
Classification and Identification of S Haplotypes in Radish Based on SRK Kinase Domain Sequence Analysis

Abstract
:1. Introduction
2. Results
2.1. Comparison and Unified Nomenclature of Published S haplotype in Radish (Raphanus sativus L.)
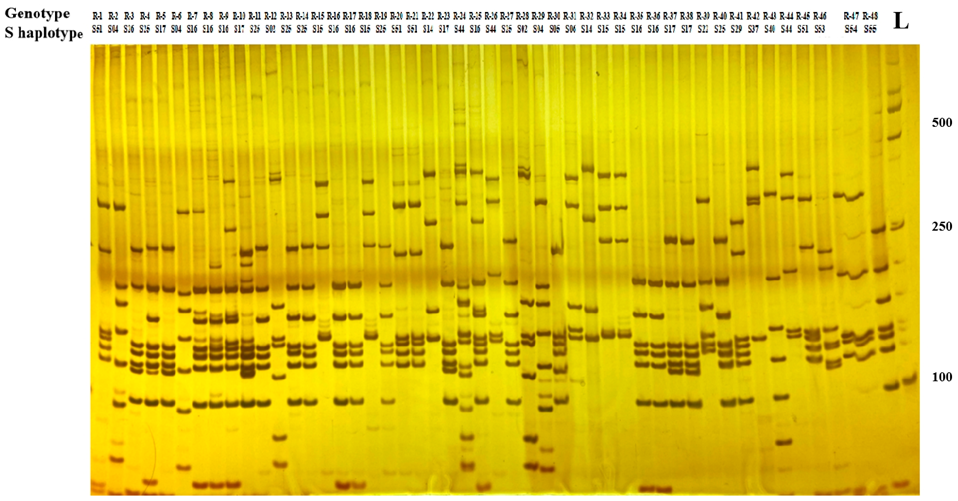
2.2. SRK Kinase Domain-Based Classification and Identification of S haplotypes
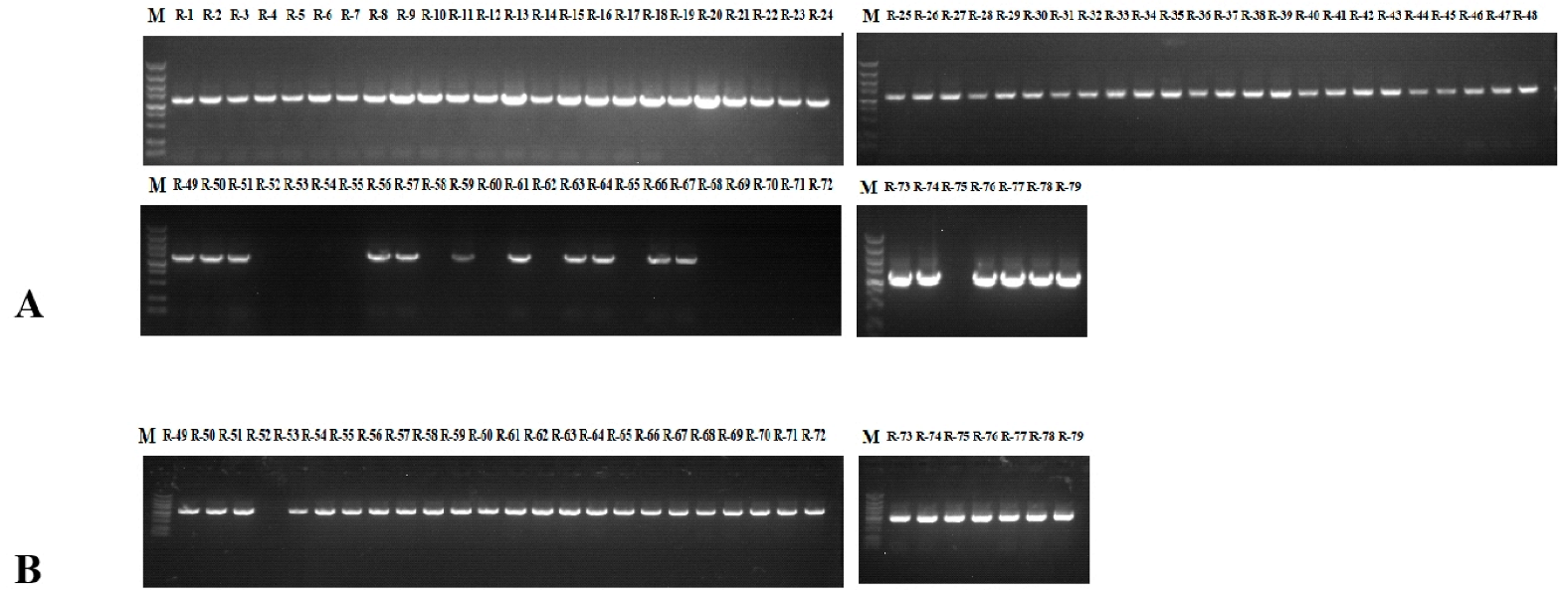
2.3. PCR-RFLP Analysis of SRK Alleles
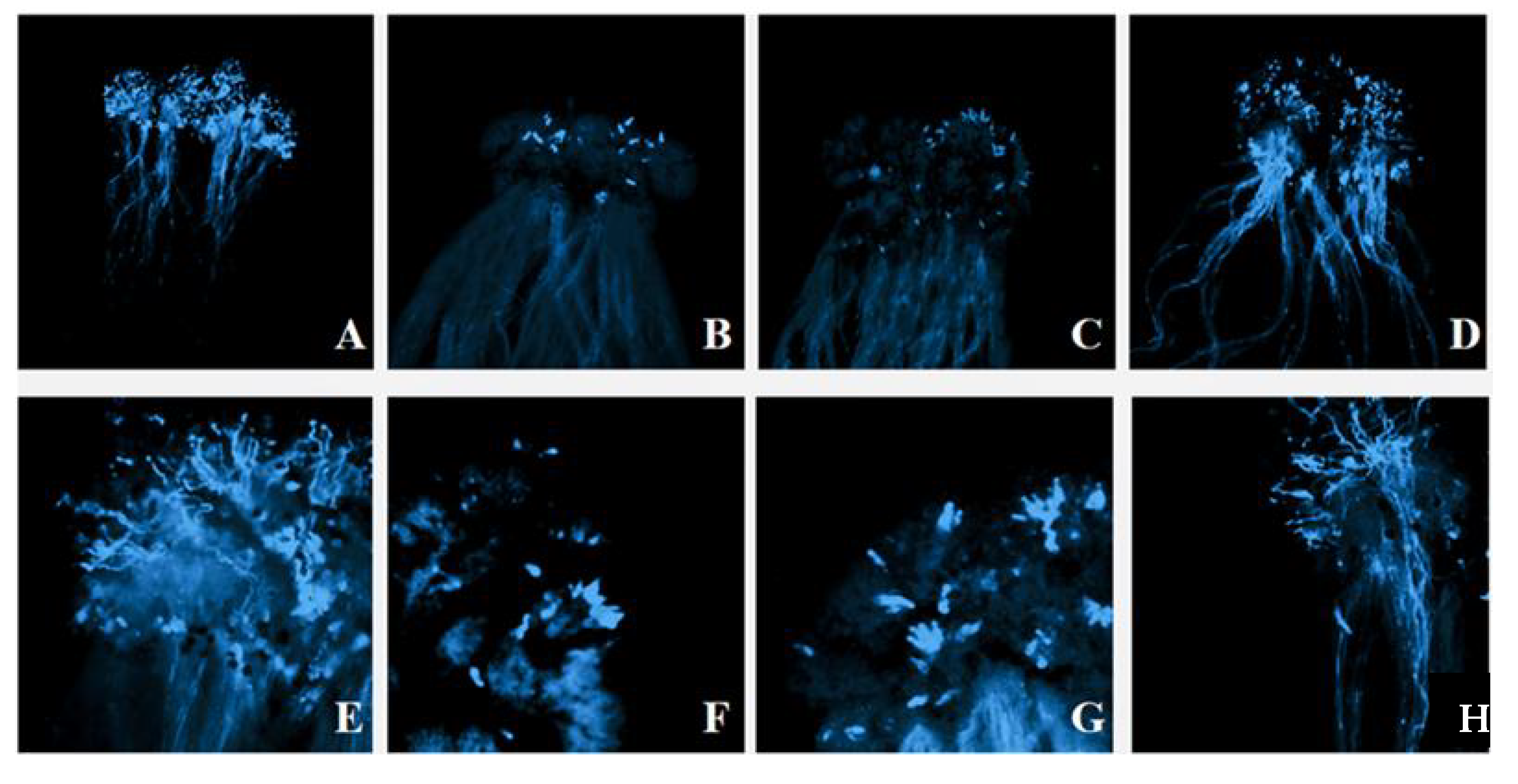
2.4. Pollen Germination and Tube Growth
2.5. Compatibility Index Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Amplification and Sequencing of the PCR Products
4.2.1. Extraction of Genomic DNA and Amplification of the SRK Gene
4.2.2. Determination of Nucleotide Sequences
4.3. PCR-RFLP Analysis
4.4. Pollination Tests
4.5. Aniline Blue Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stone, S.L.; Goring, D.R. The molecular biology of self-incompatibility systems in flowering plants. Plant Cell. Tissue Organ Culture. 2001, 67, 93–114. [Google Scholar] [CrossRef]
- Watanabe, M.; Suwabe, K.; Suzuki, G. Molecular genetics, physiology and biology of self-incompatibility in Brassicaceae. Proc. Jpn. Acad. 2012, 88, 519–535. