
Genetics of the Inverse Relationship between Grain Yield and Grain Protein Content in Common Wheat


Abstract
:1. Introduction
2. Results
2.1. Phenotypic Analysis
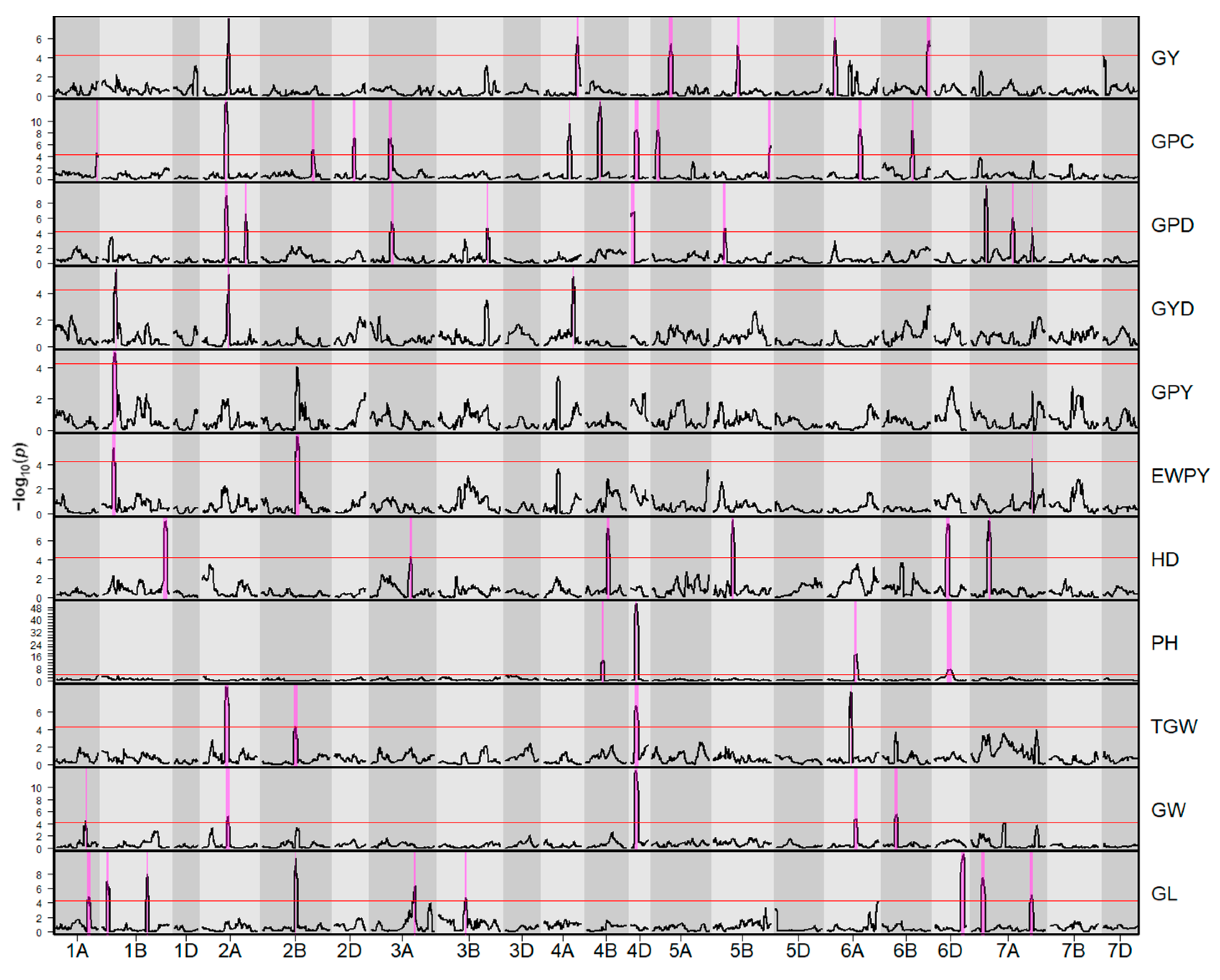
2.2. QTL Analysis
2.2.1. Summary
2.2.2. Grain Yield
2.2.3. Grain Protein Content
2.2.4. Indices Derived from Grain Yield and Grain Protein Content
2.2.5. Heading Date and Plant Height
2.2.6. Grain Morphometric Traits
2.2.7. Coinciding QTL
3. Discussion
3.1. Candidate Genes for GY, GPC, and Derived Indices
3.1.1. Rht-B1, Rht-D1, and Rht24
3.1.2. WAPO-A1
3.1.3. TaGS3-4A
3.1.4. TaGSe-4B
3.1.5. TaTEF-7A
3.1.6. TaNRT2.6-7A
3.1.7. Sucrose Synthase (TraesCS4A02G446700)
3.1.8. Limitations of Identified Candidate Genes
3.2. Colocalizing QTL from Previous Studies
3.3. Implications for Quality Wheat Breeding
4. Materials and Methods
4.1. Plant Material
4.2. Phenotyping and Phenotypic Data Analysis
4.3. Genotyping and Genotypic Data Analysis
4.4. QTL Analysis
4.5. Candidate Genes and Previously Identified QTL
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 4 July 2022).
