
Evidence for Light and Tissue Specific Regulation of Genes Involved in Fructan Metabolism in Agave tequilana

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Characterization of Genes Encoding PGHF32 Members in Agave tequilana
2.2. Identification of Regulatory Motifs in the Promoters of PGHF32
2.3. A. tequilana Transcriptome Database Searches
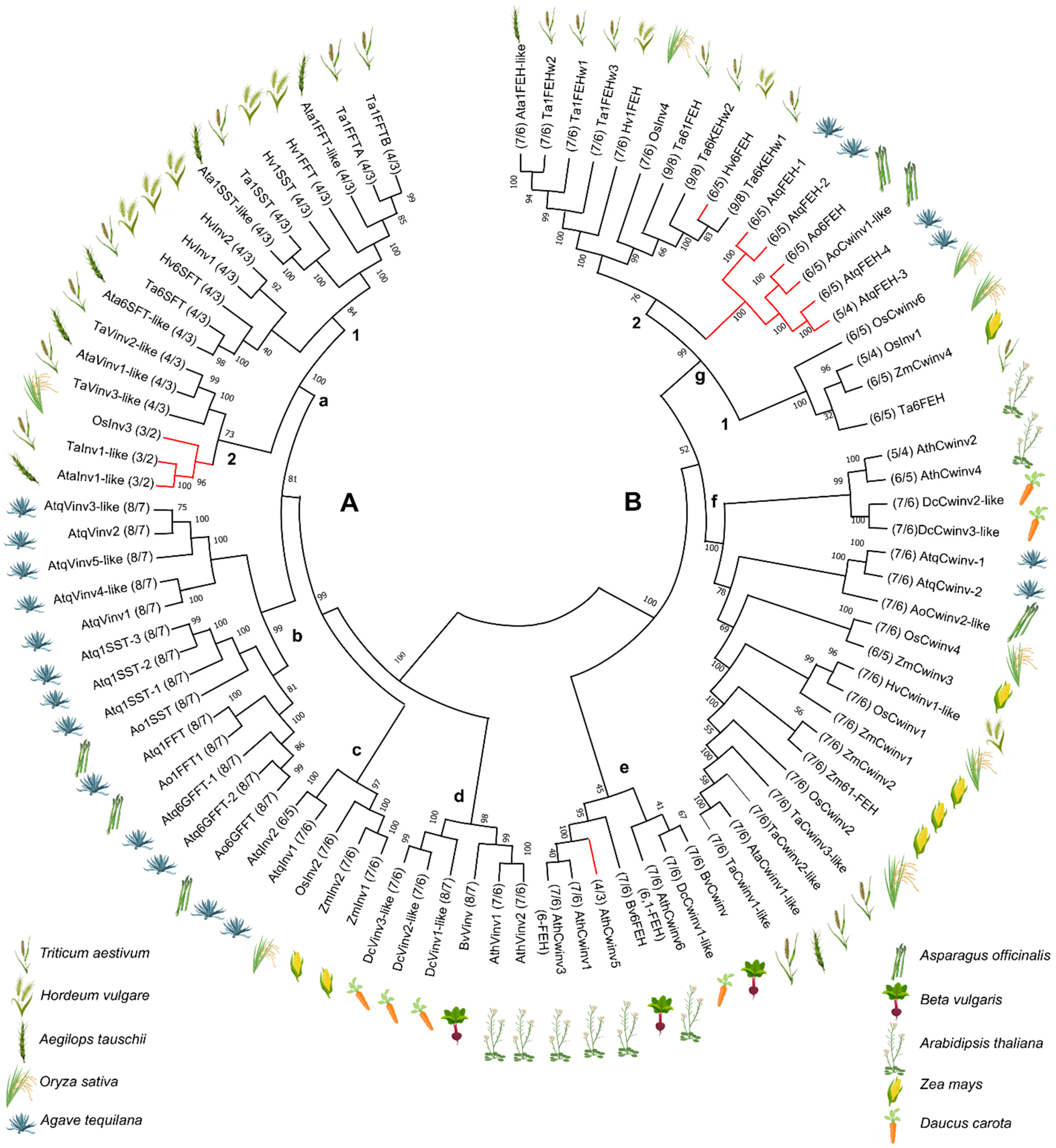
2.4. Alignment and Identification of Conserved Motifs
2.5. In Silico Expression Analysis
2.6. qRT-PCR Analysis
3. Results
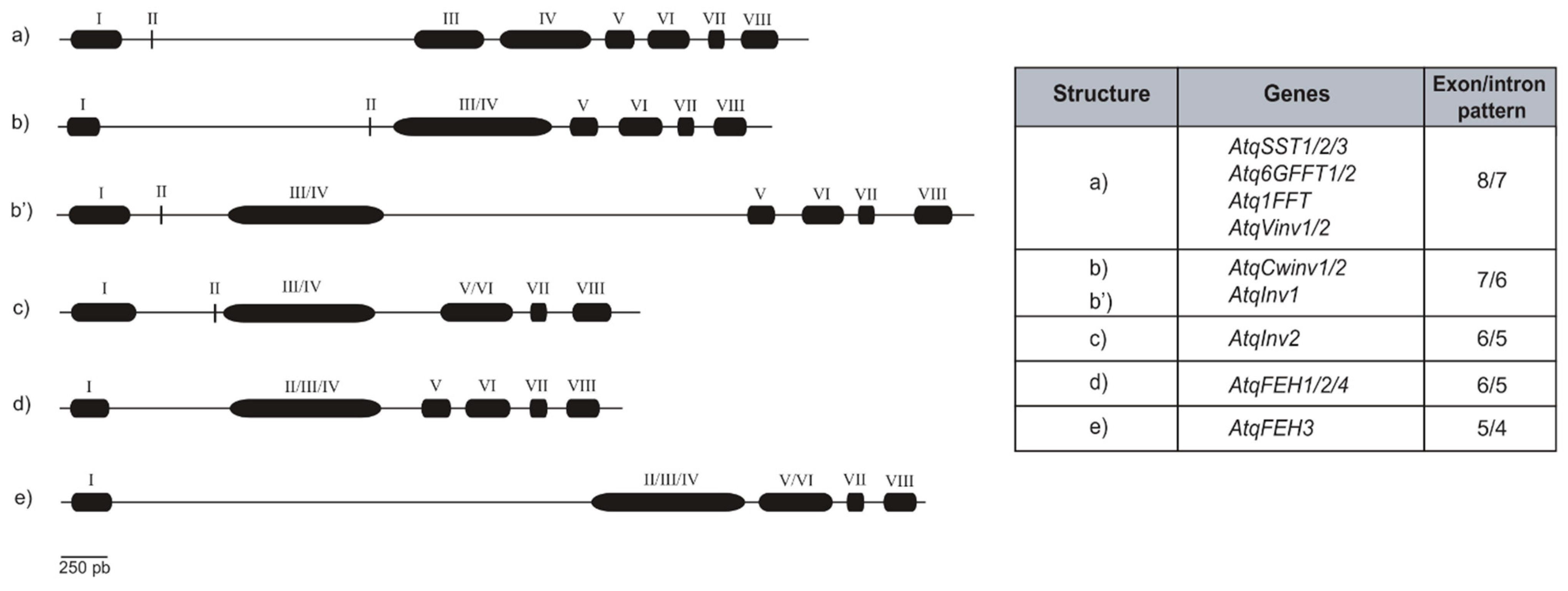
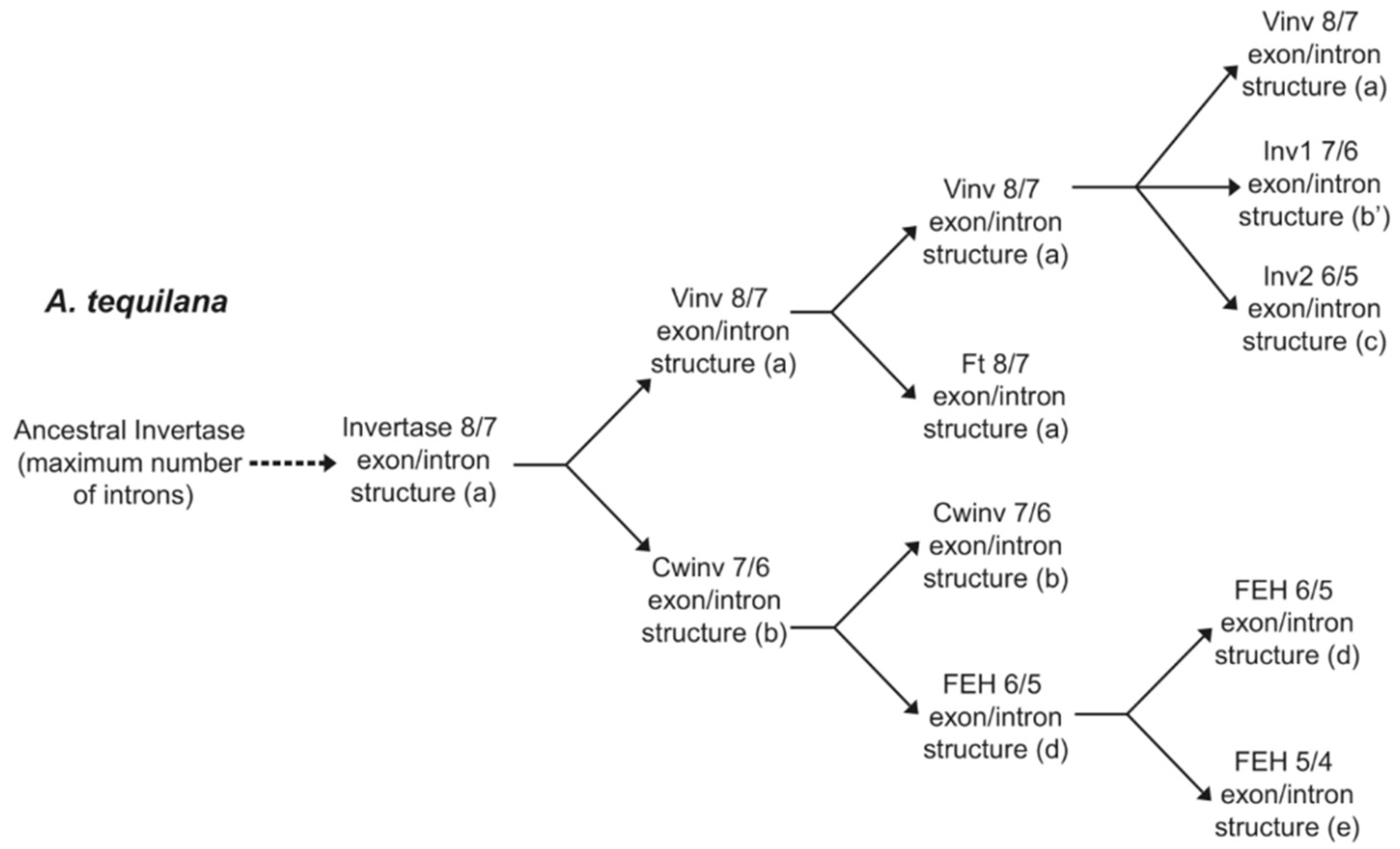
3.1. Determination of Gene Structures for A. tequilana PGHF32 Encoding Genes
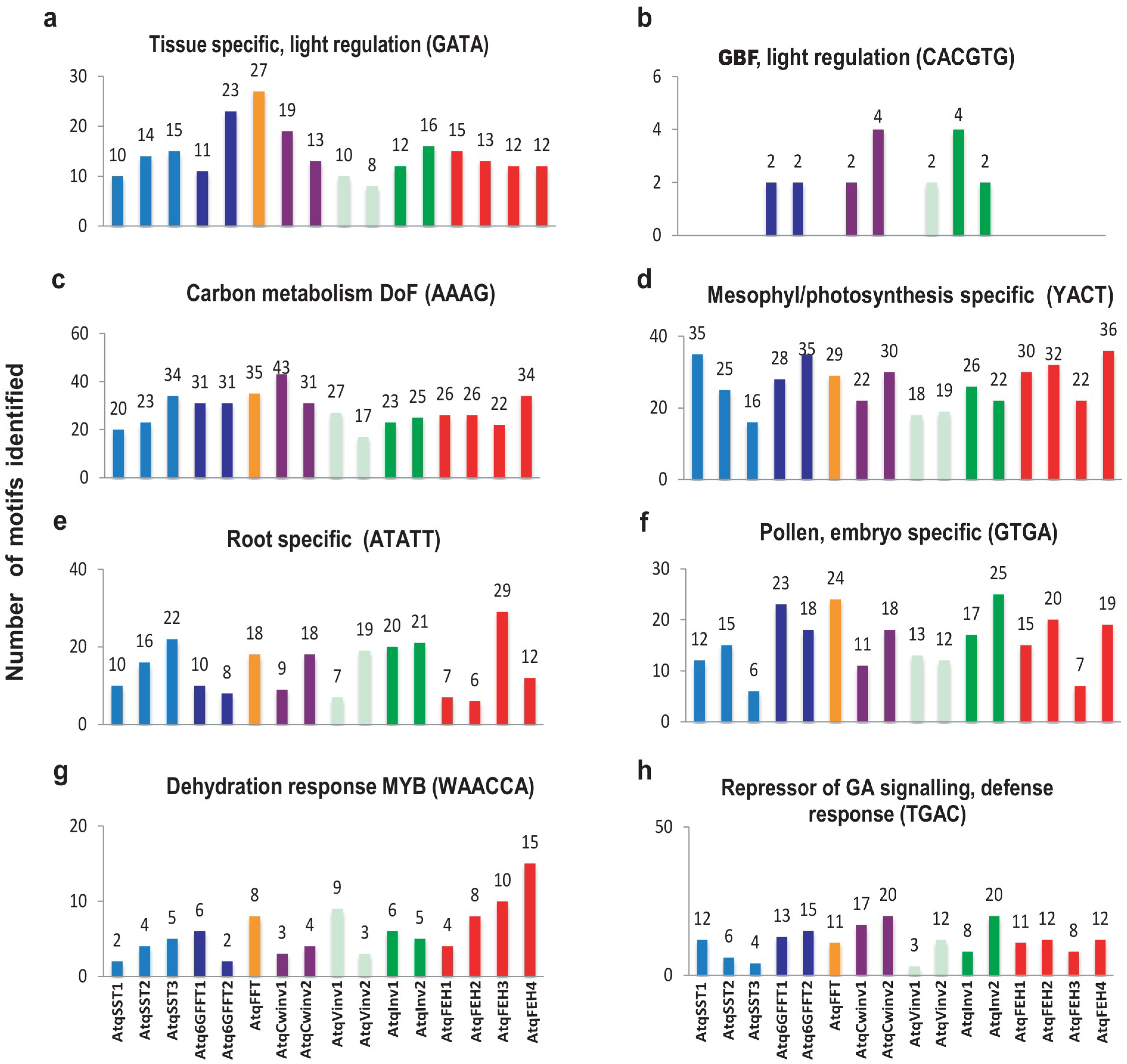
3.2. Identification of Putative Regulatory Motifs in the Promotor Regions of Genes Encoding PGHF32 Enzymes in A. tequilana
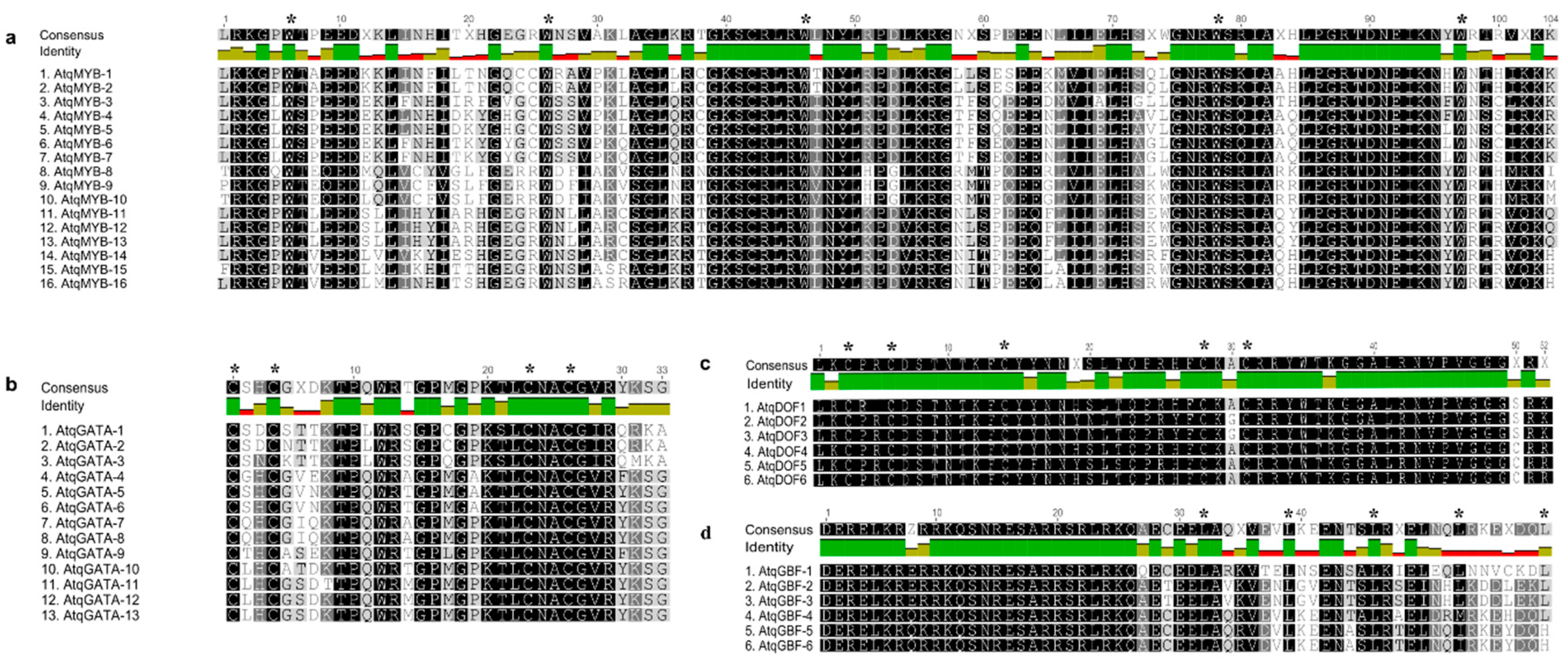
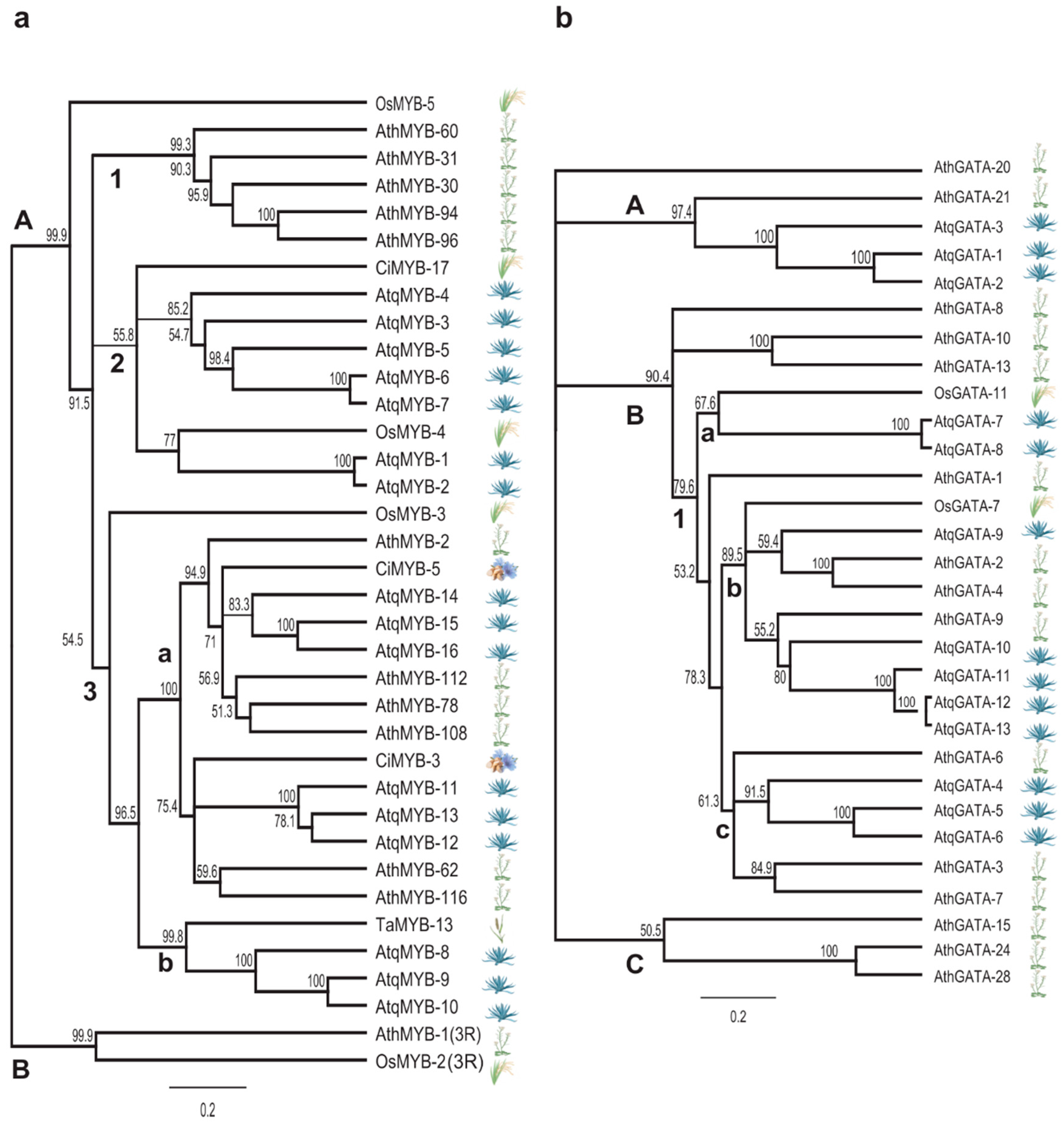
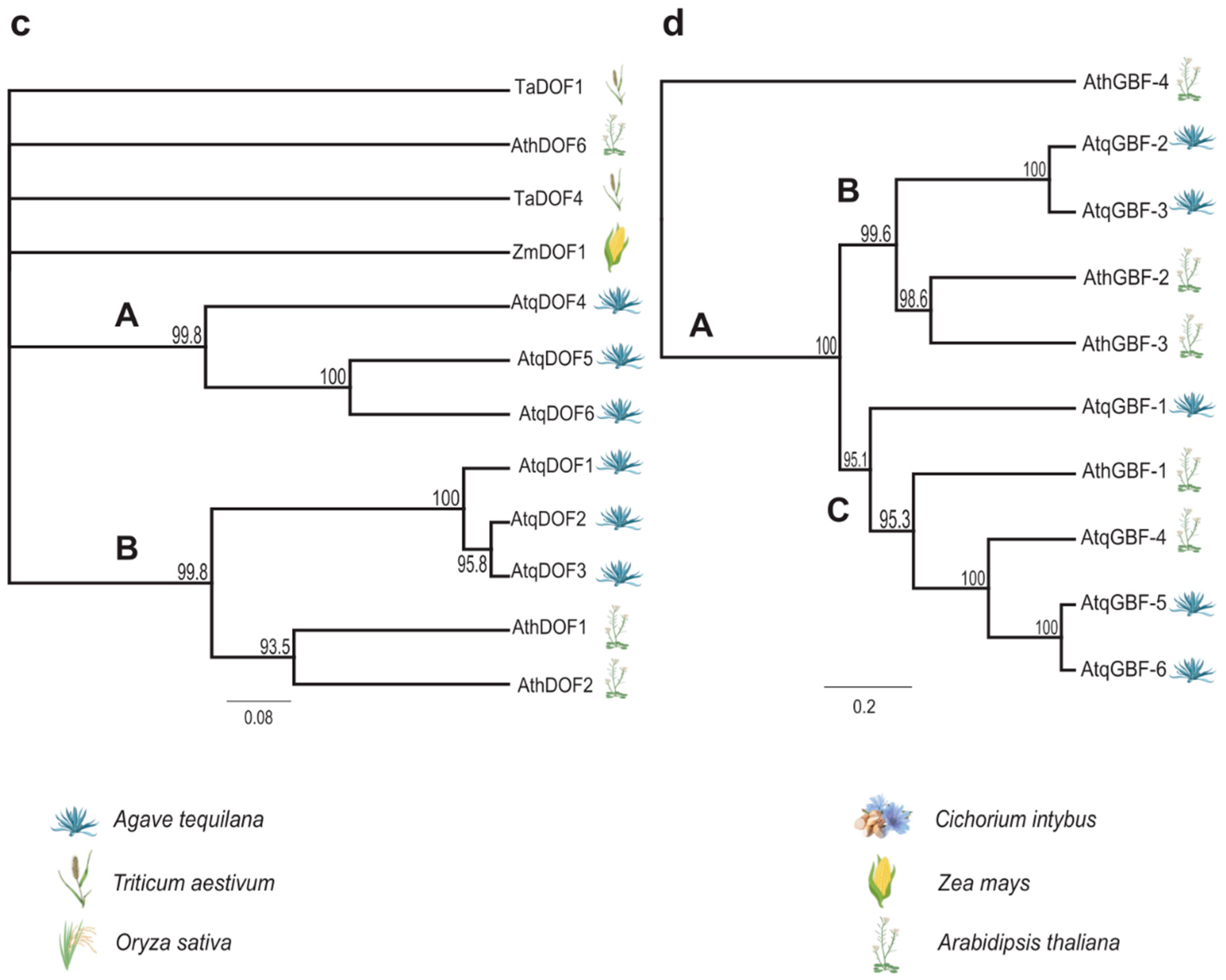
3.3. Identification and Analysis of Coding Sequences and Predicted Proteins for Transcription Factors Putatively Involved in the Regulation of PGHF32 Genes in A. tequilana
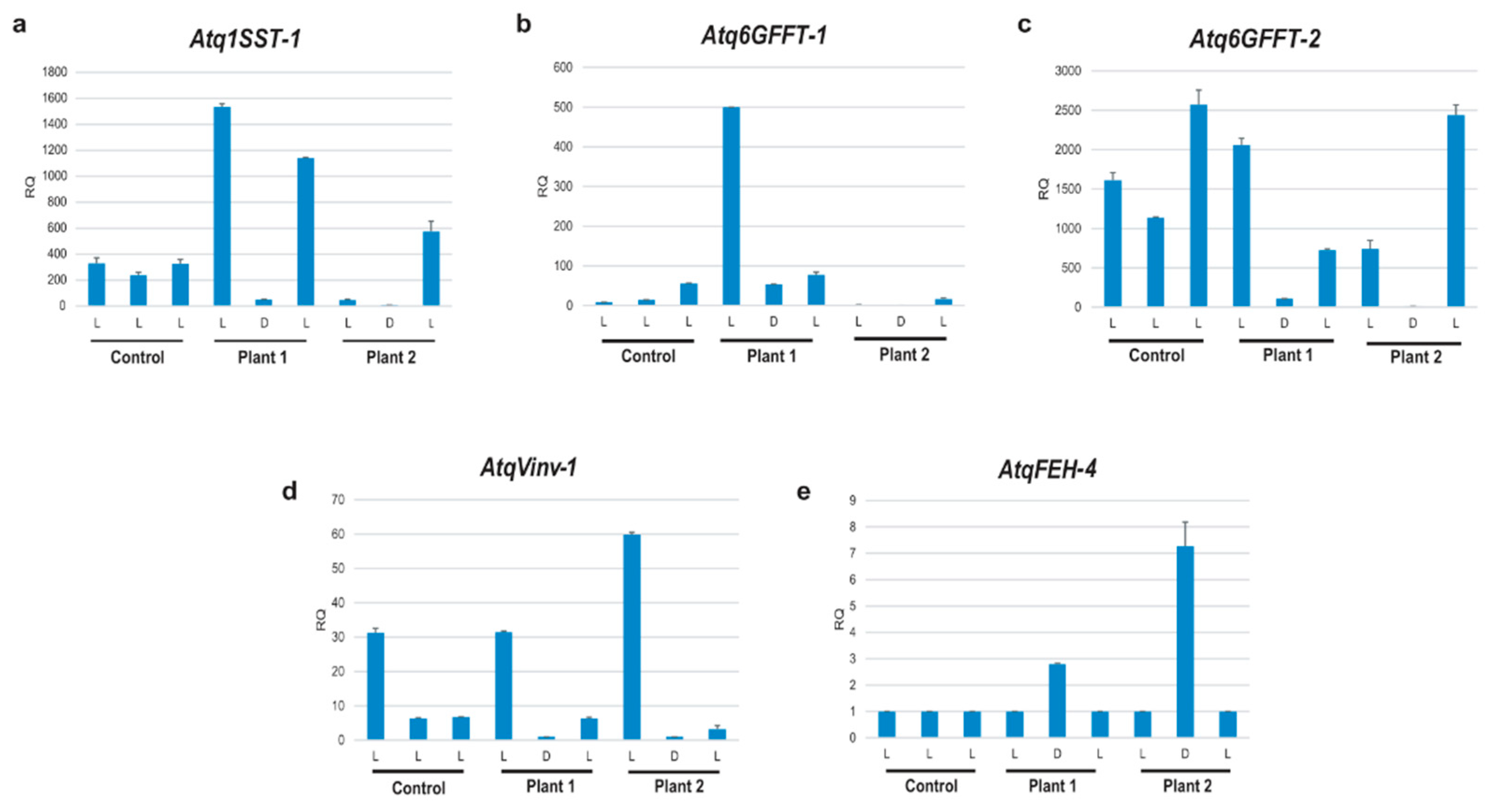
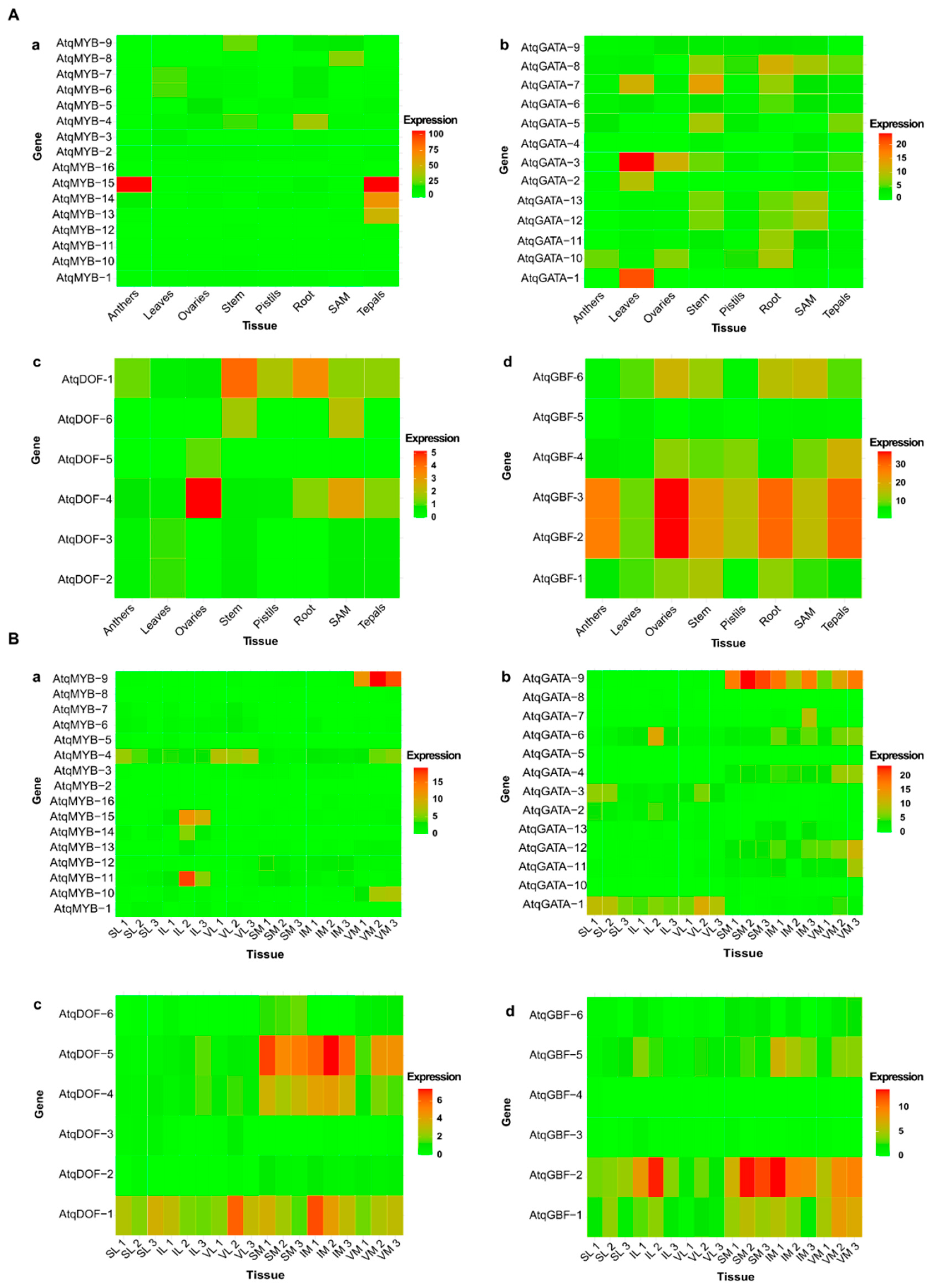
3.4. In Silico Expression Patterns of MYB, GATA GBF and DOF Transcription Factors Identified for A. tequilana
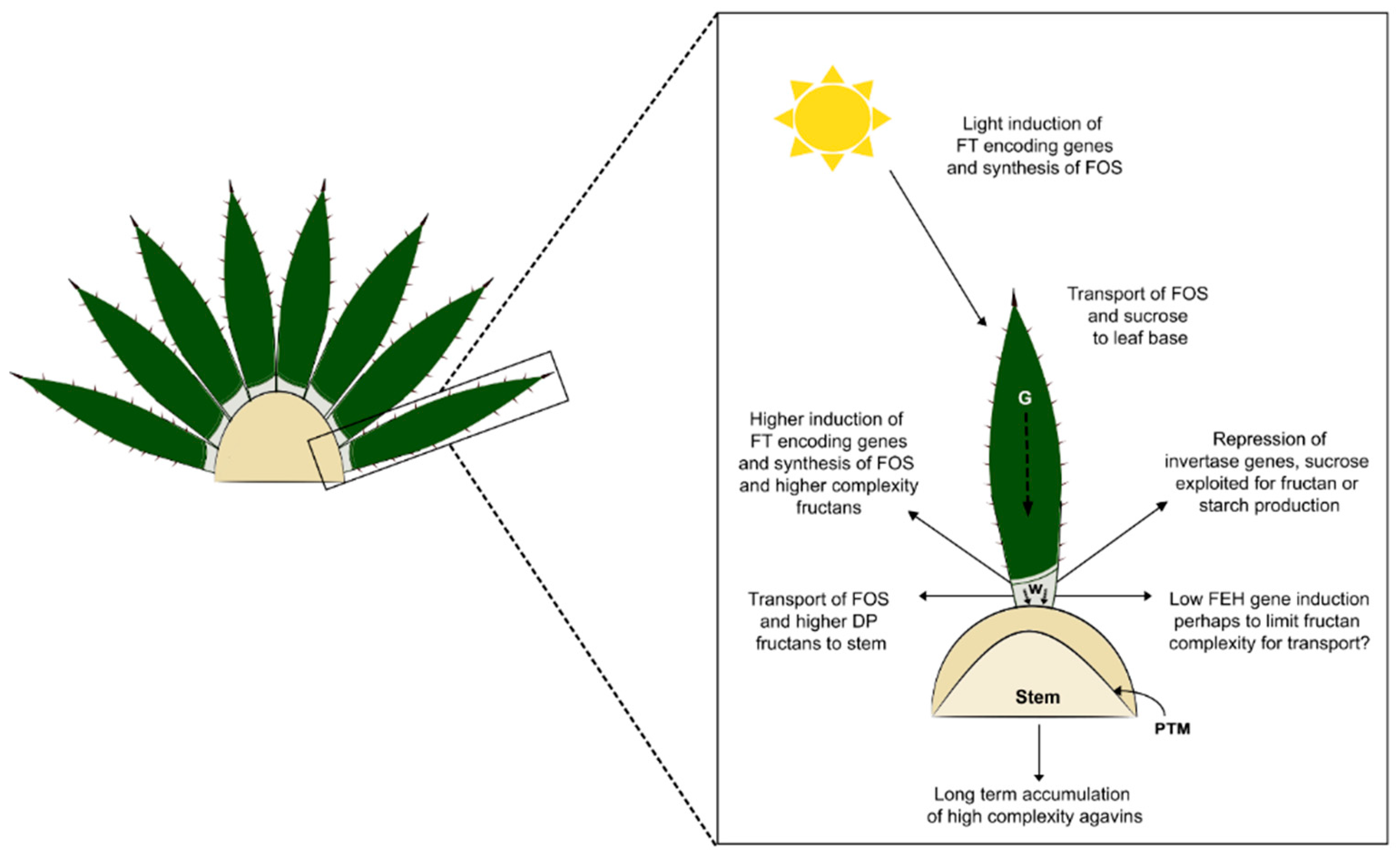
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hendry, G. Evolutionary origins and natural functions of fructans—A climatological, biogeographic and mechanistic appraisal. New Phytol. 1993, 123, 3–14. [Google Scholar] [CrossRef]
- Banguela, A.; Hernández, L. Fructans: From natural sources to transgenic plants. Biotecnol. Apl. 2006, 23, 202–210. [Google Scholar]
- Versluys, M.; Kirtel, O.; Toksoy Öner, E.; Van den Ende, W. The fructan syndrome: Evolutionary aspects and common themes among plants and microbes. Plant Cell Environ. 2017, 41, 16–38. [Google Scholar] [CrossRef]
