
Phylogenomics and Systematics of Overlooked Mesoamerican and South American Polyploid Broad-Leaved Festuca Grasses Differentiate F. sects. Glabricarpae and Ruprechtia and F. subgen. Asperifolia, Erosiflorae, Mallopetalon and Coironhuecu (subgen. nov.)
 and
and
Abstract
:1. Introduction
2. Results
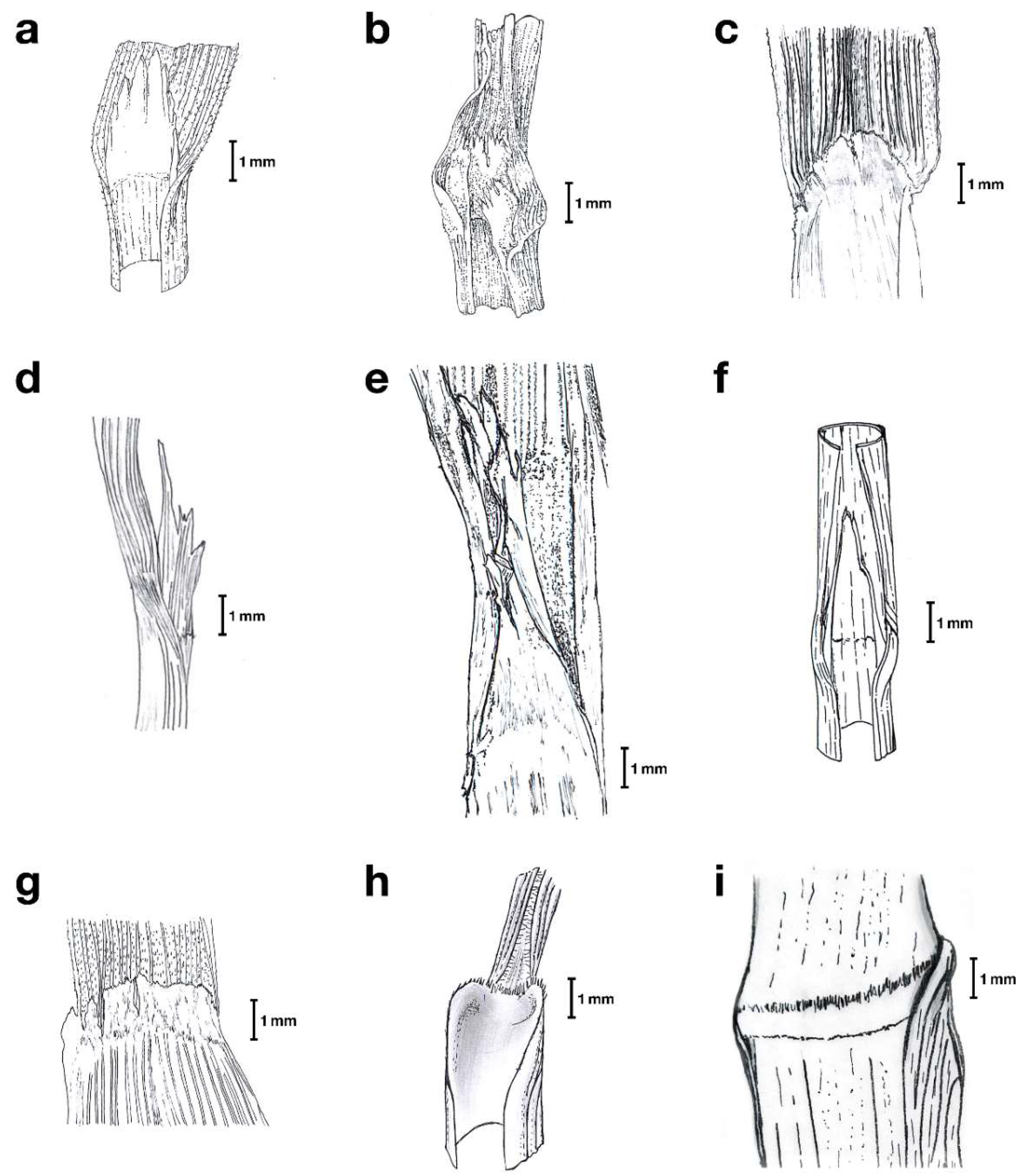
2.1. Taxonomic Study
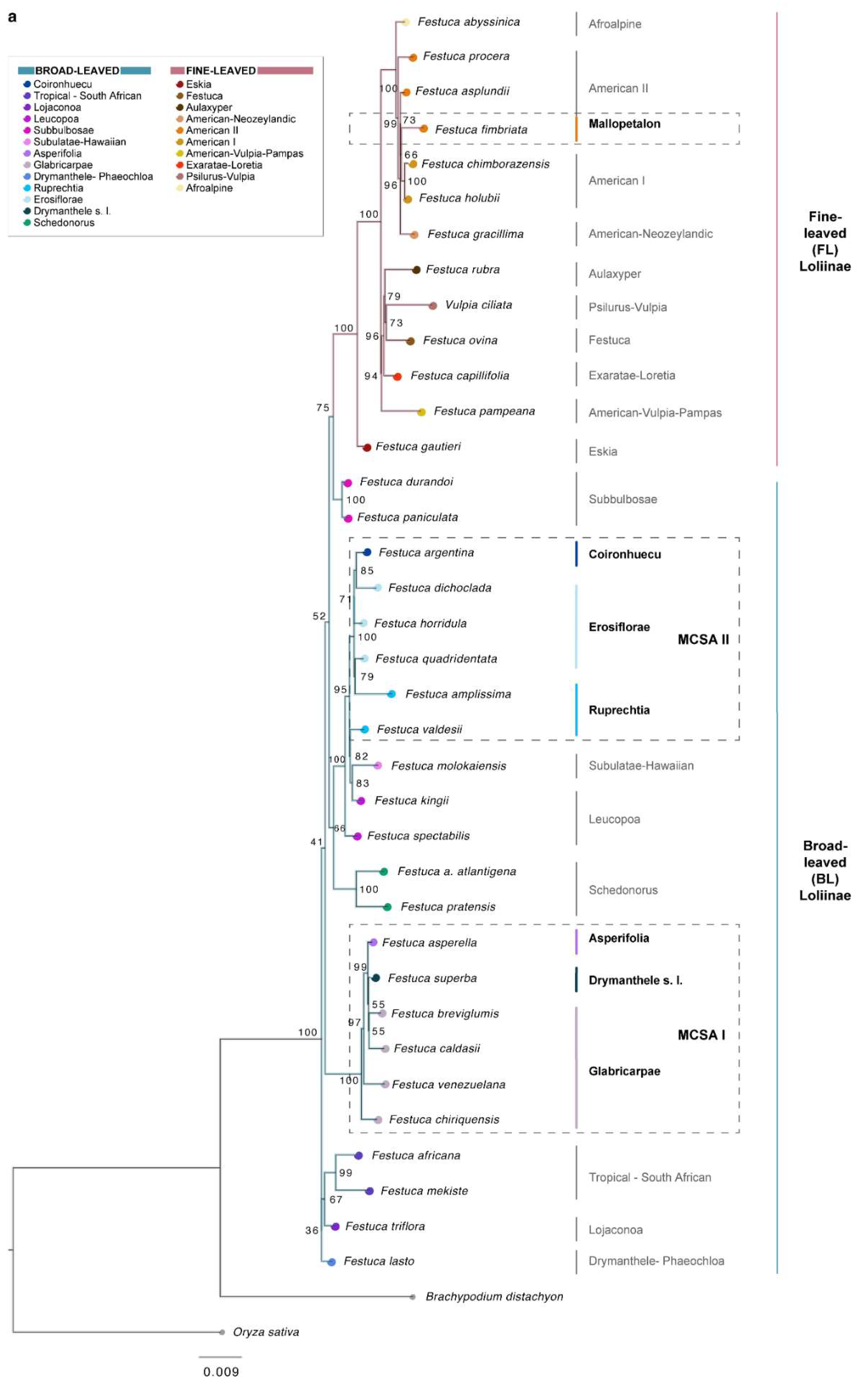