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.S. Genetic and other factors affecting wheat quality. In Achieving Sustainable Cultivation of Wheat; Langridge, P., Ed.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2017; Volume 1, pp. 177–211. [Google Scholar]
- Saint Pierre, C.; Peterson, C.J.; Ross, A.S.; Ohm, J.B.; Verhoeven, M.C.; Larson, M.; Hoefer, B. White wheat grain quality changes with genotype, nitrogen fertilization, and water stress. Agron. J. 2008, 100, 414–420. [Google Scholar] [CrossRef]
- Maphosa, L.; Langridge, P.; Taylor, H.; Emebiri, L.C.; Mather, D.E. Genetic control of grain protein, dough rheology traits and loaf traits in a bread wheat population grown in three environments. J. Cereal Sci. 2015, 64, 147–152. [Google Scholar] [CrossRef]
- Oury, F.X.; Godin, C. Yield and grain protein concentration in bread wheat: How to use the negative relationship between the two characters to identify favourable genotypes? Euphytica 2007, 157, 45–57. [Google Scholar] [CrossRef]
- Laidig, F.; Piepho, H.P.; Rentel, D.; Drobek, T.; Meyer, U.; Huesken, A. Breeding progress, environmental variation and correlation of winter wheat yield and quality traits in German official variety trials and on-farm during 1983–2014. Theor. Appl. Genet. 2017, 130, 223–245. [Google Scholar] [CrossRef]
- Monaghan, J.; Snape, J.; Chojecki, A.; Kettlewell, P. The use of grain protein deviation for identifying wheat cultivars with high grain protein concentration and yield. Int. J. Plant Breed. 2001, 122, 309–317. [Google Scholar] [CrossRef]
- Bogard, M.; Allard, V.; Brancourt-Hulmel, M.; Heumez, E.; Machet, J.M.; Jeuffroy, M.H.; Gate, P.; Martre, P.; Le Gouis, J. Deviation from the grain protein concentration-grain yield negative relationship is highly correlated to post-anthesis N uptake in winter wheat. J. Exp. Bot. 2010, 61, 4303–4312. [Google Scholar] [CrossRef]
- Taulemesse, F.; Gouis, J.L.; Gouache, D.; Gibon, Y.; Allard, V. Bread wheat (Triticum aestivum L.) grain protein concentration is related to early post-flowering nitrate uptake under putative control of plant satiety level. PLoS ONE 2016, 11, e0149668. [Google Scholar] [CrossRef]
- Nehe, A.S.; Misra, S.; Murchie, E.H.; Chinnathambi, K.; Singh Tyagi, B.; Foulkes, M.J. Nitrogen partitioning and remobilization in relation to leaf senescence, grain yield and protein concentration in Indian wheat cultivars. Field Crop. Res. 2020, 251, 107778. [Google Scholar] [CrossRef]
- Rapp, M.; Lein, V.; Lacoudre, F.; Lafferty, J.; Müller, E.; Vida, G.; Bozhanova, V.; Ibraliu, A.; Thorwarth, P.; Piepho, H.P.; et al. Simultaneous improvement of grain yield and protein content in durum wheat by different phenotypic indices and genomic selection. Theor. Appl. Genet. 2018, 131, 1315–1329. [Google Scholar] [CrossRef] [PubMed]
- Nigro, D.; Gadaleta, A.; Mangini, G.; Colasuonno, P.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Simeone, R.; Blanco, A. Candidate genes and genome-wide association study of grain protein content and protein deviation in durum wheat. Planta 2019, 249, 1157–1175. [Google Scholar] [CrossRef] [PubMed]