- Pilon-Smits, E.; Ebskamp, M.; Paul, M.J.; Jeuken, M.; Weisbeek, P.J.; Smeekens, S. Improved Performance of Transgenic Fructan-Accumulating Tobacco under Drought Stress. Plant Physiol. 1995, 107, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Nobel, P.S. Phloem Transport of Fructans in the Crassulacean Acid Metabolism Species Agave deserti. Plant Physiol. 1998, 116, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Yoshida, M. Molecular characterization of sucrose:sucrose 1-fructosyltransferase and sucrose: Fructan 6-fructosyltransferase associated with fructan accumulation in winter wheat during cold hardening. Biosci. Biotechnol. Biochem. 2002, 66, 2297–2305. [Google Scholar] [CrossRef] [PubMed]
- Hisano, H.; Kanazawa, A.; Yoshida, M.; Humphreys, M.O.; Iizuka, M.; Kitamura, K.; Yamada, T. Coordinated expression of functionally diverse fructosyltransferase genes is associated with fructan accumulation in response to low temperature in perennial ryegrass. New Phytol. 2008, 178, 766–780. [Google Scholar] [CrossRef]
- Bieleski, R.L. Fructan hydrolysis drives petal expansion in the ephemeral daylily flower. Plant Physiol. 1993, 103, 213–219. [Google Scholar] [CrossRef]
- Vergauwen, R.; Van den Ende, W.; Van Laere, A. The role of fructan in flowering of Campanula rapunculoides. J. Exp. Bot. 2000, 51, 1261–1266. [Google Scholar] [CrossRef]
- Valluru, R. Fructan and hormone connections. Front. Plant Sci. 2015, 6, 180. [Google Scholar] [CrossRef]
- Versluys, M.; Tarkowski, Ł.P.; Van den Ende, W. Fructans as DAMPs or MAMPs: Evolutionary prospects, cross-tolerance, and multistress resistance potential. Front. Plant Sci. 2017, 7, 2061. [Google Scholar] [CrossRef] [PubMed]