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, K.; Takasaki, T.; Watanabe, M.; Hinata, K. High sequence similarity between SLG and the receptor domain of SRK is not necessarily involved in higher dominance relationships in stigma in self-incompatible Brassica rapa L. Sex. Plant Reprod. 1998, 11, 292–294. [Google Scholar] [CrossRef]
- Zeng, J. Dissecting pistil responses to incompatible and compatible pollen in self-incompatibility Brassica oleracea using comparative proteomics. Protein J. 2017, 36, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Ockendon, D.J. The S-allele collection of Brassica oleracea. Acta Hortic. Sin. 2000, 539, 25–30. [Google Scholar] [CrossRef]
- Wang, Q.; Zheng, P.; Zhang, L. Identification and classification of S haplotypes in radish (Raphanus sativus L). Plant Breed. 2019, 138, 121–130. [Google Scholar]
- Zhao, T.R. Detection of self-Incompatibility of Radish and Identification of Related Genes. Master’s Dissertation, Nanjing Agricultural University, Nanjing, China, 2007. [Google Scholar]
- He, S.M.; Li, C.Y.; Lan, C.Y.; Zou, M.; Ren, X.S.; Si, J.; Li, C.Q.; Song, H.Y. Effects of transgenic MLPK antisense gene on self-incompatibility of cabbage. Chin. J. Hortic. 2015, 2, 11. [Google Scholar]
- Zhang, E.H. Determination of cabbage self-incompatibility by fluorescence microscopy. Shanxi Agric. Sci. 1989, 1, 6–7. Available online: https://CNKI:SUN:SNKX.0.1989-01-003 (accessed on 10 June 2022).