- Thorwarth, P.; Liu, G.; Ebmeyer, E.; Schacht, J.; Schachschneider, R.; Kazman, E.; Reif, J.C.; Würschum, T.; Longin, C.F.H. Dissecting the genetics underlying the relationship between protein content and grain yield in a large hybrid wheat population. Theor. Appl. Genet. 2019, 132, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Neuweiler, J.E.; Maurer, H.P.; Würschum, T. Genetic architecture of phenotypic indices for simultaneous improvement of protein content and grain yield in triticale (×triticosecale). Plant Breed. 2021, 140, 232–245. [Google Scholar] [CrossRef]
- Bogard, M.; Jourdan, M.; Allard, V.; Martre, P.; Perretant, M.R.; Ravel, C.; Heumez, E.; Orford, S.; Snape, J.; Griffiths, S.; et al. Anthesis date mainly explained correlations between post-anthesis leaf senescence, grain yield, and grain protein concentration in a winter wheat population segregating for flowering time QTLs. J. Exp. Bot. 2011, 62, 3621–3636. [Google Scholar] [CrossRef]
- Bogard, M.; Allard, V.; Martre, P.; Heumez, E.; Snape, J.W.; Orford, S.; Griffiths, S.; Gaju, O.; Foulkes, J.; Le Gouis, J. Identifying wheat genomic regions for improving grain protein concentration independently of grain yield using multiple inter-related populations. Mol. Breed. 2013, 31, 587–599. [Google Scholar] [CrossRef]
- White, J.; Sharma, R.; Balding, D.; Cockram, J.; Mackay, I.J. Genome-wide association mapping of Hagberg falling number, protein content, test weight, and grain yield in U.K. wheat. Crop Sci. 2022, 62, 965–981. [Google Scholar] [CrossRef]
- Mosleth, E.F.; Wan, Y.; Lysenko, A.; Chope, G.A.; Penson, S.P.; Shewry, P.R.; Hawkesford, M.J. A novel approach to identify genes that determine grain protein deviation in cereals. Plant Biotechnol. J. 2015, 13, 625–635. [Google Scholar] [CrossRef]
- Stadlmeier, M.; Hartl, L.; Mohler, V. Usefulness of a multiparent advanced generation intercross population with a greatly reduced mating design for genetic studies in winter wheat. Front. Plant Sci. 2018, 9, 1825. [Google Scholar] [CrossRef]
- Corsi, B.; Obinu, L.; Zanella, C.M.; Cutrupi, S.; Day, R.; Geyer, M.; Lillemo, M.; Lin, M.; Mazza, L.; Percival-Alwyn, L.; et al. Identification of eight QTL controlling multiple yield components in a German multi-parental wheat population, including Rht24, WAPO-A1, WAPO-B1 and genetic loci on chromosomes 5A and 6A. Theor. Appl. Genet. 2021, 134, 1435–1454. [Google Scholar] [CrossRef]
- Pearce, S.; Saville, R.; Vaughan, S.P.; Chandler, P.M.; Wilhelm, E.P.; Sparks, C.A.; Al-Kaff, N.; Korolev, A.; Boulton, M.I.; Phillips, A.L.; et al. Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat. Plant Physiol. 2011, 157, 1820–1831. [Google Scholar] [CrossRef] [PubMed]
- Gale, M.D.; Youssefian, S. Dwarfing genes in wheat. In Progress in Plant Breeding; Russel, G.E., Ed.; Butterworths: London, UK, 1985; Volume 1, pp. 1–35. [Google Scholar]
- Kertesz, Z.; Flintham, J.E.; Gale, M.D. Effects of Rht dwarfing genes on wheat grain yield and its components under Eastern European conditions. Cereal Res. Commun. 1991, 19, 297–304. [Google Scholar]
- Youssefian, S.; Kirby, E.; Gale, M.D. Pleiotropic effects of the GA-insensitive Rht dwarfing genes in wheat. 2. Effects on leaf, stem, ear and floret growth. Field Crops Res. 1992, 28, 191–210. [Google Scholar] [CrossRef]