- Narváez-Zapata, J.A.; Sánchez-Teyer, L.F. Agaves as a raw material: Recent technologies and applications. Recent Pat. Biotechnol. 2009, 3, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Cushman, J.C.; Davis, S.C.; Yang, X.; Borland, A.M. Development and use of bioenergy feedstocks for semi-arid and arid lands. J. Exp. Bot. 2015, 66, 4177–4193. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.C.; Simpson, J.; Gil Vega, K.C.; Niechayev, N.A.; van Tongerlo, E.; Hurtado Castano, N.; Dever, L.V.; Búrquez, A. Undervalued potential of crassulacean acid metabolism for current and future agricultural production. J. Exp. Bot. 2019, 70, 6521–6537. [Google Scholar] [CrossRef]
- Mancilla-Margalli, N.A.; Lopez, M.G. Water-soluble carbohydrates and fructan structure patterns from Agave and Dasylirion species. J. Agric. Food Chem. 2006, 54, 7832–7839. [Google Scholar] [CrossRef]
- Mellado-Mojica, E.; López, M.G. Fructan metabolism in A. tequilana Weber Blue variety along its developmental cycle in the field. J. Agric. Food Chem. 2012, 60, 11704–11713. [Google Scholar] [CrossRef]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 247. [Google Scholar] [CrossRef]
- Wei, J.Z.; Chatterton, N.J. Fructan biosynthesis and fructosyltransferase evolution: Expression of the 6-SFT (sucrose: Fructan 6-fructosyltransferase) gene in crested wheatgrass (Agropyron cristatum). J. Plant Physiol. 2001, 158, 1203–1213. [Google Scholar] [CrossRef]
- Van Den Ende, W.; De Coninck, B.; Clerens, S.; Vergauwen, R.; Van Laere, A. Unexpected presence of fructan 6-exohydrolases (6-FEHs) in non-fructan plants: Characterization, cloning, mass mapping and functional analysis of a novel “cell-wall invertase-like” specific 6-FEH from sugar beet (Beta vulgaris L.). Plant J. 2003, 36, 697–710. [Google Scholar] [CrossRef]
- Avila de Dios, E.; Gomez Vargas, A.D.; Damián Santos, M.L.; Simpson, J. New insights into plant glycoside hydrolase family 32 in Agave species. Front. Plant Sci. 2015, 6, 594. [Google Scholar] [CrossRef]