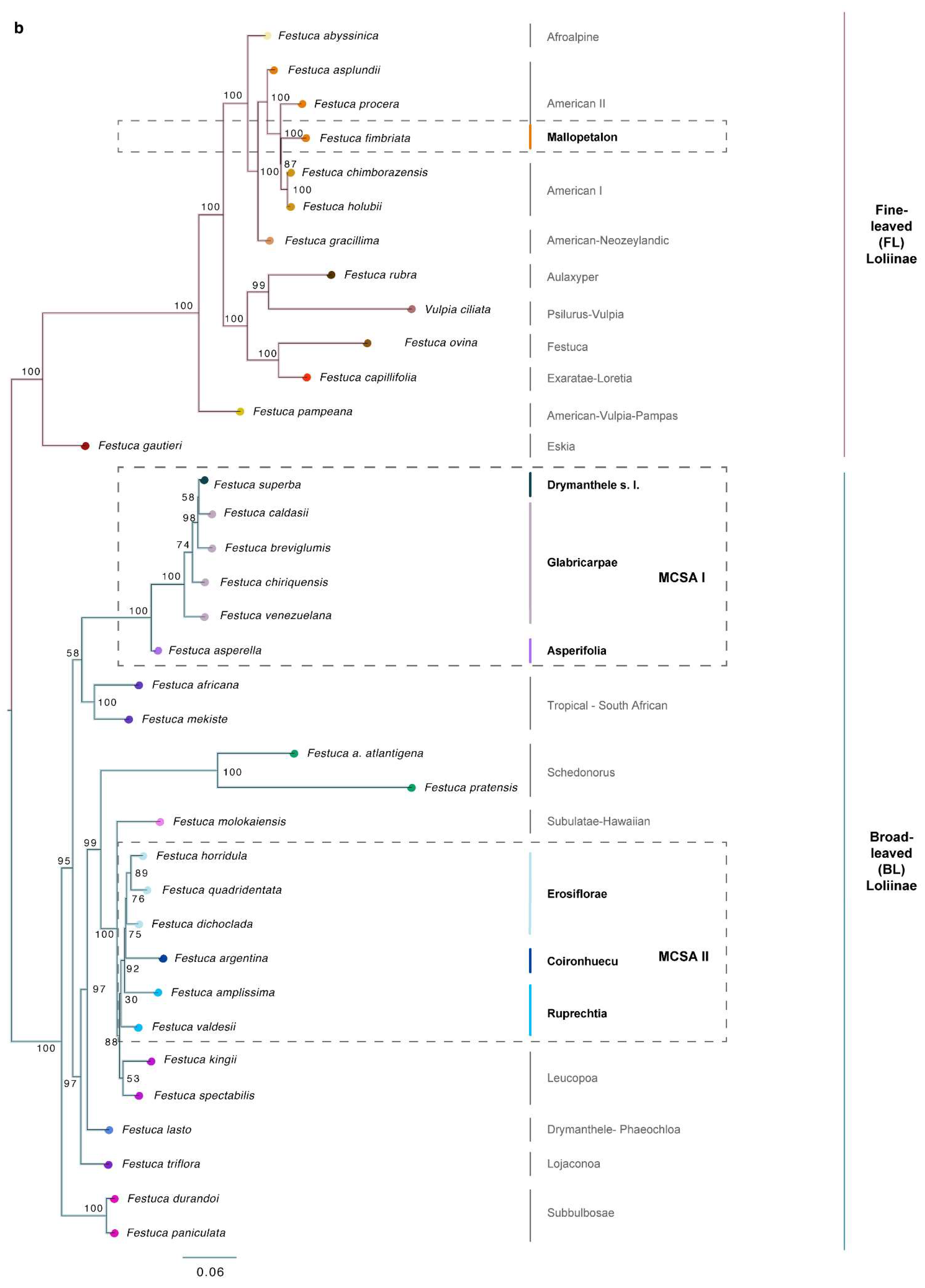
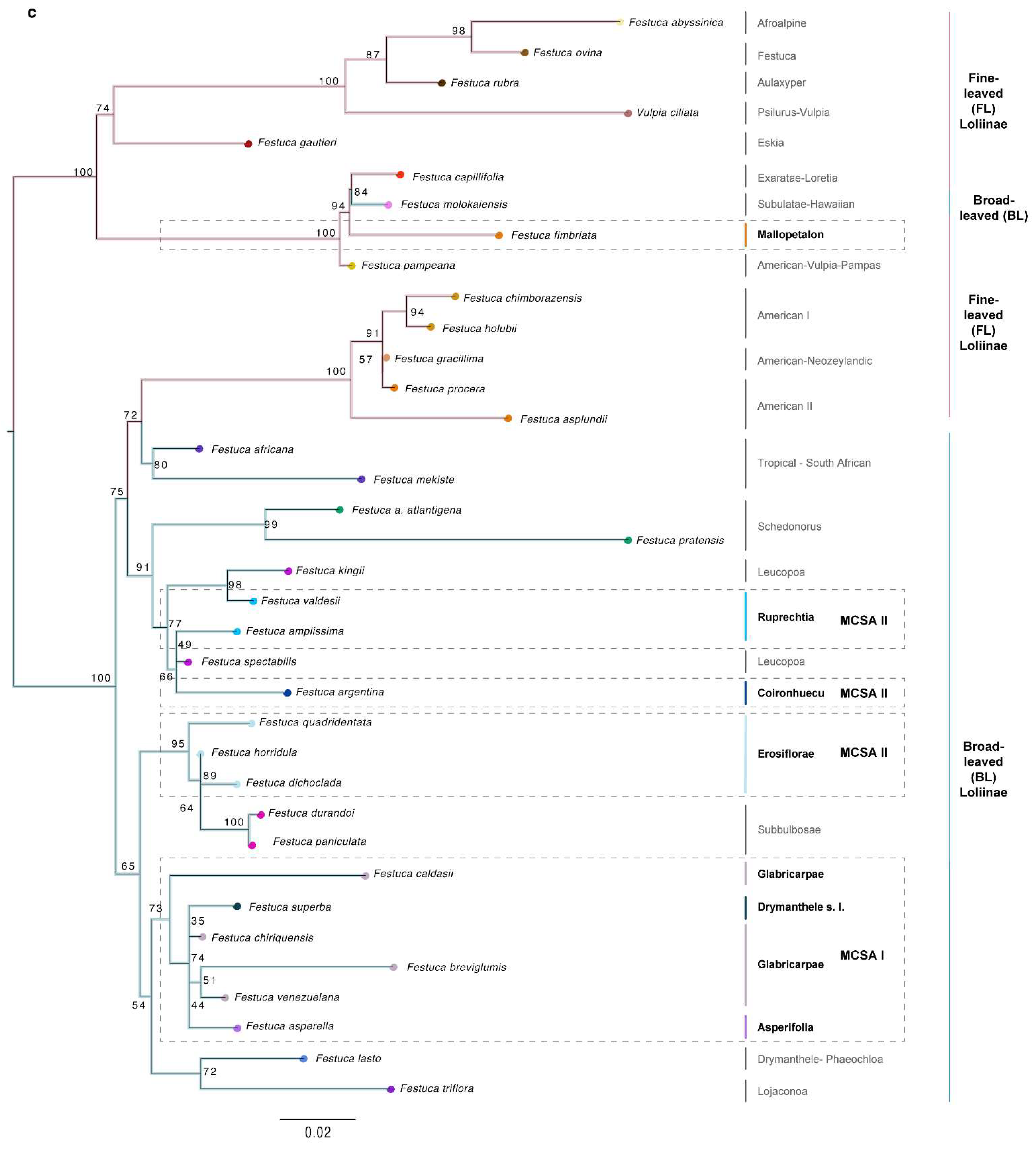
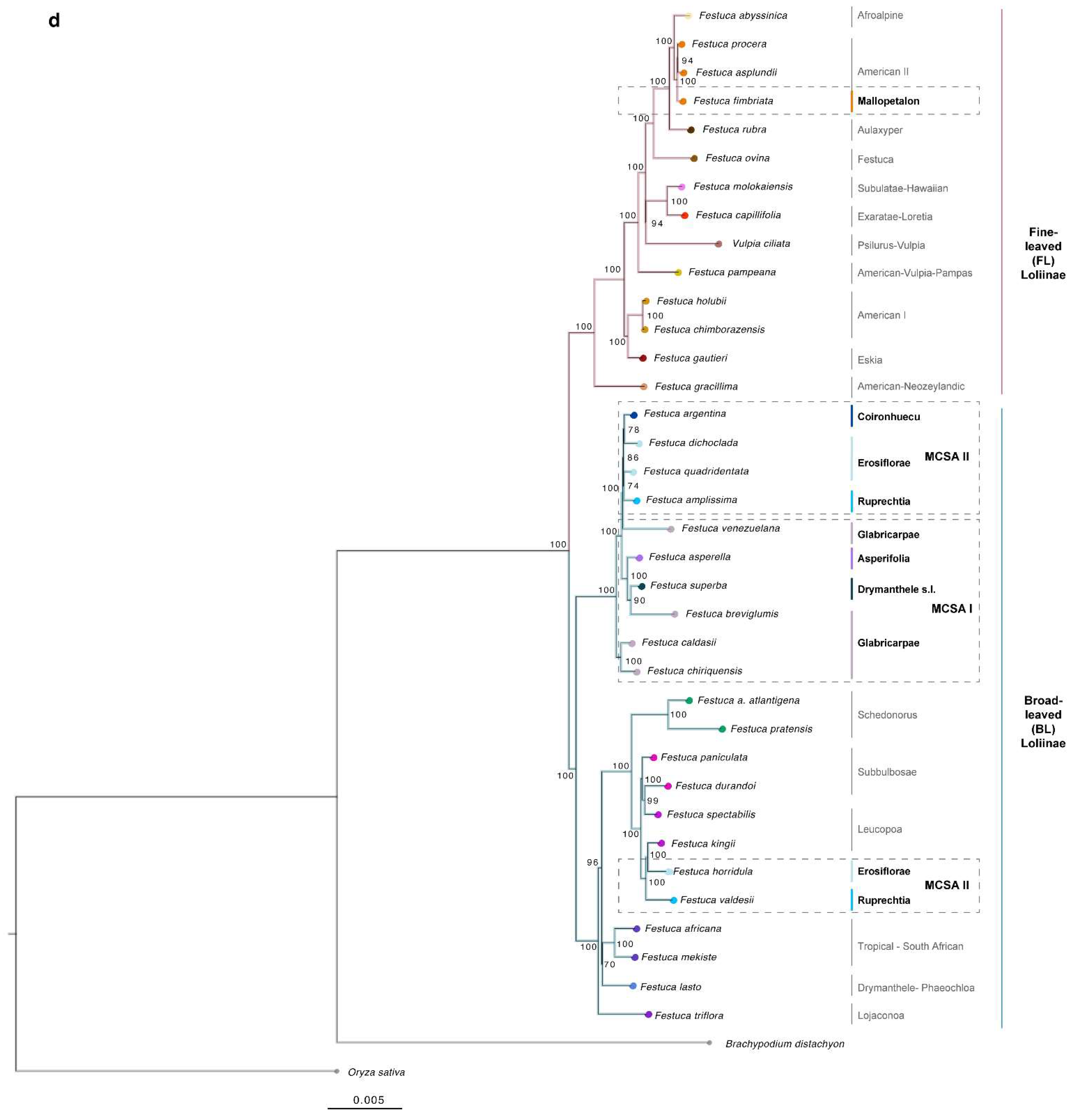
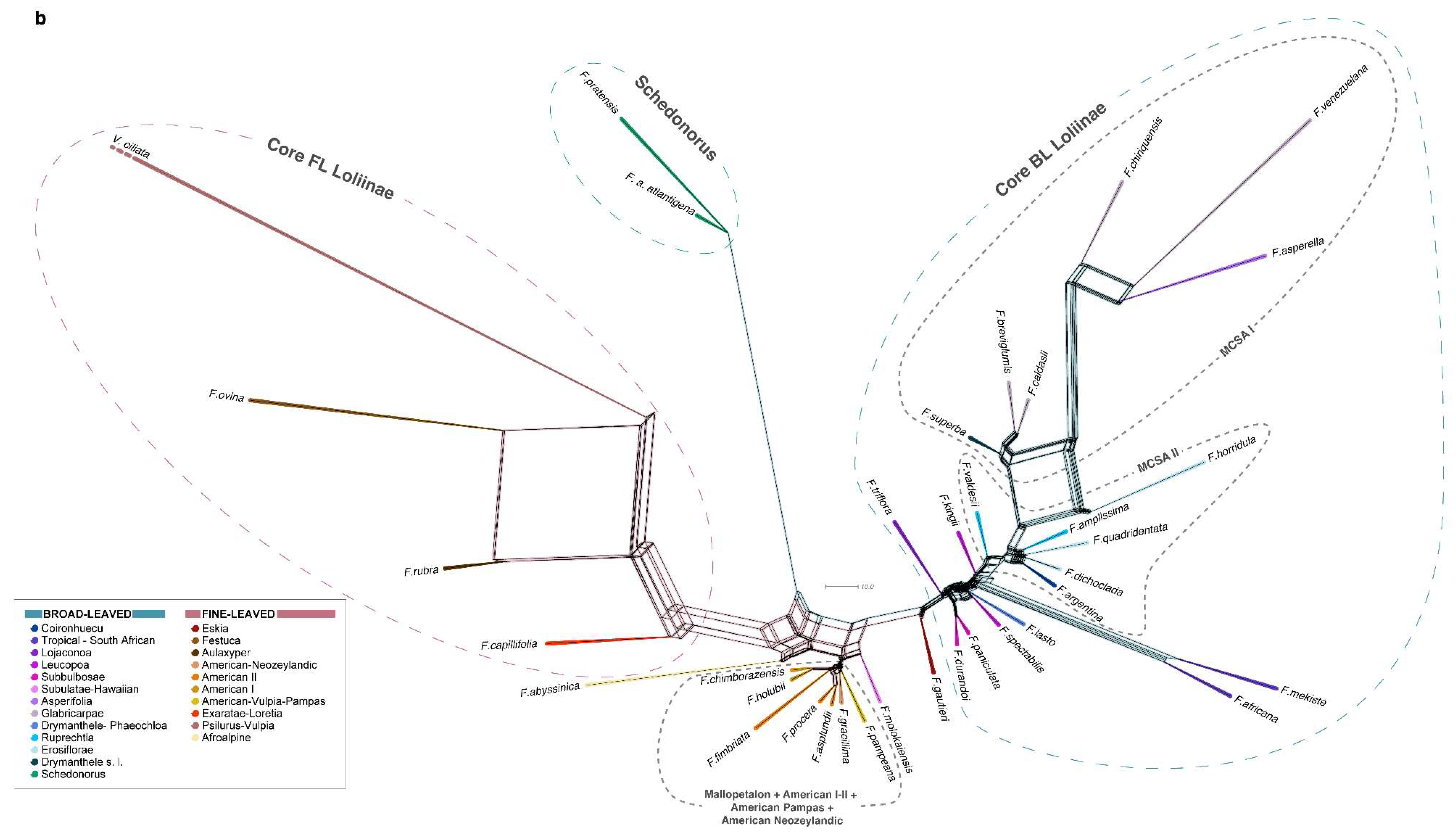
2.2. Phylogenomic Analyses
3. Discussion
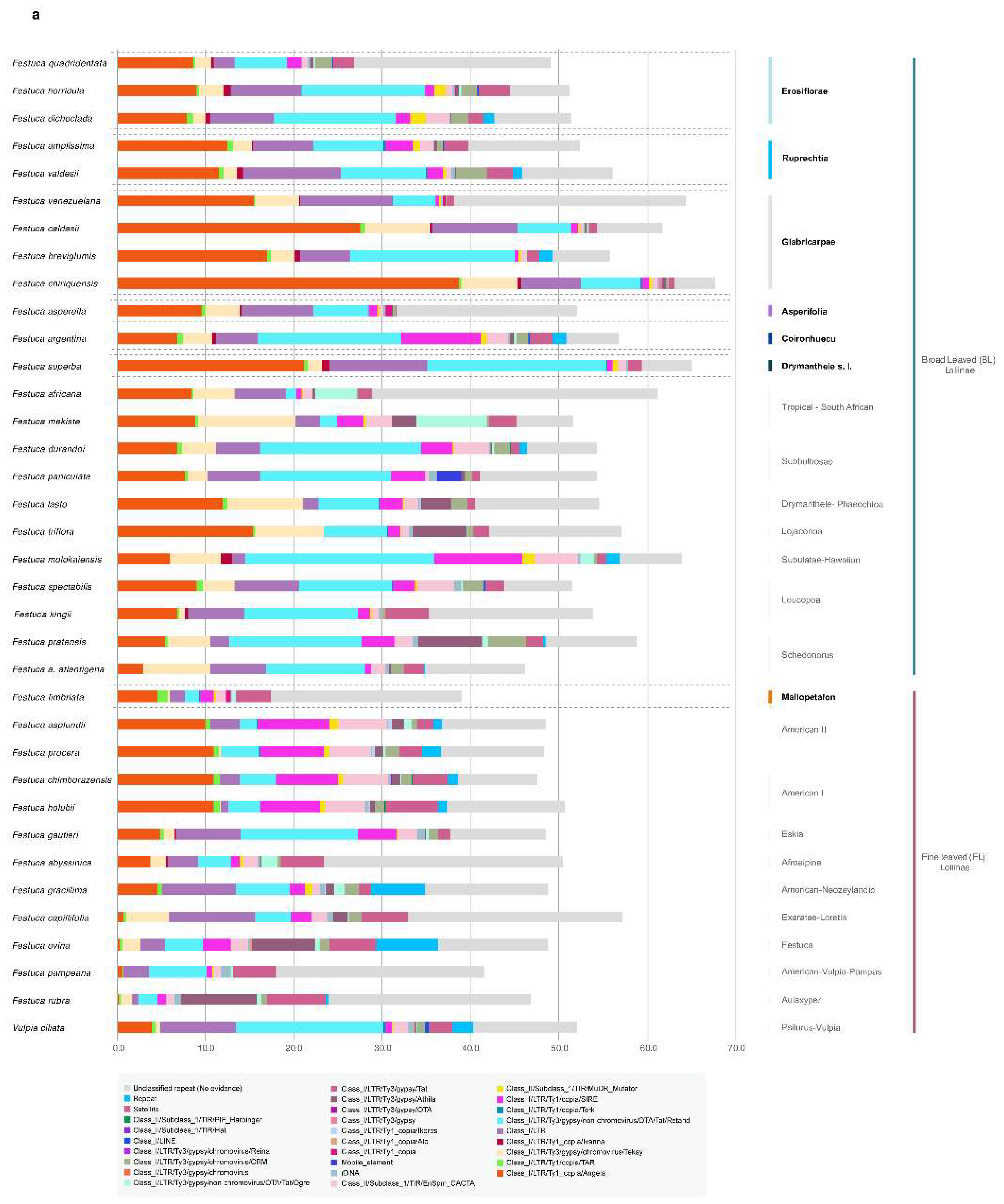
3.1. Evolutionary History of Allopolyploid Broad-Leaved Mexico–Central–South American Festuca Lineages (Erosiflorae, Ruprechtia, Glabricarpae, Asperella, Mallopetalon, F. argentina, F. superba)