- Jobson, E.M.; Johnston, R.E.; Oiestad, A.J.; Martin, J.M.; Giroux, M.J. The impact of the wheat Rht-B1b semi-dwarfing allele on photosynthesis and seed development under field conditions. Front. Plant Sci. 2019, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wen, W.; Xie, L.; Fu, L.; Xu, D.; Fu, C.; Wang, D.; Chen, X.; Xia, X.; Chen, Q.; et al. Molecular mapping of reduced plant height gene Rht24 in bread wheat. Front. Plant Sci. 2017, 8, 1379. [Google Scholar] [CrossRef] [PubMed]
- Würschum, T.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. A modern Green Revolution gene for reduced height in wheat. Plant J. 2017, 92, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Xia, X.; Xu, D.; Liu, Y.; Xie, L.; Hassan, M.A.; Song, J.; Li, F.; Wang, D.; Zhang, Y.; et al. Rht24b, an ancient variation of TaGA2ox-A9, reduces plant height without yield penalty in wheat. New Phytol. 2022, 233, 738–750. [Google Scholar] [CrossRef]
- Ikeda, K.; Nagasawa, N.; Nagato, Y. ABERRANT PANICLE ORGANIZATION 1 temporally regulates meristem identity in rice. Dev. Biol. 2005, 282, 349–360. [Google Scholar] [CrossRef]
- Kuzay, S.; Xu, Y.; Zhang, J.; Katz, A.; Pearce, S.; Su, Z.; Fraser, M.; Anderson, J.A.; Brown-Guedira, G.; DeWitt, N.; et al. Identification of a candidate gene for a QTL for spikelet number per spike on wheat chromosome arm 7AL by high-resolution genetic mapping. Theor. Appl. Genet. 2019, 132, 2689–2705. [Google Scholar] [CrossRef]
- Muqaddasi, Q.H.; Brassac, J.; Koppolu, R.; Plieske, J.; Ganal, M.W.; Röder, M.S. TaAPO-A1, an ortholog of rice ABERRANT PANICLE ORGANIZATION 1, is associated with total spikelet number per spike in elite European hexaploid winter wheat (Triticum aestivum L.) varieties. Sci. Rep. 2019, 9, 13853. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Keeble-Gagnère, G.; Hickey, L.T.; Tibbits, J.; Hayden, M.; Pasam, R.K.; Friedt, W.; Snowdon, R.J.; Appels, R.; Wittkop, B. High-resolution mapping of rachis nodes per rachis, a critical determinant of grain yield components in wheat. Theor. Appl. Genet. 2019, 132, 2707–2719. [Google Scholar] [CrossRef]
- Kuzay, S.; Lin, H.; Li, C.; Chen, S.; Woods, D.P.; Zhang, J.; Lan, T.; von Korff, M.; Dubcovsky, J. WAPO-A1 is the causal gene of the 7AL QTL for spikelet number per spike in wheat. PLoS Genet. 2022, 18, e1009747. [Google Scholar] [CrossRef]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef]
- Zhang, W.; Li, H.F.; Zhi, L.Y.; Su, Q.N.; Liu, J.J.; Ren, X.L.; Meng, D.Y.; Zhang, N.; Ji, J.; Zhang, X.; et al. Functional markers developed from TaGS3, a negative regulator of grain weight and size, for marker-assisted selection in wheat. Crop J. 2020, 8, 943–952. [Google Scholar] [CrossRef]
- Bernard, S.M.; Møller, A.L.B.; Dionisio, G.; Kichey, T.; Jahn, T.P.; Dubois, F.; Baudo, M.; Lopes, M.S.; Tercé-Laforgue, T.; Foyer, C.H.; et al. Gene expression, cellular localisation and function of glutamine synthetase isozymes in wheat (Triticum aestivum L.). Plant Mol. Biol. 2008, 67, 89–105. [Google Scholar] [CrossRef]