- Avila de Dios, E.; Delaye, L.; Simpson, J. Transcriptome analysis of bolting in A. tequilana reveals roles for florigen, MADS, fructans and gibberellins. BMC Genom. 2019, 20, 473. [Google Scholar] [CrossRef] [PubMed]
- Pérez López, A.V.; Simpson, J.; Clench, M.R.; Gomez Vargas, A.D.; Ordaz Ortiz, J.J. Localization and Composition of Fructans in Stem and Rhizome of Agave tequilana Weber var. azul. Front. Plant Sci. 2021, 11, 608850. [Google Scholar] [CrossRef] [PubMed]
- Suárez-González, E.M.; López, M.G.; Délano-Frier, J.P.; Gómez-Leyva, J.F. Expression of the 1-SST and 1-FFT genes and consequent fructan accumulation in Agave tequilana and A. inaequidens is differentially induced by diverse abiotic stress related elicitors. J. Plant Physiol. 2014, 171, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Suárez-González, E.M.; Palmeros Suárez, P.A.; Cruz-Rubio, J.M.; Martínez-Gallardo, N.A.; Cisneros Hernández, I.; Délano-Frier, J.P.; Gómez-Leyva, J.F. Differential fructan accumulation and expression of fructan biosynthesis, invertase and defense genes is induced in Agave tequilana plantlets by sucrose or stress-related elicitors. Agri Gene 2016, 2, 17–28. [Google Scholar] [CrossRef]
- Kooiker, M.; Drenth, J.; Glassop, D.; McIntyre, C.L.; Xue, G.P. TaMYB13-1, a R2R3 MYB transcription factor, regulates the fructan synthetic pathway and contributes to enhanced fructan accumulation in bread wheat. J. Exp. Bot. 2013, 64, 3681–3696. [Google Scholar] [CrossRef]
- Wei, H.; Zhao, H.; Su, T.; Bausewein, A.; Greiner, S.; Harms, K.; Rausch, T. Chicory R2R3-MYB transcription factors CiMYB5 and CiMYB3 regulate fructan 1-exohydrolase expression in response to abiotic stress and hormonal cues. J. Exp. Bot. 2017, 68, 4323–4338. [Google Scholar] [CrossRef]