- Fang, Z.Y.; Sun, P.T.; Liu, Y.M. Several issues on utilization of heterosis and selection of self-incompatible lines in cabbage. Chin. Agric. Sci. 1983, 3, 51–62. Available online: https://www.chinaagrisci.com/CN/Y1983/V16/I03/51 (accessed on 10 June 2022).
- Pastuglia, M.; Ruffio, C.V.; Delorme, V.; Gaude, T.; Cock, J.M. A functional S locus anther gene is not required for the self-incompatibility response in Brassica oleracea. Plant Cell. 1997, 9, 2065. [Google Scholar]
- Kitashiba, H.; Nasrallah, J.B. Self-incompatibility in Brassicaceae crops: Lessons for interspecific incompatibility. Breed Sci. 2014, 64, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Kusaba, M.; Watanabe, M.; Hinata, K. Registration of S alleles in Brassica campestris L. by the restriction fragment sizes of SLGs. Theor. Appl. Genet. 1996, 92, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Brace, J.; Ockendon, D.J.; King, G.J. Development of a method for the identification of S alleles in Brassica oleracea based on digestion of PCR-amplified DNA with restriction endonucleases. Sex. Plant Reprod. 1993, 6, 133–138. [Google Scholar] [CrossRef]
- Sakamoto, K.; Kusaba, M.; Nishio, T. Polymorphism of the S-locus glycoprotein gene (SLG) and the S-locus related gene (SLR1) in Raphanus sativus L. and self-incompatible ornamental plants in the Brassicaceae. Mol. Gen. Genet. 1998, 258, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Cho, H.; Lee, S.; Cho, Y.H.; Kim, B.D. Identification and classification of S haplotypes in Raphanus sativus by PCR-RFLP of the S locus glycoprotein (SLG) gene and the S locus receptor kinase (SRK) gene. Theor. Appl. Genet. 2002, 104, 1253–1262. [Google Scholar] [CrossRef]
- Kawamura, K.; Kawanabe, T.; Shimizu, M. Genetic characterization of inbred lines of Chinese cabbage by DNA markers; towards the application of DNA markers to breeding of F1 hybrid cultivars. Data Brief. 2015, 6, 229–237. [Google Scholar] [CrossRef]
- Yu, H.F.; Wang, J.S.; Zhao, Z.Q.; Sheng, X.G.; Gu, H.H. Identification of the S Haplotype of the Self-Incompatible DH Line in Broccoli; Chinese Horticultural Society: LiJiang, China, 2013. [Google Scholar]
- Zhang, A.F.; Li, Y.; Gao, S.Y.; Shi, G.J.; Hou, X.L. Identification of self-incompatible S haplotypes in non-heading Chinese cabbage. Northwestern J. Bot. 2008, 28, 1720–1727. [Google Scholar]
- Cui, Y.; Brugiere, N.; Jackman, L.; Bi, Y.M.; Rothstein, S.J. Structural and transcriptional comparative analysis of the S locus regions in two self-incompatible Brassica napus lines. Plant Cell. 1999, 11, 2217–2231. [Google Scholar] [CrossRef]
- Fukai, E.; Fujimoto, R.; Nishio, T. Genomic organization of the S core region and the S flanking regions of a Class-II S haplotype in Brassica rapa. Mol. Genet. Genom. 2003, 269, 361–369. [Google Scholar] [CrossRef]
- Zhou, Y.K.; Gong, Y.Q.; Liu, L.W.; Ma, E.L.; Song, L.J.; Wang, L.Z. Rapid Identification of Self-Incompatibility between Radish and Broccoli; The Second Council and Academic Symposium of the Tenth Chinese Horticultural Society: Nanjing, China, 2007. [Google Scholar]
- Okamoto, S.; Sato, Y.; Sakamoto, K.; Nishio, T. Distribution of similar self-incompatibility (S) haplotypes in different genera, Raphanus and Brassica. Sex. Plant Reprod. 2004, 17, 33–39. [Google Scholar] [CrossRef]
- Zhou, Y.K.; Shao, W.T.; Gong, Y.Q.; Zhu, X.W.; Ma, E.L.; Zhao, T.R. Radish RsSRK-a gene cDNA cloning and expression characteristics analysis. J. Nanjing Agric. Univ. 2010, 33, 13–18. [Google Scholar]