- Li, X.; Zhao, X.; He, X.; Zhao, G.; Li, B.; Liu, D.; Zhang, A.; Zhang, X.; Tong, Y.; Li, Z. Haplotype analysis of the genes encoding glutamine synthetase plastic isoforms and their association with nitrogen-use- and yield-related traits in bread wheat. New Phytol. 2011, 189, 449–458. [Google Scholar] [CrossRef]
- Hu, M.; Zhao, X.; Liu, Q.; Hong, X.; Zhang, W.; Zhang, Y.; Sun, L.; Li, H.; Tong, Y. Transgenic expression of plastidic glutamine synthetase increases nitrogen uptake and yield in wheat. Plant Biotechnol. J. 2018, 16, 1858–1867. [Google Scholar] [CrossRef]
- Nigro, D.; Fortunato, S.; Giove, S.L.; Mazzucotelli, E.; Gadaleta, A. Functional Validation of Glutamine synthetase and Glutamate synthase Genes in Durum Wheat near Isogenic Lines with QTL for High GPC. Int. J. Mol. Sci. 2020, 21, 9253. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, J.; Zhao, Y.; Liu, Q.; Islam, S.; Yang, W.; Ma, W. Wheat glutamine synthetase TaGSr-4B is a candidate gene for a QTL of thousand grain weight on chromosome 4B. Theor. Appl. Genet. 2022, 135, 2369–2384. [Google Scholar] [CrossRef]
- Zheng, J.; Liu, H.; Wang, Y.; Wang, L.; Chang, X.; Jing, R.; Hao, C.; Zhang, X. TEF-7A, a transcript elongation factor gene, influences yield-related traits in bread wheat (Triticum aestivum L.). J. Exp. Bot. 2014, 65, 5351–5365. [Google Scholar] [CrossRef]
- Cristina, D.; Ciuca, M.; Mandea, V.; Cornea, C.P. Assessment of 25 genes reported to influence thousand grain weight in winter wheat germplasm. Cereal Res. Commun. 2022, 50, 237–243. [Google Scholar] [CrossRef]
- Teng, W.; He, X.; Tong, Y. Genetic Control of Efficient Nitrogen Use for High Yield and Grain Protein Concentration in Wheat: A Review. Plants 2022, 11, 492. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, P.; Liu, Q.; Li, G.; Di, D.; Xia, G.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W. TaANR1-TaBG1 and TaWabi5-TaNRT2s/NARs Link ABA Metabolism and Nitrate Acquisition in Wheat Roots. Plant Physiol. 2020, 182, 1440–1453. [Google Scholar] [CrossRef]
- Taulemesse, F.; Le Gouis, J.; Gouache, D.; Gibon, Y.; Allard, V. Post-flowering nitrate uptake in wheat is controlled by N status at flowering, with a putative major role of root nitrate transporter NRT2.1. PLoS ONE 2015, 10, e0120291. [Google Scholar] [CrossRef]
- Li, W.; He, X.; Chen, Y.; Jing, Y.; Shen, C.; Yang, J.; Teng, W.; Zhao, X.; Hu, W.; Hu, M.; et al. A wheat transcription factor positively sets seed vigour by regulating the grain nitrate signal. New Phytol. 2020, 225, 1667–1680. [Google Scholar] [CrossRef]
- Malik, P.; Kumar, J.; Sharma, S.; Sharma, R.; Sharma, S. Multi-locus genome-wide association mapping for spike-related traits in bread wheat (Triticum aestivum L.). BMC Genomics 2021, 22, 597. [Google Scholar] [CrossRef]
- Jiang, Q.; Hou, J.; Hao, C.; Wang, L.; Ge, H.; Dong, Y.; Zhang, X. The wheat (T. aestivum) sucrose synthase 2 gene (TaSus2) active in endosperm development is associated with yield traits. Funct. Integr. Genomics 2011, 11, 49–61. [Google Scholar] [CrossRef]
- Hou, J.; Jiang, Q.; Hao, C.; Wang, Y.; Zhang, H.; Zhang, X. Global selection on sucrose synthase haplotypes during a century of wheat breeding. Plant Physiol. 2014, 164, 1918–1929. [Google Scholar] [CrossRef]