- Shaw, L.M.; McIntyre, C.L.; Gresshoff, P.M.; Xue, G.P. Members of the Dof transcription factor family in Triticum aestivum are associated with light-mediated gene regulation. Funct. Integr. Genom. 2009, 9, 485–498. [Google Scholar] [CrossRef]
- Xue, G.P.; Kooiker, M.; Drenth, J.; McIntyre, C.L. TaMYB13 is a transcriptional activator of fructosyltransferase genes involved in β-2,6-linked fructan synthesis in wheat. Plant J. 2011, 68, 857–870. [Google Scholar] [CrossRef]
- Chiang, Y.H.; Zubo, Y.O.; Tapken, W.; Kim, H.J.; Lavanway, A.M.; Howard, L.; Pilon, M.; Kieber, J.J.; Schaller, G.E. Functional characterization of the GATA transcription factors GNC and CGA1 reveals their key role in chloroplast development, growth, and division in Arabidopsis. Plant Physiol. 2012, 160, 332–348. [Google Scholar] [CrossRef]
- Zhao, H.; Greiner, S.; Scheffzek, K.; Rausch, T.; Wang, G.A. 6&1-FEH encodes an enzyme for fructan degradation and interact with invertase inhibitor protein in maize (Zea mays L.). Int. J. Mol. Sci. 2019, 20, 3807. [Google Scholar] [CrossRef]
- Wei, H.; Bausewein, A.; Greiner, S.; Dauchot, N.; Harms, K.; Rausch, T. CiMYB17, a stress-induced chicory R2R3-MYB transcription factor, activates promoters of genes involved in fructan synthesis and degradation. New Phytol. 2017, 215, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Escamilla-Treviño, L. Potential of Plants from the Genus Agave as Bioenergy Crops. Bioenergy Res. 2011, 5, 1–9. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gertz, E.M.; Agarwala, R.; Schäffer, A.A.; Yu, Y.K. PSI-BLAST pseudocounts and the minimum description length principle. Nucleic Acids Res. 2009, 37, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Cortés-Romero, C.; Martínez-Hernández, A.; Mellado-Mojica, E.; López, M.G.; Simpson, J. Molecular and Functional Characterization of Novel Fructosyltransferases and Invertases from Agave tequilana. PLoS ONE 2012, 7, e35878. [Google Scholar] [CrossRef]
