, X.; Li, X.; Dong, S. Physiology and metabonomics reveal differences in drought resistance among soybean varieties. Bot. Stud. 2022, 63, 8. [Google Scholar] [PubMed]
- Nuruzzaman, M.; Sharoni, A.M.; Satoh, K.; Al-Shammari, T.; Shimizu, T.; Sasaya, T.; Omura, T.; Kikuchi, S. The thioredoxin gene family in rice: Genome-wide identification and expression profiling under different biotic and abiotic treatments. Biochem. Biophys. Res. Commun. 2013, 423, 417–423. [Google Scholar] [CrossRef]
- Elasad, M.; Ahmad, A.; Wang, H.; Ma, L.; Yu, S.; Wei, H. Overexpression of CDSP32 (GhTRX134) cotton gene enhances drought, salt, and oxidative stress tolerance in Arabidopsis. Plants 2020, 9, 1388. [Google Scholar] [CrossRef]
- Lv, Y.; Xu, L.; Dossa, K.; Zhou, K.; Zhu, M.; Xie, H.; Tang, S.; Yu, Y.; Guo, X.; Zhou, B. Identification of putative drought-responsive genes in rice using gene co-expression analysis. Bioinformation 2019, 15, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, L.; Li, J.; Sun, P.; Lu, M.; Hu, J. Comparative metabolomics analysis reveals different metabolic responses to drought in tolerant and susceptible poplar species. Physiol. Plant 2020, 168, 531–546. [Google Scholar]
- La Rota, M.; Kantety, R.; Yu, J.K.; Sorrells, M. Nonrandom distribution and frequencies of genomic and EST-derived microsatellite markers in rice, wheat, and barley. BMC Genom. 2005, 6, 23. [Google Scholar]
- Liang, L.; Pardee, A.B. Differential display of eukaryotic massager RNA by means of the polymerase chain reaction. Science 1992, 258, 967–997. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar]
- Hulsen, T.; Jacob de Vlieg, T.D.; Alkema, W. BioVenn—A web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genom. 2008, 9, 488. [Google Scholar]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Leaf Sample | Root Sample | ||||
|---|---|---|---|---|---|---|
| Total Bands | Polymorphism (%) | Monophism (%) | Total Bands | Polymorphism (%) | Monophism (%) | |
| O1A2 | 45 | 86.67 | 13.33 | 51 | 90.20 | 9.80 |
| O1A3 | 47 | 72.34 | 27.66 | 67 | 95.52 | 4.48 |
| O1A4 | 74 | 81.08 | 18.92 | 107 | 97.21 | 2.89 |
| O1A5 | 82 | 87.80 | 12.20 | 90 | 96.67 | 3.33 |
| O2A2 | 90 | 80.00 | 20.00 | 95 | 93.68 | 6.32 |
| O2A3 | 82 | 75.61 | 24.39 | 106 | 97.20 | 1.80 |
| O2A5 | 135 | 96.30 | 3.70 | 82 | 98.78 | 1.22 |
| O3A3 | 109 | 83.47 | 16.53 | 111 | 99.10 | 0.90 |
| O3A4 | 116 | 79.31 | 20.69 | 105 | 98.10 | 1.90 |
| O3A5 | 111 | 79.28 | 20.72 | 118 | 97.46 | 2.54 |
| O4A2 | 117 | 86.32 | 13.68 | 111 | 94.59 | 5.41 |
| O4A3 | 137 | 88.32 | 11.68 | 129 | 98.45 | 1.55 |
| O4A5 | 75 | 94.67 | 5.33 | 76 | 97.37 | 2.63 |
| O4A6 | 106 | 66.04 | 33.96 | 111 | 90.99 | 9.01 |
| Total | 1326 | - | - | 1359 | - | - |
| Average | 94.71 | 77.66 | 17.34 | 97.07 | 96.09 | 3.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuanlaong, S.; Wuthisuthimethavee, S.; Azzeme, A.M.; Suraninpong, P. Optimized Method for the Identification of Candidate Genes and Molecular Maker Development Related to Drought Tolerance in Oil Palm (Elaeis guineensis Jacq.). Plants 2022, 11, 2317. https://doi.org/10.3390/plants11172317
Nuanlaong S, Wuthisuthimethavee S, Azzeme AM, Suraninpong P. Optimized Method for the Identification of Candidate Genes and Molecular Maker Development Related to Drought Tolerance in Oil Palm (Elaeis guineensis Jacq.). Plants. 2022; 11(17):2317. https://doi.org/10.3390/plants11172317
Chicago/Turabian StyleNuanlaong, Sunya, Suwit Wuthisuthimethavee, Azzreena Mohamad Azzeme, and Potjamarn Suraninpong. 2022. "Optimized Method for the Identification of Candidate Genes and Molecular Maker Development Related to Drought Tolerance in Oil Palm (Elaeis guineensis Jacq.)" Plants 11, no. 17: 2317. https://doi.org/10.3390/plants11172317