- Mohler, V.; Albrecht, T.; Castell, A.; Diethelm, M.; Schweizer, G.; Hartl, L. Considering causal genes in the genetic dissection of kernel traits in common wheat. J. Appl. Genet. 2016, 57, 467–476. [Google Scholar] [CrossRef]
- Sehgal, D.; Mondal, S.; Guzman, C.; Garcia Barrios, G.; Franco, C.; Singh, R.; Dreisigacker, S. Validation of candidate gene-based markers and identification of novel loci for thousand-grain weight in spring bread wheat. Front. Plant Sci. 2019, 10, 1189. [Google Scholar] [CrossRef]
- Saini, D.K.; Srivastava, P.; Pal, N.; Gupta, P.K. Meta-QTLs, ortho-meta-QTLs and candidate genes for grain yield and associated traits in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2022, 135, 1049–1081. [Google Scholar] [CrossRef]
- Gudi, S.; Saini, D.K.; Singh, G.; Halladakeri, P.; Kumar, P.; Shamshad, M.; Tanin, M.J.; Singh, S.; Sharma, A. Unravelling consensus genomic regions associated with quality traits in wheat using meta-analysis of quantitative trait loci. Planta 2022, 255, 115. [Google Scholar] [CrossRef]
- Mosleth, E.F.; Lillehammer, M.; Pellny, T.K.; Wood, A.J.; Riche, A.B.; Hussain, A.; Griffiths, S.; Hawkesford, M.J.; Shewry, P.R. Genetic variation and heritability of grain protein deviation in European wheat genotypes. Field Crops. Res. 2020, 255, 107896. [Google Scholar] [CrossRef]
- Michel, S.; Löschenberger, F.; Ametz, C.; Pachler, B.; Sparry, E.; Bürstmayr, H. Simultaneous selection for grain yield and protein content in genomics-assisted wheat breeding. Theor. Appl. Genet. 2019, 132, 1745–1760. [Google Scholar] [CrossRef]
- Grubbs, F.E. Sample criteria for testing outlying observations. Ann. Math. Stat. 1950, 21, 27–58. [Google Scholar] [CrossRef]
- Komsta, L. Outliers: Tests for Outliers. R Package Version 0.14. 2011. Available online: https://CRAN.R-project.org/package=outliers (accessed on 9 June 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Plaschke, J.; Ganal, M.W.; Röder, M.S. Detection of genetic diversity in closely related bread wheat using microsatellite markers. Theor. Appl. Genet. 1995, 91, 1001–1007. [Google Scholar] [CrossRef]
- Wang, S.C.; Wong, D.B.; Forrest, K.; Allen, A.; Chao, S.M.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef]
- Winfield, M.O.; Allen, A.M.; Burridge, A.; Barker, G.L.A.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G.; et al. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- Huang, B.E.; George, A.W. R/mpMap: A computational platform for the genetic analysis of multiparent recombinant inbred lines. Bioinformatics 2011, 27, 727–729. [Google Scholar] [CrossRef]
- Durinck, S.; Spellman, P.T.; Birney, E.; Huber, W. Mapping identifiers for the integration of genomic datasets with the R/Bioconductor package biomaRt. Nat. Protoc. 2009, 4, 1184–1191. [Google Scholar] [CrossRef]


{kind=link}
| Trait | Env. | rep2 | h2 | σG2 | σE2 | σR2 | Range | Mean ± SD |
|---|---|---|---|---|---|---|---|---|
| Grain yield (GY; t/ha) | 9 | 0.71–0.92 | 0.83 | 0.17 | 0.83 | 0.31 | 6.2–9.3 | 8.1 ± 0.5 |
| Grain protein content (GPC; %) | 9 | 0.80–0.93 | 0.93 | 0.40 | 1.11 | 0.28 | 11.8–16.2 | 13.6 ± 0.7 |
| Grain protein deviation (GPD; %) | 8 | 0.67–0.90 | 0.88 | 0.21 | <0.01 | 0.22 | −1.6–1.6 | 0.0 ± 0.5 |