- Sibéril, Y.; Doireau, P.; Gantet, P. Plant bZIP G-box binding factors. Eur. J. Biochem. 2001, 268, 5655–5666. [Google Scholar] [CrossRef]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Ezer, D.; Shepherd, S.J.K.; Brestovitsky, A.; Dickinson, P.; Cortijo, S.; Charoensawan, V.; Box, M.S.; Biswas, S.; Jaeger, K.E.; Wigge, P.A. The G-box transcriptional regulatory code in arabidopsis. Plant Physiol. 2017, 175, 628–640. [Google Scholar] [CrossRef]
- Wang, W.; Sijacic, P.; Xu, P.; Lian, H.; Liu, Z. Arabidopsis TSO1 and MYB3R1 form a regulatory module to coordinate cell proliferation with differentiation in shoot and root. Proc. Natl. Acad. Sci. USA 2018, 115, E3045–E3054. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. Abiotic stresses cause differential regulation of alternative splice forms of GATA transcription factor in rice. Front. Front. Plant Sci. 2017, 8, 1944. [Google Scholar] [CrossRef] [PubMed]
- Manfield, I.W.; Devlin, P.F.; Jen, C.H.; Westhead, D.R.; Gilmartin, P.M. Conservation, convergence, and divergence of light-responsive, circadian-regulated, and tissue-specific expression patterns during evolution of the Arabidopsis GATA gene family. Plant Physiol. 2007, 143, 941–958. [Google Scholar] [CrossRef]
- Soitamo, A.J.; Piippo, M.; Allahverdiyeva, Y.; Battchikova, N.; Aro, E.M. Light has a specific role in modulating Arabidopsis gene expression at low temperature. BMC Plant Biol. 2008, 8, 13. [Google Scholar] [CrossRef]
- Tsujimoto-Inui, Y.; Naito, Y.; Sakurai, N.; Suzuki, H.; Sasaki, R.; Takahashi, H.; Ohtsuki, N.; Nakano, T.; Yanagisawa, S.; Shibata, D.; et al. Functional genomics of the Dof transcription factor family genes in suspension-cultured cells of Arabidopsis thaliana. Plant Biotechnol. 2009, 26, 15–28. [Google Scholar] [CrossRef]