3.2. Systematics of Broad-Leaved MCSA and Mallopetalon Loliinae Taxa
3.3. Description of Festuca subgen. Coironhuecu subgen. nov.
4. Material and Methods
4.1. Morphological Study of Herbarium Festuca Specimens
4.2. DNA Sampling of Festuca Species, Genome Sequencing, Data Assembling and Phylogenomic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Mayrose, I.; Zhan, S.H.; Rothfels, C.J.; Magnuson-Ford, K.; Barker, M.S.; Rieseberg, L.H.; Otto, S.P. Recently formed polyploid plants diversify at lower rates. Science 2011, 333, 1257. [Google Scholar] [CrossRef] [PubMed]
- Otto, S.P. The Evolutionary Consequences of Polyploidy. Cell 2007, 131, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Van De Peer, Y.; Maere, S.; Meyer, A. The evolutionary significance of ancient genome duplications. Nat. Rev. Genet. 2009, 10, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Visger, C.J.; Soltis, P.S. The polyploidy revolution then and now: Stebbins revisited. Am. J. Bot. 2014, 101, 1057–1078. [Google Scholar] [CrossRef]
- Soltis, D.E.; Albert, V.A.; Leebens-Mack, J.; Bell, C.D.; Paterson, A.H.; Zheng, C.; Sankoff, D.; DePamphilis, C.W.; Wall, P.K.; Soltis, P.S. Polyploidy and angiosperm diversification. Am. J. Bot. 2009, 96, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.E.; McKain, M.R.; McNeal, J.; Rolf, M.; Ruzicka, D.R.; Wafula, E.; Wickett, N.J.; et al. A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef]
- Stebbins, G.L. The evolutionary significance of natural and artificial polyploids in the family Gramineae. Hereditas 1949, 35, 461–485. [Google Scholar] [CrossRef]
- Kellogg, E.A. The Families and Genera of Vascular Plants. Vol. XIII. Flowering Plants. Monocots. Poaceae; Kubitzki, K., Ed.; Springer: New York, NY, USA, 2015; ISBN 978-3-319-15332-2. [Google Scholar]
- Marcussen, T.; Heier, L.; Brysting, A.K.; Oxelman, B.; Jakobsen, K.S. From gene trees to a dated allopolyploid network: Insights from the angiosperm genus Viola (Violaceae). Syst. Biol. 2015, 64, 84–101. [Google Scholar] [CrossRef] [Green Version]
- Salse, J.; Bolot, S.; Throude, M.; Jouffe, V.; Piegu, B.; Quraishi, U.M.; Calcagno, T.; Cooke, R.; Delseny, M.; Feuillet, C. Identification and characterization of shared duplications between rice and wheat provide new insight into grass genome evolution. Plant Cell 2008, 20, 11–24. [Google Scholar] [CrossRef]
- Murat, F.; Xu, J.H.; Tannier, E.; Abrouk, M.; Guilhot, N.; Pont, C.; Messing, J.; Salse, J. Ancestral grass karyotype reconstruction unravels new mechanisms of genome shuffling as a source of plant evolution. Genome Res. 2010, 20, 1545–1557. [Google Scholar] [CrossRef]
- Soltis, D.E.; Visger, C.J.; Blaine Marchant, D.; Soltis, P.S. Polyploidy: Pitfalls and paths to a paradigm. Am. J. Bot. 2016, 103, 1146–1166. [Google Scholar] [CrossRef]
- Stebbins, G.L. Polyploidy, hybridization and the invasion of new habitats. Ann. Missouri Bot. Gard. 1985, 72, 824–832. [Google Scholar] [CrossRef]
- Estep, M.C.; McKain, M.R.; Vela Diaz, D.; Zhong, J.; Hodge, J.G.; Hodkinson, T.R.; Layton, D.J.; Malcomber, S.T.; Pasquet, R.; Kellogg, E.A. Allopolyploidy, diversification, and the Miocene grassland expansion. Proc. Natl. Acad. Sci. USA 2014, 111, 15149–15154. [Google Scholar] [CrossRef] [PubMed]
- Mason-Gamer, R.J.; Burns, M.M.; Naum, M. Reticulate evolutionary history of a complex group of grasses: Phylogeny of Elymus StStHH allotetraploids based on three nuclear genes. PLoS ONE 2010, 5, e10989. [Google Scholar] [CrossRef] [PubMed]
- Catalán, P. Phylogeny and Evolution of Festuca L. and related genera of subtribe Loliinae (Poeae, Poaceae). In Plant Genome: Biodiversity and Evolution; Sharma, A.K., Ed.; Science Publishers: Enfield, UK, 2006; Volume 1, pp. 255–303. ISBN 9781578084203. [Google Scholar]
- Soreng, R.J.; Bull, R.D.; Gillespie, L.J. Phylogeny and Reticulation in Poa Based on Plastid trnTLF and nrITS Sequences with Attention to Diploids. Divers. Phylogeny Evol. Monocotyledons 2010, 619–644. [Google Scholar]
- Catalan, P.; López-Álvarez, D.; Díaz-Pérez, A.; Sancho, R.; López-Herránz, M.L. Phylogeny and Evolution of the Genus Brachypodium. In Genetics and Genomics of Brachypodium; John, P.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 18, pp. 9–38. ISBN 978-3-319-26942-9. [Google Scholar] [CrossRef]
- Sancho, R.; Inda, L.A.; Díaz-Pérez, A.; Des Marais, D.L.; Gordon, S.; Vogel, J.P.; Lusinska, J.; Hasterok, R.; Contreras-Moreira, B.; Catalán, P. Tracking the ancestry of known and ‘ghost’ homeologous subgenomes in model grass Brachypodium polyploids. Plant J. 2022, 109, 1535–1558. [Google Scholar] [CrossRef]
- Catalán, P.; Torrecilla, P.; López Rodríguez, J.Á.; Olmstead, R.G. Phylogeny of the festucoid grasses of subtribe Loliinae and allies (Poeae, Pooideae) inferred from ITS and trnL-F sequences. Mol. Phylogenet. Evol. 2004, 31, 517–541. [Google Scholar] [CrossRef]
- Catalán, P.; Torrecilla, P.; López-Rodríguez, J.; Müller, J.; Stace, C. A Systematic Approach to Subtribe Loliinae (Poaceae: Pooideae) Based on Phylogenetic Evidence. Aliso 2007, 23, 380–405. [Google Scholar] [CrossRef] [Green Version]
- Inda, L.A.; Segarra-Moragues, J.G.; Müller, J.; Peterson, P.M.; Catalán, P. Dated historical biogeography of the temperate Loliinae (Poaceae, Pooideae) grasses in the northern and southern hemispheres. Mol. Phylogenet. Evol. 2008, 46, 932–957. [Google Scholar] [CrossRef] [PubMed]
- Minaya, M.; Hackel, J.; Namaganda, M.; Brochmann, C.; Vorontsova, M.S.; Besnard, G.; Catalán, P. Contrasting dispersal histories of broad- and fine-leaved temperate Loliinae grasses: Range expansion, founder events, and the roles of distance and barriers. J. Biogeogr. 2017, 44, 1980–1993. [Google Scholar] [CrossRef]