| Grain yield deviation (GYD; t/ha) | 8 | 0.55–0.86 | 0.76 | 0.08 | <0.01 | 0.19 | −1.4–0.7 | 0.0 ± 0.3 |
| Grain protein yield (GPY; t/ha) | 8 | 0.38–0.83 | 0.76 | <0.01 | 0.01 | <0.01 | 0.9–1.2 | 1.1 ± 0.0 |
| Equal weight protein yield (EWPY) | 8 | 0.29–0.83 | 0.79 | 0.19 | <0.01 | 0.41 | −1.9–1.3 | 0.0 ± 0.5 |
| Heading date (HD; d) | 7 | 0.84–0.93 | 0.96 | 2.38 | 18.62 | 0.73 | 25.6–35.1 | 30.1 ± 1.6 |
| Plant height (PH; cm) | 7 | 0.78–0.96 | 0.97 | 54.70 | 44.70 | 10.60 | 71.8–110.9 | 92.1 ± 7.5 |
| Thousand-grain weight (TGW; g) | 8 | 0.69–0.97 | 0.95 | 9.95 | 16.43 | 3.91 | 34.6–53.7 | 43.4 ± 3.2 |
| Grain width (GW; mm) | 7 | 0.70–0.95 | 0.94 | 0.01 | 0.02 | <0.01 | 3.1–3.8 | 3.5 ± 0.1 |
| Grain length (GL; mm) | 7 | 0.81–0.99 | 0.97 | 0.05 | 0.02 | 0.01 | 5.6–6.9 | 6.2 ± 0.2 |
| GY | GPC | GPD | GYD | GPY | EWPY | HD | PH | TGW | GW | |
|---|---|---|---|---|---|---|---|---|---|---|
| GPC | −0.60 | |||||||||
| GPD | −0.28 | 0.92 | ||||||||
| GYD | 0.86 | −0.20 | 0.18 | |||||||
| GPY | 0.58 | 0.24 | 0.58 | 0.90 | ||||||
| EWPY | 0.27 | 0.56 | 0.84 | 0.69 | 0.93 | |||||
| HD | −0.17 | ns | −0.10 | −0.18 | −0.20 | −0.19 | ||||
| PH | ns | 0.25 | 0.20 | ns | 0.14 | 0.17 | ns | |||
| TGW | 0.21 | 0.22 | 0.30 | 0.31 | 0.39 | 0.38 | −0.20 | 0.51 | ||
| GW | 0.13 | 0.28 | 0.33 | 0.26 | 0.37 | 0.38 | ns | 0.46 | 0.86 | |
| GL | 0.26 | ns | ns | 0.26 | 0.23 | 0.16 | −0.17 | 0.25 | 0.70 | 0.35 |
| Trait | Chr. | QTL | Position | Env. | −log10(p) | R2 | Candidate Gene |
|---|---|---|---|---|---|---|---|
| GY | 2A | QGy.lfl-2A | 139.6 (137.0–143.0) | 3 | 8.0 | 0.10 | - |
| 4A | QGy.lfl-4A | 183.6 (176.0–184.6) | 3 | 6.1 | 0.08 | TaGS3-4A | |
| 5A | QGy.lfl-5A | 99.0 (89.1–109.0) | 1 | 5.4 | 0.06 | - | |
| 5B | QGy.lfl-5B | 126.7 (124.0–137.0) | 0 | 5.2 | 0.06 | - | |
| 6A | QGy.lfl-6A | 44.5 (40.5–50.0) | 0 | 6.0 | 0.04 | - | |
| 6B | QGy.lfl-6B | 245.0 (235.1–249.8) | 2 | 5.7 | 0.04 | - | |
| GPC | 1A | QGpc.lfl-1A | 218.0 (212.3–220.2) | 1 | 4.6 | <0.01 | - |
| 2A | QGpc.lfl-2A | 127.3 (119.0–131.0) | 6 | 13.1 | 0.09 | - | |
| 2B | QGpc.lfl-2B | 271.0 (265.7–277.5) | 0 | 5.2 | 0.04 | - | |
| 2D | QGpc.lfl-2D | 108.6 (100.0–113.0) | 3 | 7.1 | 0.07 | - | |
| 3A | QGpc.lfl-3A | 111.0 (98.0–116.1) | 1 | 7.0 | 0.08 | - | |
| 4A | QGpc.lfl-4A | 139.0 (137.6–144.0) | 2 | 9.5 | 0.06 | - | |
| 4B | QGpc.lfl-4B | 75.0 (71.0–80.5) | 3 | 13.2 | 0.10 | Rht-B1; TaGSe-4B | |
| 4D | QGpc.lfl-4D | 29.0 (17.0–38.0) | 4 | 8.4 | 0.07 | Rht-D1 | |
| 5A | QGpc.lfl-5A | 29.0 (24.0–38.0) | 0 | 8.4 | 0.07 | - | |
| 5B | QGpc.lfl-5B | 302.5 (291.5–302.5) | 1 | 5.9 | 0.04 | - | |
| 6A | QGpc.lfl-6A | 179.2 (170.0–186.0) | 1 | 8.6 | 0.07 | - | |
| 6B | QGpc.lfl-6B | 156.1 (154.0–161.0) | 2 | 8.4 | 0.05 | - | |
| GPD | 2A | QGpd.lfl-2A.1 | 127.8 (122.0–131.9) | 2 | 9.1 | 0.08 | - |
| 2A | QGpd.lfl-2A.2 | 230.6 (227.0–234.7) | 0 | 6.6 | 0.06 | - | |
| 3A | QGpd.lfl-3A | 112.0 (109.9–125.0) | 1 | 5.6 | 0.08 | - | |
| 3B | QGpd.lfl-3B | 255.9 (254.4–266.0) | 1 | 4.6 | <0.01 | - | |
| 4D | QGpd.lfl-4D | 12.0 (0.0–17.0) | 1 | 6.9 | 0.04 | - | |
| 5B | QGpd.lfl-5B | 61.0 (52.0–63.7) | 0 | 4.7 | 0.03 | - | |
| 7A | QGpd.lfl-7A.1 | 80.4 (78.0–82.5) | 1 | 10.4 | 0.04 | TaTEF-7A | |