- Noguero, M.; Atif, R.M.; Ochatt, S.; Thompson, R.D. The role of the DNA-binding One Zinc Finger (DOF) transcription factor family in plants. Plant Sci. 2013, 209, 32–45. [Google Scholar] [CrossRef]
- Ramegowda, V.; Gill, U.S.; Sivalingam, P.N.; Gupta, A.; Gupta, C.; Govind, G.; Nataraja, K.N.; Pereira, A.; Udayakumar, M.; Mysore, K.S.; et al. GBF3 transcription factor imparts drought tolerance in Arabidopsis thaliana. Sci. Rep. 2017, 7, 9148. [Google Scholar] [CrossRef]
- Ritsema, T.; Smeekens, S.C.M. Engineering fructan metabolism in plants. J. Plant Physiol. 2003, 160, 811–820. [Google Scholar] [CrossRef]
- Ueno, K.; Onodera, S.; Kawakami, A.; Yoshida, M.; Shiomi, N. Molecular characterization and expression of a cDNA encoding fructan:fructan 6G-fructosyltransferase from asparagus (Asparagus officinalis). New Phytol. 2005, 165, 813–824. [Google Scholar] [CrossRef]
- Le Roy, K.; Vergauwen, R.; Struyf, T.; Yuan, S.; Lammens, W.; Mátrai, J.; De Maeyer, M.; Van den Ende, W. Understanding the role of defective invertases in plants: Tobacco Nin88 fails to degrade sucrose. Plant Physiol. 2013, 161, 1670–1681. [Google Scholar] [CrossRef]
- Simpson, C.G.; Hedley, P.E.; Watters, J.A.; Clark, G.P.; McQuade, C.; Machray, G.C.; Brown, W.S. Requirements for mini-exon inclusion in potato invertase mRNAs provide evidence for exon-scanning interactions in plants. RNA 2000, 6, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Zavala-García, L.E.; Sánchez-Segura, L.; Avila de Dios, E.; Pérez-López, A.; Simpson, J. Starch accumulation is associated with active growth in A. tequilana. Plant Physiol. Biochem. 2018, 130, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Lau, O.; Deng, X. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 2007, 8, 217–2300. [Google Scholar] [CrossRef]