- Koh, J.C.O.; Hoebee, S.E.; Newbigin, E.J. Sporophytic Self-Incompatability; Academic Press: Oxford, UK, 2017; Volume 2, pp. 334–340. [Google Scholar]
- Kim, D.; Jung, J.; Choi, Y.; Kim, S. Development of a system for S locus haplotyping based on the polymorphic SLL2 gene tightly linked to the locus determining self-incompatibility in radish (Raphanus sativus L.). Euphytica 2016, 209, 525–535. [Google Scholar] [CrossRef]
- Kim, D.; Kim, S. Identification of the S locus core sequences determining self-incompatibility and S multigene family from draft genome sequences of radish (Raphanus sativus L.). Euphytica 2018, 214, 16–28. [Google Scholar] [CrossRef]
- Kim, D.; Kim, S. Development of a new S locus haplotyping system based on three tightly linked genes in the S locus controlling self-incompatibility in radish (Raphanus sativus L.). Sci. Hortic. 2019, 243, 70–77. [Google Scholar] [CrossRef]
- Nishio, T.; Sakamoto, K. Polymorphism of self-incompatibility genes. In The Radish Genome; Springer International Publishing: Cham, Switzerland, 2017; pp. 177–188. [Google Scholar]
- Haseyama, Y.; Kitashiba, H.; Okamoto, S.; Tonouchi, E.; Sakamoto, K.; Nishio, A. Nucleotide sequence analysis of S-locus genes to unify S haplotype nomenclature in radish. Mol. Breed. 2018, 38, 116–127. [Google Scholar] [CrossRef]
- Nasrallah, J.B.; Nasrallah, M.E. Pollen-stigma signaling in the sporophytic self-incompatibility response. Plant Cell. 1993, 5, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.C.; Yao, Q.J.; Yuan, Y.X.; Wei, F.; Xu, Q.; Zhao, Y.Y.; Zhang, Q.; Wang, Z.Y.; Jiang, W.S.; Zhang, X.W. Distribution of S haplotypes in Chinese cabbage high-generation inbred lines. Henan Agric. Sci. 2015, 44, 109. [Google Scholar]
- Nasrallah, J.B. Evolution of the Brassica self-incompatibility locus: A look into S-locus gene polymorphisms. Proc. Natl. Acad. Sci. USA 1997, 94, 9516–9519. Available online: https://www.pnas.org/doi/abs/10.1073/pnas.94.18.9516 (accessed on 10 June 2022). [CrossRef] [Green Version]
- Li, X.; Hu, Z.H.; Zhou, G.L.; Wang, A.H. Molecular identification of self-incompatible S haplotypes in four non-heading Chinese cabbage. Hubei Agric. Sci. 2015, 54, 948–3951. [Google Scholar]
- Wei, W.H. Inheritance and SSR Markers of Self-Compatibility in Brassica campestris L.; Gansu Agricultural University: Lanzhou, China, 2009; Available online: https://CNKI:CDMD:2.2009.253310 (accessed on 10 June 2022).
- Tian, L.; Miao, W.W.; Liu, J.S.; Fang, Z.Y.; Liu, Y.M.; Yang, L.M.; Zhuang, M. Identification of S haplotypes in cabbage inbred lines (Brassica oleracea var. capitata L.). Sci. Hortic. 2013, 164, 400–408. [Google Scholar] [CrossRef]
- Tian, L.; Zhuang, M.; Miao, W.W.; Fang, Z.Y.; Liu, Y.M.; Yang, L.M.; Zhang, Y.Y. The rapid classification and identification of S haplotypes in cabbage inbred lines. Acta Hortic. Sin. 2011, 38, 2545. Available online: http://www.ahs.ac.cn/CN/article/downloadArticleFile.do?attachType=PDF&id=3405 (accessed on 1 August 2011).
- Zheng, M.; Zhu, C.Z.; Liu, M.C.; Xu, B.B.; Ma, C.F.; Li, Q.F.; Ren, X.S.; Si, J.; Song, H.Y. Haplotype identification and verification of self-incompatible cabbage lines based on SRK gene sequence analysis. Chin. Veg. 2018, 3, 32–39. Available online: http://www.cqvip.com/QK/94878X/201803/674591563.html (accessed on 24 December 2017).