| 7A | QGpd.lfl-7A.2 | 219.5 (215.0–227.0) | 1 | 6.2 | 0.05 | NRT2.6-7A | |
| 7A | QGpd.lfl-7A.3 | 323.5 (322.0–326.0) | 2 | 4.8 | 0.04 | WAPO-A1 | |
| GYD | 1B | QGyd.lfl-1B | 73.3 (68.0–76.0) | 2 | 5.8 | 0.08 | - |
| 2A | QGyd.lfl-2A | 139.0 (134.0–143.0) | 0 | 5.4 | 0.06 | - | |
| 4A | QGyd.lfl-4A | 159.3 (154.0–161.8) | 1 | 5.3 | 0.08 | TraesCS4A02G446700 | |
| GPY | 1B | QGpy.lfl-1B | 67.2 (57.0–75.0) | 2 | 4.9 | 0.08 | - |
| EWPY | 1B | QEwpy.lfl-1B | 59.4 (52.8–67.7) | 1 | 5.4 | 0.06 | - |
| 2B | QEwpy.lfl-2B | 186.8 (182.3–199.0) | 1 | 6.3 | 0.07 | - | |
| 7A | QEwpy.lfl-7A | 323.5 (323.0–327.0) | 0 | 4.5 | 0.01 | WAPO-A1 | |
| HD | 1B | QHd.lfl-1B | 333.0 (324.0–345.0) | 3 | 8.1 | 0.04 | - |
| 3A | QHd.lfl-3A | 214.5 (209.4–223.0) | 1 | 4.3 | 0.05 | - | |
| 4B | QHd.lfl-4B | 116.7 (110.0–124.0) | 3 | 7.4 | 0.06 | - | |
| 5B | QHd.lfl-5B | 103.8 (96.2–105.8) | 3 | 8.2 | 0.10 | - | |
| 6D | QHd.lfl-6D | 70.0 (63.0–78.0) | 3 | 7.8 | 0.02 | - | |
| 7A | QHd.lfl-7A | 94.0 (91.2–102.0) | 1 | 8.2 | 0.09 | - | |
| PH | 4B | QPh.lfl-4B | 89.4 (83.0–95.7) | 6 | 13.3 | 0.11 | Rht-B1 |
| 4D | QPh.lfl-4D | 27.4 (23.0–31.0) | 7 | 50.4 | 0.27 | Rht-D1 | |
| 6A | QPh.lfl-6A | 159.4 (148.0–161.0) | 7 | 17.4 | 0.12 | Rht24 | |
| 6D | QPh.lfl-6D | 68.5 (67.5–94.0) | 6 | 7.5 | 0.03 | - | |
| TGW | 2A | QTgw.lfl-2A | 125.3 (121.8–136.5) | 5 | 8.8 | 0.05 | - |
| 2B | QTgw.lfl-2B | 178.8 (168.6–188.9) | 6 | 4.3 | 0.07 | - | |
| 4D | QTgw.lfl-4D | 25.0 (17.0–38.0) | 5 | 6.7 | 0.13 | Rht-D1 | |
| 6A | QTgw.lfl-6A | 130.0 (128.7–134.0) | 2 | 8.3 | 0.03 | - | |
| GW | 1A | QGw.lfl-1A | 155.8 (153.3–165.0) | 2 | 4.5 | 0.08 | - |
| 2A | QGw.lfl-2A | 136.0 (126.3–144.7) | 2 | 5.2 | 0.08 | - | |
| 4D | QGw.lfl-4D | 23.0 (17.0–34.0) | 6 | 12.8 | 0.20 | Rht-D1 | |
| 6A | QGw.lfl-6A | 156.0 (145.0–164.0) | 2 | 4.7 | 0.04 | Rht24 | |
| 6B | QGw.lfl-6B | 72.9 (60.0–80.0) | 3 | 5.5 | 0.06 | - | |
| GL | 1A | QGl.lfl-1A | 174.9 (164.0–181.0) | 3 | 4.9 | 0.03 | - |
| 1B | QGl.lfl-1B.1 | 23.0 (20.0–33.6) | 6 | 6.9 | 0.11 | - | |
| 1B | QGl.lfl-1B.2 | 238.9 (235.0–242.0) | 5 | 8.0 | 0.06 | - | |
| 2B | QGl.lfl-2B | 182.3 (178.8–183.8) | 4 | 10.3 | 0.04 | - | |
| 3A | QGl.lfl-3A | 239.0 (230.2–241.0) | 3 | 6.3 | 0.06 | - | |
| 3B | QGl.lfl-3B | 142.9 (139.0–150.0) | 4 | 4.7 | 0.04 | - | |
| 6D | QGl.lfl-6D | 153.0 (140.0–157.0) | 5 | 10.8 | 0.06 | - | |
| 7A | QGl.lfl-7A.1 | 60.0 (52.5–70.0) | 4 | 7.5 | 0.10 | - | |
| 7A | QGl.lfl-7A.2 | 324.0 (310.0–327.0) | 3 | 5.1 | <0.01 | WAPO-A1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geyer, M.; Mohler, V.; Hartl, L. Genetics of the Inverse Relationship between Grain Yield and Grain Protein Content in Common Wheat. Plants 2022, 11, 2146. https://doi.org/10.3390/plants11162146
Geyer M, Mohler V, Hartl L. Genetics of the Inverse Relationship between Grain Yield and Grain Protein Content in Common Wheat. Plants. 2022; 11(16):2146. https://doi.org/10.3390/plants11162146
Chicago/Turabian StyleGeyer, Manuel, Volker Mohler, and Lorenz Hartl. 2022. "Genetics of the Inverse Relationship between Grain Yield and Grain Protein Content in Common Wheat" Plants 11, no. 16: 2146. https://doi.org/10.3390/plants11162146