- Yanagisawa, S. Dof1 and Dof2 transcription factors are associated with expression of multiple genes involved in carbon metabolism in maize. Plant J. 2000, 21, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Gao, X.; Ma, K.; Li, D.; Jia, C.; Zhai, M.; Xu, Z. The walnut transcription factor JrGRAS2 contributes to high temperature stress tolerance involving in Dof transcriptional regulation and HSP protein expression. BMC Plant Biol. 2018, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Banu, S.; Huda, K.; Tuteja, N. Isolation and functional characterization of the promoter of a DEAD-box helicase Psp68 using Agrobacterium-mediated transient assay. Plant Signal. Behav. 2014, 9, e28992. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, Z.; Sebastian, A.; Xu, W.; Grain, D.; Salsac, F.; Avon, A.; Berger, N.; Tran, J.; Dubreucq, B.; Lurin, C.; et al. Analysis of the DNA-Binding Activities of the Arabidopsis R2R3-MYB Transcription Factor Family by One-Hybrid Experiments in Yeast. PLoS ONE 2015, 10, e0141044. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Vargas, A.D.; Hernández-Martínez, K.M.; López-Rosas, M.E.; Alejo Jacuinde, G.; Simpson, J. Evidence for Light and Tissue Specific Regulation of Genes Involved in Fructan Metabolism in Agave tequilana. Plants 2022, 11, 2153. https://doi.org/10.3390/plants11162153
Gomez-Vargas AD, Hernández-Martínez KM, López-Rosas ME, Alejo Jacuinde G, Simpson J. Evidence for Light and Tissue Specific Regulation of Genes Involved in Fructan Metabolism in Agave tequilana. Plants. 2022; 11(16):2153. https://doi.org/10.3390/plants11162153
Chicago/Turabian StyleGomez-Vargas, Alan D., Karen M. Hernández-Martínez, Macrina E. López-Rosas, Gerardo Alejo Jacuinde, and June Simpson. 2022. "Evidence for Light and Tissue Specific Regulation of Genes Involved in Fructan Metabolism in Agave tequilana" Plants 11, no. 16: 2153. https://doi.org/10.3390/plants11162153