- Park, S.H.; Kim, K.T.; Lim, S.H.; Yoon, M.K.; Park, H.G. Classification and identification of S-haplotypes using PCR-RFLP and measuring the self-incompatibility activity in radish (Raphanus sativus L.). HortScience 2005, 40, 1113. [Google Scholar] [CrossRef]
- Kitahara, K.; Matsumoto, S. Cloning of the S25 cDNA from ′McIntosh′ apple and an S25-allele identification method. J. Hortic. Sci. 2002, 77, 724–728. [Google Scholar] [CrossRef]
- Zisovich, A.H.; Stern, R.A.; Shafir, S.; Goldway, M. Identification of seven S-alleles from the European pear (Pyrus communis) and the determination of compatibility among cultivars. J. Pomol. Hortic. Sci. 2004, 79, 101–106. [Google Scholar]
- Park, J.I.; Lee, S.S.; Watanabe, M.; Takahata, Y.; Nou, S.I. Identification of S-alleles using polymerase chain reaction-cleaved amplified polymorphic sequence of the S-locus receptor kinase in inbreeding lines of Brassica oleracea. Plant Breed. 2002, 121, 192–197. [Google Scholar] [CrossRef]
- Duan, Q.H.; Kita, D.; Johnson, E.A.; Aggarwal, M.; Gates, L.; Wu, H.M.; Cheung, A.Y. Reactive oxygen species mediate pollen tube rupture to release sperm for fertilization in Arabidopsis. Nat. Commun. 2014, 5, 547–572. [Google Scholar] [CrossRef] [PubMed]
- Su, S.Q.; Dai, H.M.; Wang, X.Y.; Wang, C.; Zeng, W.Q.; Huang, J.B.; Duan, Q.H. Ethylene negatively mediates self-incompatibility response in Brassica rapa. Biochem. Biophys. Res. Commun. 2020, 525, 600–606. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S Haplotyes (Renumbered in This Study) | S Haplotypes (Okamoto) [23] | S Haplotypes (Lim) [16] | S Haplotypes (Kim D) [26,27,28] | S Haplotypes (Other Group) | Similarity Okamoto vs. Lim | Similarity Lim vs. Kim D (or Other Group) |
|---|---|---|---|---|---|---|
| NAU-S1 | S1 | S6 | S31 | 99.8% | 100% | |
| NAU-S2 | S2 | S2 | S2 | 100% | 100% | |
| NAU-S3 | S3 | S12 | 99.9% | |||
| NAU-S4 | S4 | S7 | S6 | 100% | 100% | |
| NAU-S5 | S5 | |||||
| NAU-S6 | S6 | S18 | S15 | 100% | 100% | |
| NAU-S7 | S8 | |||||
| NAU-S8 | S9 | S21 | S4/S5 (SP11-4/5) (Kim H, 2003) | 99.3% | 98.7% | |
| NAU-S9 | S11 | |||||
| NAU-S10 | S14 | |||||
| NAU-S11 | S15 | S2 (SP11-2) (Kim H, 2003) | 100% | |||
| NAU-S12 | S17 | S9 | S5 | 99.7% | 100% | |
| NAU-S13 | S18 | |||||
| NAU-S14 | S19 | S8 | S7 (S30) | 100% | 100% | |
| NAU-S15 | S21 | |||||
| NAU-S16 | S22 (S7) | S16 | S13 | 100% | 99.1% | |
| NAU-S17 | S23 | S20 | S16 | 99.9% | 100% | |
| NAU-S18 | S25 | S17 | 99.9% | |||
| NAU-S19 | S26 | S4 | S1 (SP11-1) (Kim H, 2003) | 100% | 100% | |
| NAU-S20 | S28 | |||||
| NAU-S21 | S29 | S26 | S3 (SP11-3) (Kim H, 2003) | 99.4% | 100% | |
| NAU-S22 | S30 | S1 | S1 | 100% | 100% | |
| NAU-S23 | S31 | |||||
| NAU-S24 | S5 | |||||
| NAU-S25 | S10 | S14 | 99.73% | |||
| NAU-S26 | S11 | S9 (S27) | S38 [6] DQ984139 [7] | 100% | ||
| NAU-S27 | S15 | S12 | 100% | |||
| NAU-S28 | S22 | |||||
| NAU-S29 | S23 | S19 | 100% | |||
| NAU-S30 | S24 | |||||
| NAU-S31 | S27 | S24 | 99.8% | |||
| NAU-S32 | S29 | |||||
| NAU-S33 | S30 | |||||
| NAU-S34 | S31 | S29 | 100% | |||
| NAU-S35 | S4 | |||||
| NAU-S36 | S8 | |||||
| NAU-S37 | S10 | |||||
| NAU-S38 | S11 | |||||
| NAU-S39 | S17 | |||||
| NAU-S40 | S18 | |||||
| NAU-S41 | S20 | |||||
| NAU-S42 | S21 | |||||
| NAU-S43 | S22 | |||||
| NAU-S44 | S23 | |||||