- Moreno-Aguilar, M.F.; Arnelas, I.; Sánchez-Rodríguez, A.; Viruel, J.; Catalán, P. Museomics Unveil the Phylogeny and Biogeography of the Neglected Juan Fernandez Archipelago Megalachne and Podophorus Endemic Grasses and Their Connection with Relict Pampean-Ventanian Fescues. Front. Plant Sci. 2020, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Aguilar, M.F.; Inda, L.A.; Sánchez-Rodríguez, A.; Arnelas, I.; Catalán, P. Evolutionary Dynamics of the Repeatome Explains Contrasting Differences in Genome Sizes and Hybrid and Polyploid Origins of Grass Loliinae Lineages. Front. Plant Sci. 2022, 13, 901733. [Google Scholar] [CrossRef]
- Torrecilla, P.; Catalan, P. Phylogeny of Broad-leaved and Fine-leaved Festuca Lineages (Poaceae) based on Nuclear ITS Sequences. Syst. Bot. 2002, 27, 241–251. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Martínez, A. Distribución geográfica de los niveles de ploidía en Festuca. Parodiana 1992, 7, 91–99. [Google Scholar]
- Tovar, O. Revisión de las Especies Peruanas del Género Festuca, Gramineae; Memorias del Museo de Historia Natural “Javier Prado”; Universidad Nacional Mayor de San Marcos, Dirección Universitaria de Biblioteca y Publicaciones: Lima, Peru, 1972; pp. 1–93. [Google Scholar]
- Stančik, D.; Peterson, P.M. A revision of Festuca (Poaceae: Loliinae) in South American paramos. Contrib. USA Natl. Herb. 2007, 56, 1–184. [Google Scholar]
- Catalán, P.; Muller, J.; Festuca, L. Flora Argentina; Instituto de Botánica Darwinion: Buenos Aires, Argentina, 2012; Volume 3, pp. 219–250. [Google Scholar]
- Ospina, J.C.; Aliscioni, S.S.; Denham, S.S. A Revision of Festuca (Loliinae, Pooideae, Poaceae) in Chile. Phytotaxa 2015, 223, 1–66. [Google Scholar] [CrossRef]
- Ospina, J.C. Estudios Morfológicos, Anatómicos, Taxonómicos y Relaciones Filogenéticas de las Especies de Festuca del Cono Sur (Argentina, Chile, Uruguay, Paraguay y Sur de Brasil). Ph.D. Thesis, Universidad Nacional de La Plata, La Plata, Argentina, 2016; pp. 1–229. [Google Scholar]
- Alexeev, E. To the systematics of Asian Fescues (Festuca subgenera Drymanthele, Subulatae, Schedonorus, Leucopoa). Byull. Mosk. Obs. Isp. Prir. Otd. Biol. 1977, 82, 95–102. [Google Scholar]
- Alexeev, E.B. Sistematike asiatskich ovsjaniz (Festuca). II. Podrod Festuca. Byull. Mosk. Obs. Isp. Prir. Otd. Biol. 1978, 83, 109–122. [Google Scholar]
- Alexeev, E.B. Festuca, L. Subgenera et Sectiones novae ex America boreali et Mex. Nov. Sist. Vyss. Nizsh. Rast. 1980, 17, 42–53. [Google Scholar]
- Alexeev, E.B. The new taxa of the Festuca (Poaceae) from México and Central America. Bot. Zhurn. 1981, 66, 1492–1501. [Google Scholar]
- Alexeev, E.B. Genus Festuca L. (Poaceae) in Mexico et America Centrali. Nov. Sist. Vyss. Rast. 1984, 21, 25–59. [Google Scholar]
- Alexeev, E.B. Novye rody slakov. Byull. Mosk. Obs. Isp. Prir. Otd. Biol. 1985, 90, 102–109. [Google Scholar]
- Alexeev, E.B. New taxa and typification of Festuca (Poaceae) of Bolivia. Bot. Zhurn. 1985, 70, 1241–1247. [Google Scholar]
- Alexeev, E.B. Festuca L. (Poaceae) in Venezuela, Colombia & Ecuador. Nov. Sist. Vyss. Nizsh. Rast. 1986, 23, 5–23. [Google Scholar]
- Alexeev, E.B. A new section and three new species of the genus Festuca (Poaceae) from México and Central America. Bot. Zhurn. 1982, 67, 1289–1292. [Google Scholar]
- Alexeev, E.B. On the new taxa and typification of some taxa of the genus Festuca (Poaceae) from South America. Bot. Zhurn. 1984, 69, 346–353. [Google Scholar]
- Kopecký, D.; Studer, B. Emerging technologies advancing forage and turf grass genomics. Biotechnol. Adv. 2014, 32, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Saint-Yves, A. 1927. Contribution a l’étude des Festuca (subg. Eu-Festuca) de l’Amérique du Sud. Candollea 1927, 3, 151–315. [Google Scholar]
- Dubcovsky, J. Estudios Citogenéticos y Evolutivos de las Especies Patagónicas del Género Festuca (Poaceae). Ph.D. Thesis, Universidad de Buenos Aires, Facultad de Ciencias Exactas y Naturales, Buenos Aires, Argentina, 1989; pp. 1–134. [Google Scholar]
- Stančík, D.; Renvoize, S. Two new and two little-known species of Festuca (Poaceae) from the Bolivian Andes. Kew Bull. 2007, 62, 485–492. [Google Scholar]
- Türpe, A. Las especies argentinas de Festuca. Darwiniana 1969, 15, 189–283. [Google Scholar]
- Stančík, D.; Peterson, P.M. Festuca dentiflora (Poaceae: Loliinae: Sect. Glabricarpae), a new species from Peru and taxonomic status of F. presliana. SIDA Contrib. Bot. 2003, 20, 1015–1022. [Google Scholar]
- Stebbins, G. Chromosomal Evolution in Higher Plants; Edward Arnold Publication: London, UK, 1971. [Google Scholar] [CrossRef]
- Pires, J.C.; Lim, K.Y.; Kovarik, A.; Matyasek, R.; Boyd, A.; Leitch, A.R.; Leitch, I.J.; Bennett, M.D.; Soltis, P.S.; Soltis, D.E. Molecular Cytogenetic Analysis of Recently Reveal a Karyotype That Is Additive of the diploid progenitors. Amer. J. Bot. 2004, 91, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- González-Ledesma, M.; Koch, S.; Gómez-Sánchez, M. Two new species of Festuca (Gramineae: Pooideae) from Mexico. Novon 1998, 8, 147–151. [Google Scholar] [CrossRef]
- Frajman, B.; Schönswetter, P.; Weiss-Schneeweiss, H.; Oxelman, B. Origin and Diversification of South American Polyploid Silene Sect. Physolychnis (Caryophyllaceae) in the Andes and Patagonia. Front. Genet. 2018, 9, 639. [Google Scholar] [CrossRef]
- Tzvelev, N. On the taxonomy and phylogeny of genus Festuca L. of the U.S.R.R. flora. I. The system of the genus and main trends of evolution. Bot. Zhurn. 1971, 56, 1252–1262. [Google Scholar]
- Lu, S.; Chen, X.; Aiken, S. Festuca Linnaeus. Flora China 2006, 22, 225–242. [Google Scholar]
- Giussani, L.M.; Gillespie, L.J.; Scataglini, M.A.; Negritto, M.A.; Anton, A.M.; Soreng, R.J. Breeding system diversification and evolution in American Poa supersect. Homalopoa (Poaceae: Poeae: Poinae). Ann. Bot. 2016, 118, 281–303. [Google Scholar] [CrossRef] [PubMed]
- Toups, M.A.; Vicoso, B.; Pannell, J.R. Dioecy and chromosomal sex determination are maintained through allopolyploid speciation in the plant genus Mercurialis. PLoS Genet. 2022, 18, e1010226. [Google Scholar] [CrossRef]