| NAU-S45 | S25 | S39 [6] | 100% | |||
| NAU-S46 | S28 | |||||
| NAU-S47 | S201 (Niikura, 1997) | |||||
| NAU-S48 | EF056499 [7] | |||||
| NAU-S49 | GQ121139 [22] | |||||
| NAU-S50 | S40 [6] | |||||
| NAU-S51 | S48 (Kim S, 2021) | |||||
| NAU-S52 | S6 [25] |
| Class of S Haplotypes | Genotype | Frequency (%) |
|---|---|---|
| Class I | NAU-Rs1, NAU-Rs2, NAU-Rs3, NAU-Rs4, NAU-Rs5, NAU-Rs6, NAU-Rs7, NAU-Rs8, NAU-Rs9, NAU-Rs10, NAU-Rs11, NAU-Rs12, NAU-Rs13, NAU-Rs14, NAU-Rs15, NAU-Rs16, NAU-Rs17, NAU-Rs18, NAU-Rs19, NAU-Rs20, NAU-Rs21, NAU-Rs22, NAU-Rs23, NAU-Rs24, NAU-Rs25, NAU-Rs26, NAU-Rs27, NAU-Rs28, NAU-Rs29, NAU-Rs30, NAU-Rs31, NAU-Rs32, NAU-Rs33, NAU-Rs34, NAU-Rs35, NAU-Rs36, NAU-Rs37, NAU-Rs38, NAU-Rs39, NAU-Rs40, NAU-Rs41, NAU-Rs42, NAU-Rs43, NAU-Rs44, NAU-Rs45, NAU-Rs46, NAU-Rs47, NAU-Rs48 | 60. 76 |
| Class II | NAU-Rs53, NAU-Rs54, NAU-Rs55, NAU-Rs58, NAU-Rs60, NAU-Rs62, NAU-Rs65, NAU-Rs68, NAU-Rs69, NAU-Rs70, NAU-Rs71, NAU-Rs72, NAU-Rs75 | 16. 46 |
| Class I/Class II | NAU-Rs49, NAU-Rs50, NAU-Rs51, NAU-Rs56, NAU-Rs57, NAU-Rs59, NAU-Rs61, NAU-Rs63, NAU-Rs64, NAU-Rs66, NAU-Rs67, NAU-Rs73, NAU-Rs74, NAU-Rs76, NAU-Rs77, NAU-Rs78, NAU-Rs79 | 21. 52 |
| S Haplotype | Number of Occurrences | Frequency (%) |
|---|---|---|
| NAU-S16 | 8 | 16. 67 |
| NAU-S25 | 7 | 14. 58 |
| NAU-S17 | 5 | 10. 41 |
| NAU-S15/NAU-S51 | 4 | 8. 33 |
| NAU-S04/NAU-S44 | 3 | 6. 25 |
| NAU-S02/NAU-S14 | 2 | 4. 17 |
| NAU-S05/NAU-S06/NAU-S22/NAU-S26/NAU-S29/NAU-S37/NAU-S40NAU-S53/NAU-S54/NAU-S55 | 1 | 2. 08 |
| S Haplotype | Genotype | Number of Occurrences | Frequency (%) |
|---|---|---|---|
| NAU-S39 | NAU-Rs53, NAU-Rs55, NAU-Rs58, NAU-Rs70, NAU-Rs72 | 5 | 38.46 |
| NAU-S52 | NAU-Rs60, NAU-Rs65, NAU-Rs69, NAU-Rs75 | 4 | 30.77 |
| NAU-S43 | NAU-Rs54, NAU-Rs68, NAU-Rs71 | 3 | 23.08 |
| NAU-S38 | NAU-Rs62 | 1 | 7.69 |
| Genotype | S Haplotype (Class I) | S Haplotype (Class II) |
|---|---|---|
| NAU-Rs49 | NAU-S17 | NAU-S39 |
| NAU-Rs50 | NAU-S17 | NAU-S43 |
| NAU-Rs51 | NAU-S17 | NAU-S43 |
| NAU-Rs56 | NAU-S26 | NAU-S43 |
| NAU-Rs57 | NAU-S14 | NAU-S43 |
| NAU-Rs59 | NAU-S17 | NAU-S52 |
| NAU-Rs61 | NAU-S17 | NAU-S52 |
| NAU-Rs63 | NAU-S26 | NAU-S39 |
| NAU-Rs64 | NAU-S17 | NAU-S39 |
| NAU-Rs66 | NAU-S26 | NAU-S38 |
| NAU-Rs67 | NAU-S25 | NAU-S52 |
| NAU-Rs73 | NAU-S26 | NAU-S38 |
| NAU-Rs74 | NAU-S26 | NAU-S52 |
| NAU-Rs76 | NAU-S17 | NAU-S52 |
| NAU-Rs77 | NAU-S55 | NAU-S39 |
| NAU-Rs78 | NAU-S26 | NAU-S39 |
| NAU-Rs79 | NAU-S17 | NAU-S52 |
| Pollination Combinations | Compatibility Index | Podding Rate (%) | Compatibility | S Haplotype |
|---|---|---|---|---|
| NAU-Rs5 × NAU-Rs38 | 0.88 | 27.57 | Weak Incompatibility | NAU-S17 |
| NAU-Rs44 × NAU-Rs24 | 0.52 | 24.09 | Weak Incompatibility | NAU-S44 |
| NAU-Rs7 × NAU-Rs9 | 0.20 | 12.07 | Incompatibility | NAU-S16 |
| NAU-Rs7 × NAU-Rs16 | 0.17 | 12.09 | Incompatibility | NAU-S16 |
| NAU-Rs4 × NAU-Rs40 | 0.47 | 19.75 | Incompatibility | NAU-S25 |
| NAU-Rs40 × NAU-Rs4 | 0.37 | 21.79 | Incompatibility | NAU-S25 |
| NAU-Rs1 × NAU-Rs40 | 3.76 | 78.93 | Compatibility | NAU-S51, NAU-S25 |
| NAU-Rs44 × NAU-Rs30 | 3.08 | 74.64 | Compatibility | NAU-S44, NAU-S05 |
| NAU-Rs34 × NAU-Rs40 | 2.83 | 67.90 | Compatibility | NAU-S15, NAU-S25 |
| NAU-Rs7 × NAU-Rs18 | 1.85 | 72.98 | Weak Incompatibility | NAU-S16, NAU-S15 |
| NAU-Rs7 × NAU-Rs1 | 2.05 | 71.75 | Compatibility | NAU-S16, NAU-S51 |
| NAU-Rs1⊗ | 0.09 | 12.40 | Self-incompatibility | NAU-S51 |
| NAU-Rs30⊗ | 0.46 | 20.69 | Self-incompatibility | NAU-S05 |
| NAU-Rs33⊗ | 0.17 | 13.22 | Self-incompatibility | NAU-S15 |
| NAU-Rs45⊗ | 0.11 | 14.59 | Self-incompatibility | NAU-S51 |
| Material Code | Color of Root Skin | Color of Fleshy Root | Fleshy Root Shape | Leaf Morphology | Source |
|---|---|---|---|---|---|
| NAU-Rs1 | Green | Green | Cylindrical | Entire | Henan China |
| NAU-Rs2 | Red | White | Spherical | Entire | Jiangsu China |
| NAU-Rs3 | Green | Green | Cylindrical | Entire | Henan China |
| NAU-Rs4 | Green | Green | Cylindrical | Lyrate | Jiangsu China |
| NAU-Rs5 | Red | White | Spherical | Entire | Sichuan China |
| NAU-Rs6 | White | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs7 | White | White | Long and Tapered | Entire | Beijing China |
| NAU-Rs8 | White | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs9 | White | White | Long and Tapered | Lyrate | Jiangsu China |
| NAU-Rs10 | Red | White | Cylindrical | Entire | Sichuan China |
| NAU-Rs11 | White | White | Cylindrical | Lyrate | South Korea |
| NAU-Rs12 | Red | White | Apically Bulbous | Lyrate | America |
| NAU-Rs13 | White | White | Cylindrical | Lyrate | Jiangsu China |
| NAU-Rs14 | Red | White | Elliptic | Lyrate | Jiangsu China |
| NAU-Rs15 | Red | White | Elliptic | Lyrate | Jiangsu China |
| NAU-Rs16 | White | White | Long and Tapered | Lyrate | South Korea |
| NAU-Rs17 | White | White | Long and Tapered | Lyrate | South Korea |
| NAU-Rs18 | White | White | Long and Tapered | Lyrate | Guangdong China |
| NAU-Rs19 | Purple | White | Apically Bulbous | Lyrate | Wuhan China |
| NAU-Rs20 | White | White | Long and Tapered | Lyrate | Jiangsu China |
| NAU-Rs21 | Red | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs22 | White | White | Long and Tapered | Lyrate | South Korea |
| NAU-Rs23 | Green | White | Apically Bulbous | Lyrate | Henan China |
| NAU-Rs24 | Red | White | Elliptic | Entire | Jiangsu China |
| NAU-Rs25 | White | White | Long and Tapered | Entire | South Korea |
| NAU-Rs26 | White | White | Long and Tapered | Sinuate | Wuhan China |
| NAU-Rs27 | Purple | White | Apically Bulbous | Lyrate | Xizang China |
| NAU-Rs28 | White | White | Spherical | Lyrate | Jiangsu China |
| NAU-Rs29 | Red | White | Elliptic | Entire | Jiangsu China |
| NAU-Rs30 | White | White | Long and Tapered | Lyrate | Anhui China |
| NAU-Rs31 | White | White | Apically Bulbous | Lyrate | Jiangsu China |
| NAU-Rs32 | Red | White | Elliptic | Entire | Jiangsu China |
| NAU-Rs33 | Green | Red | Spherical | Entire | Jiangsu China |
| NAU-Rs34 | White | White | Cylindrical | Lyrate | South Korea |
| NAU-Rs35 | White | White | Long and Tapered | Lyrate | Jiangsu China |
| NAU-Rs36 | Yellow | White | Spherical | Lyrate | Jiangsu China |
| NAU-Rs37 | Red | White | Cylindrical | Entire | Sichuan China |
| NAU-Rs38 | White | White | Apically Bulbous | Entire | Jiangsu China |
| NAU-Rs39 | Red | White | Cylindrical | Entire | Shandong China |
| NAU-Rs40 | Green | Green | Long and Tapered | Lyrate | Shandong China |
| NAU-Rs41 | Green | Green | Cylindrical | Lyrate | Jiangsu China |
| NAU-Rs42 | White | White | Cylindrical | Lyrate | Jiangsu China |