- Lu, S. New taxa of Festuca L. from China. Acta Phytotax. Sin. 1992, 30, 529–540. [Google Scholar]
- Alexeev, E.B. Viviparous Fescues (Festuca L., Gramineae) of South America. Bjull. Mock. Izpit. Prirod. 1984, 89, 111–117. [Google Scholar]
- Saint-Yves, A. Tentamen. Claves analyticae Festucarum veteris orbis (subgen. Eu-Festucarum) ad subspecies, multas varietates et nonullas subvarietates usque ducentes. Rev. Bretonne Bot. Pure Appl. 1927, 3, 151–315. [Google Scholar]
- Tovar, O. Las Gramíneas (Poaceae) del Perú. Ruizia 1993, 13, 1–480. [Google Scholar]
- Doyle, J.; Doyle, J. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef]
- Garcia, S.; Wendel, J.F.; Borowska-Zuchowska, N.; Aïnouche, M.; Kuderova, A.; Kovarik, A. The Utility of Graph Clustering of 5S Ribosomal DNA Homoeologs in Plant Allopolyploids, Homoploid Hybrids, and Cryptic Introgressants. Front. Plant Sci. 2020, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Macas, J. Global analysis of repetitive DNA from unassembled sequence reads using RepeatExplorer2. Nat. Protoc. 2020, 15, 3745–3776. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Swofford, D.L.; Sullivan, J. Phylogeny inference based on parsimony and other methods using PAUP*. In The Phylogenetic Handbook: A Practical Approach to DNA and Protein Phylogeny, Cáp, 7; Cambridge University Press: Cambridge, UK, 2003; pp. 160–206. [Google Scholar]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Festuca Subgenera, Sections, Species/Morphological Diagnostic Traits | Subgen. Erosiflorae sensu Alexeev: F. dichoclada * F. horridula * F. quadridentata* sensu Stančík & Peterson: F. carrascana F. chuquisacae F. urubambana | Subgen. Drymanthele Sect. Ruprechtia sensu Alexeev: F. amplissima* F. jaliscana sensu Gonzalez-Ledesma et al.: F. valdesii * | Subgen. Subulatae Sect. Glabricarpae sensu Alexeev: F. breviglumis* F. chiriquensis* F. dentiflora F. steinbachii F. caldasii* F. woodii This Study: F. venezuelana * | Subgen. Asperifolia sensu Alexeev: F. lugens F. asperella * F. tancitaroensis | Subgen. Mallopetalon sensu Alexeev: F. fimbriata * | Subgen. Coironhuecu subgen. nov. This Study: F. argentina * | Subgen. Drymanthele sensu lato (Without Sectional Assignation) This Study: F. superba * |
|---|---|---|---|---|---|---|---|
| Reproduction | Monoecious | Monoecious | Monoecious | Monoecious | Monoecious | Dioecious | Monoecious |
| Habit | Largely tussocked or rhizomatous or mixed | Rhizomatous or caespitose | Rhizomatous or loosely tufted | Densely tussocked or rhizomatous | Rhizomatose | Caespitose | Laxely caespitose to rhizomatose |
| Innovations | Extravaginal or intravaginal | Extravaginal or/and intravaginal | Extravaginal | Extravaginal or intravaginal | Extravaginal | Intravaginal | Mixed |
| Ligule | Membranaceous, apex acute, erose or lacerate, 5.5–21 mm long | Non-membranaceous, apex truncate shortly ciliate, or short membranaceous, apex truncate and ciliate; 0.1–0.5 (1) mm long | Membranceous or hyaline, apex truncate or rounded, lacerate or dentate; or shortly ciliate; 0.3–4 mm long | Membranaceous, apex truncate or slightly rounded and lacerate or dentate, 1.4–8 mm long | Membranaceous, apex truncate, erose and ciliate, 0.5–1.5 mm long | Membranaceous, apex truncate and densely ciliate, 0.4–1.5 mm long | Hyaline, apex truncate, erose and dentate, 2.7–5.5 mm long |
| Leaf blade | Flat, involute in the middle and subconvolute at the apex | Flat, involute in the middle and subconvolute at the apex | Flat, involute in the middle and subconvolute at the apex | Flat, involute in the middle and subconvolute at the apex | Largely flat | Plicate, junciform | Largely flat, subconvolute |
| Inflorescence | Erect | Erect | Nutant or erect with nutant branches | Erect or scarcely nutant | Erect, lax | Erect, contracted | Erect, branches flexuous |
| Lemma apex | Dentate or entire, unawned | Entire, unawned | Entire or bifid, awned | Bifid, shortly awned or unawned | Entire, scariose, rolled and fimbriate, unawned, muticous | Entire, unawned, muticous or mucronulate | Entire, unawned, muticous |
| Ovary tip | Glabrescent | Glabrous or hispid | Glabrous or sparsely hispid | Glabrous or hispid | Densely hairy | Sparsely hispid | Densely hairy |
| Taxon | Phylogenetic Lineage | Ploidy | Locality/Voucher | Illumina PE Reads (Millions) | GenBank Accession No. | |||
|---|---|---|---|---|---|---|---|---|
| 35S | IGS | 5S | Plastome | |||||
| Broad-Leaved (BL) Loliinae | ||||||||
| Festuca asperella E.B. Alexeev | Asperifolia (MCSAI) | ? | Mexico: Mexico DF; MO 2744225 | 5683 | OP120918 | OP158136 | OP142677 | SAMN30029288 |
| Festuca breviglumis Swallen | Glabicarpae | ? | Mexico: Mexico DF; P, M, Peterson 21366; US s.n | 12,181 | OP120919 | OP158139 | OP142678 | SAMN30029289 |
| Festuca caldasii (Kunth) Kunth | Glabicarpae (MCSAI) | 4× | Ecuador: Catamayo, Chinchas-Tambara; HUTPL14055 | 9863 | MT145280 | OP158140 | ON248977 | SAMN14647047 |
| Festuca chiriquensis Swallen | Glabicarpae (MCSAI) | 4× | Costa Rica: Cartago, Cantón Turrialba; MO 5175763 | 8653 | OP120920 | OP158143 | OP142679 | SAMN30029290 |
| Festuca superba Parodi ex Türpe | Drymanthele s. l. (MCSAI) | 8× | Argentina: Jujuy, Yala, Laguna Rodeo; PC 356.08 UZ | 12,193 | MT145305 | OP158163 | ON248977 | SAMN14647072 |
| Festuca venezuelana Stančík | Glabicarpae (MCSAI) | 6× | Venezuela: Tachira, La Grita; AAU-4262 | 7957 | OP120926 | OP158166 | OP142686 | SAMN30029296 |
| Festuca dichoclada Pilg. | Erosiflorae (MCSAII) | ? | Peru: Cuzco, Quispicanchi; P, M, Peterson 20603; US s.n. | 12,466 | OP120921 | OP158144 | OP142680 | SAMN30029291 |
| Festuca horridula Pilg. | Erosiflorae (MCSAII) | ? | Peru: Junín, Yauli; Tovar, O, and H, Soplín 6607 | 32,417 | OP120923 | OP158150 | OP142682 | SAMN30029293 |
| Festuca quadridentata Kunth | Erosiflorae (MCSAII) | ? | Ecuador: Chimborazo, Alao; US 1911313 | 15,091 | MT145303 | OP158160 | OP142684 | SAMN14647070 |
| Festuca amplissima Rupr. | Ruprechtia (MCSAII) | 6× | Mexico: Nuevo Leon; Peterson 21097, US s.n. | 12,058 | MT145278 | OP158134 | ON248975 | SAMN14647045 |
| Festuca valdesii Gonz.-Led. & S.D. Koch | Ruprechtia (MCSAII) | ? | Mexico: Coahuila; P, M, Peterson 21456; US s.n. | 10,937 | OP120925 | OP158165 | OP142685 | SAMN30029295 |
| Festuca argentina (Speg.) Parodi | Coironhuecu (MCSAII) | 4× | Argentina: Rio Negro, Bariloche; PC, 0210 | 22,928 | OP120917 | OP158135 | OP142676 | SAMN30029287 |
| Festuca kingii (S. Watson) Cassidy | Leucopoa | 8× | USA: California: San Bernardino Mnts, Leg: Quibell 149; LE | 12,397 | OP120924 | OP158151 | OP142683 | SAMN30029294 |
| Festuca spectabilis Bertol. | Leucopoa | 6× | Bosnia-Hercegovina: Troglav, Sajkovacko zdrlo, UZ | 12,960 | MT145304 | OP158162 | ON249004 | SAMN14647071 |
| Festuca africana (Hack.) Clayton | Tropical and South African | 10× | Uganda: Gahinga; Namaganda 190Vg; MHU1603 | 13,549 | MT145277 | OP158133 | ON248974 | SAMN14647044 |
| Festuca mekiste Clayton | Tropical and South African | ? | Kenya: Mt, Elgon National Park, Kambi Mtamaiwa; Carvalho 4521 | 16,245 | ON243855 | OP158153 | ON248992 | SAMN27777779 |
| Festuca durandoi Clauson | Subbulbosae | 2× | Portugal: Serra Arga Alto do Espinheiro; UZ s.n. | 12,688 | MT145283 | OP158145 | ON248980 | SAMN14647050 |
| Festuca paniculata (L.) Schinz & Thell | Subbulbosae | 2× | Spain: Caceres, Puerto de los Castaños; UZ 40.07 | 35,808 | MT145297 | OP158157 | ON248996 | SAMN14647064 |
| Festuca triflora J.F. Gmel. | Lojaconoa | 2× | Morocco: Rif Mountains, Bab Barret-Ketama; PC 39.17 UZ | 24,472 | MT145306 | OP158164 | ON249006 | SAMN14647073 |
| Festuca lasto Boiss. | Drymanthele (Phaeochloa) | 2× | Spain: Cadiz, Los Alcornocales; UZ 29.08 | 21,581 | MT145291 | OP158152 | ON248989 | SAMN14647058 |
| Festuca pratensis Huds. | Schedonorus | 2× | UK: England; USDA PI 283306 | 12,189 | MT145301 | OP158158 | ON248998 | SAMN14647066 |
| Festuca arundinacea subsp. atlantigena (St.-Yves) Auquier | Schedonorus | 8× | Morocco: Atlas mountains; ABY BN 807 | 15,091 | ON243851 | OP158138 | ON248990 | SAMN27777775 |
| Festuca molokaiensis Soreng, P.M. Peterson & Catalán | Subulatae-Hawaiian | ? | USA: Hawaii: Molokai, BISH 728771 | 12,188 | MT145294 | OP158154 | ON248993 | SAMN14647061 |
| Fine-leaved (FL) Loliinae | ||||||||
| Festuca fimbriata Nees | American II | 6× | Argentina: Misiones, Dpto, Apóstoles; UZ 498.08 | 15,741 | MT145286 | OP158146 | ON248983 | SAMN14647053 |
| Festuca asplundii E.B. Alexeev | American II | 6× | Ecuador: Loja, Saraguro; HUTPL14046 | 25,088 | MT145279 | OP158137 | ON248976 | SAMN14647046 |
| Festuca procera Kunth | American II | ? | Ecuador: Riobamba, Chimborazo; HUTPL14079 | 40,669 | MT145299 | OP158159 | ON248999 | SAMN14647067 |
| Festuca chimborazensis E.B. Alexeev | American I | 6× | Ecuador: Riobamba, Chimborazo; HUTPL14066 | 10,913 | MT145282 | OP158142 | ON248979 | SAMN14647049 |
| Festuca holubii Stančík | American I | ? | Ecuador: Saraguro, to Cerro de Arcos; HUTPL14071 | 10,264 | MT145289 | OP158149 | ON248988 | SAMN14647056 |
| Festuca pampeana Speg. | American Pampas | 8× | Argentina: Buenos Aires, Sierra de la Ventana; PC 428.08 | 14,862 | MT145296 | OP158156 | ON248995 | SAMN14647063 |
| Festuca gracillima Hook. f. | American–Neozeylandic I | 6× | Argentina: Tierra de Fuego, E. San Pablo; UZ 482.08 | 13,888 | MT145288 | OP158148 | ON248986 | SAMN14647055 |
| Festuca abyssinica Hochst. ex A. Rich. | Afroalpine | 4× | Tanzania: Kilimanjaro; Afroalp O-DP-42737 | 12,041 | MT145276 | OP158132 | ON248973 | SAMN14647043 |
| Festuca rubra L. | Aulaxyper | 6× | Argentina: Tierra de Fuego, Cabo Annicolta; UZ 03.09 | 25,260 | ON243856 | OP158161 | ON249002 | SAMN27777780 |
| Festuca ovina L. | Festuca | 2× | Germany: Thüringen; Müller 10789 | 11,364 | MT145295 | OP158155 | ON248994 | SAMN14647062 |
| Festuca capillifolia Dufour ex Roem. & Schult. | Exaratae | 2× | Morocco: Middle Atlas, Ifrane National Park; PC 77.17 | 13,430 | MT145281 | OP158141 | ON248978 | SAMN14647048 |
| Vulpia ciliata Dumort. | Psilurus–Vulpia | 4× | Spain: Toledo, Mar de Ontígola; UZ 109.07 | 11,801 | MT145309 | OP158167 | ON249009 | SAMN14647076 |
| Festuca gautieri (Hack.) K. Richt. | Eskia | 2× | Spain: Granada, Huéscar; UZ 232.07 | 13,941 | OP120922 | OP158147 | OP142681 | SAMN30029292 |
| Outgroups | ||||||||
| Brachypodium distachyon (L.) P. Beauv. | --- | 2× | Spain: Caceres; UZ 28.07 | --- | Phytozome Bd21 v.3.1 | --- | --- | NC_011032, 1 |
| Oryza sativa L. | --- | 2× | China: National Rice Research Center, cv | --- | AP008215 | --- | --- | AY522331, 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Aguilar, M.F.; Inda, L.A.; Sánchez-Rodríguez, A.; Catalán, P.; Arnelas, I. Phylogenomics and Systematics of Overlooked Mesoamerican and South American Polyploid Broad-Leaved Festuca Grasses Differentiate F. sects. Glabricarpae and Ruprechtia and F. subgen. Asperifolia, Erosiflorae, Mallopetalon and Coironhuecu (subgen. nov.). Plants 2022, 11, 2303. https://doi.org/10.3390/plants11172303
Moreno-Aguilar MF, Inda LA, Sánchez-Rodríguez A, Catalán P, Arnelas I. Phylogenomics and Systematics of Overlooked Mesoamerican and South American Polyploid Broad-Leaved Festuca Grasses Differentiate F. sects. Glabricarpae and Ruprechtia and F. subgen. Asperifolia, Erosiflorae, Mallopetalon and Coironhuecu (subgen. nov.). Plants. 2022; 11(17):2303. https://doi.org/10.3390/plants11172303
Chicago/Turabian StyleMoreno-Aguilar, María Fernanda, Luís. A. Inda, Aminael Sánchez-Rodríguez, Pilar Catalán, and Itziar Arnelas. 2022. "Phylogenomics and Systematics of Overlooked Mesoamerican and South American Polyploid Broad-Leaved Festuca Grasses Differentiate F. sects. Glabricarpae and Ruprechtia and F. subgen. Asperifolia, Erosiflorae, Mallopetalon and Coironhuecu (subgen. nov.)" Plants 11, no. 17: 2303. https://doi.org/10.3390/plants11172303