| NAU-Rs43 | Green | Green | Apically Bulbous | Lyrate | Henan China |
| NAU-Rs44 | Red | White | Apically Bulbous | Entire | Jiangsu China |
| NAU-Rs45 | White | White | Elliptic | Lyrate | Jiangsu China |
| NAU-Rs46 | White | White | Spherical | Lyrate | Jiangsu China |
| NAU-Rs47 | Red | White | Long and Tapered | Entire | Sichuan China |
| NAU-Rs48 | Red | White | Cylindrical | Sinuate | Jiangsu China |
| NAU-Rs49 | White | White | Long and Tapered | Entire | Yunnan China |
| NAU-Rs50 | Red | White | Cylindrical | Sinuate | Jiangsu China |
| NAU-Rs51 | Green | Green | Cylindrical | Entire | Shandong China |
| NAU-Rs52 | Green | Red | Cylindrical | Entire | Beijing China |
| NAU-Rs53 | White | White | Long and Tapered | Lyrate | Japan |
| NAU-Rs54 | Red | White | Cylindrical | Sinuate | Sichuan China |
| NAU-Rs55 | White | White | Cylindrical | Sinuate | Jiangsu China |
| NAU-Rs56 | Red | White | Cylindrical | Sinuate | Jiangsu China |
| NAU-Rs57 | White | White | Cylindrical | Sinuate | South Korea |
| NAU-Rs58 | White | White | Long and Tapered | Sinuate | Jiangsu China |
| NAU-Rs59 | White | White | Cylindrical | Entire | Japan |
| NAU-Rs60 | White | White | Cylindrical | Entire | South Korea |
| NAU-Rs61 | White | White | Cylindrical | Entire | Jiangsu China |
| NAU-Rs62 | White | White | Long and Tapered | Entire | South Korea |
| NAU-Rs63 | White | White | Cylindrical | Entire | Jiangsu China |
| NAU-Rs64 | White | White | Cylindrical | Entire | Jiangsu China |
| NAU-Rs65 | White | White | Long and Tapered | Entire | South Korea |
| NAU-Rs66 | Red | White | Cylindrical | Entire | Jiangsu China |
| NAU-Rs67 | White | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs68 | White | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs69 | Red | White | Cylindrical | Entire | Jiangsu China |
| NAU-Rs70 | Green | Green | Cylindrical | Entire | Tianjin China |
| NAU-Rs71 | Green | White | Cylindrical | Entire | Henan China |
| NAU-Rs72 | Green | White | Cylindrical | Entire | Shandong China |
| NAU-Rs73 | White | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs74 | Red | White | Cylindrical | Entire | Jiangsu China |
| NAU-Rs75 | White | White | Long and Tapered | Entire | Shandong China |
| NAU-Rs76 | Red | White | Long and Tapered | Entire | Jiangsu China |
| NAU-Rs77 | White | White | Long and Tapered | Lyrate | Jiangsu China |
| NAU-Rs78 | White | White | Long and Tapered | Entire | Beijing China |
| NAU-Rs79 | White | White | Long and Tapered | Entire | Jiangsu China |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, M.; Yi, X.; Wang, Q.; Wang, J.; Wang, S.; Liu, L.; Xu, L.; Wang, Y. Classification and Identification of S Haplotypes in Radish Based on SRK Kinase Domain Sequence Analysis. Plants 2022, 11, 2304. https://doi.org/10.3390/plants11172304
Ni M, Yi X, Wang Q, Wang J, Wang S, Liu L, Xu L, Wang Y. Classification and Identification of S Haplotypes in Radish Based on SRK Kinase Domain Sequence Analysis. Plants. 2022; 11(17):2304. https://doi.org/10.3390/plants11172304
Chicago/Turabian StyleNi, Meng, Xiaofang Yi, Qin Wang, Juan Wang, Shuang Wang, Liwang Liu, Liang Xu, and Yan Wang. 2022. "Classification and Identification of S Haplotypes in Radish Based on SRK Kinase Domain Sequence Analysis" Plants 11, no. 17: 2304. https://doi.org/10.3390/plants11172304