
Light-Dependence of Formate (C1) and Acetate (C2) Transport and Oxidation in Poplar Trees

 ,
,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
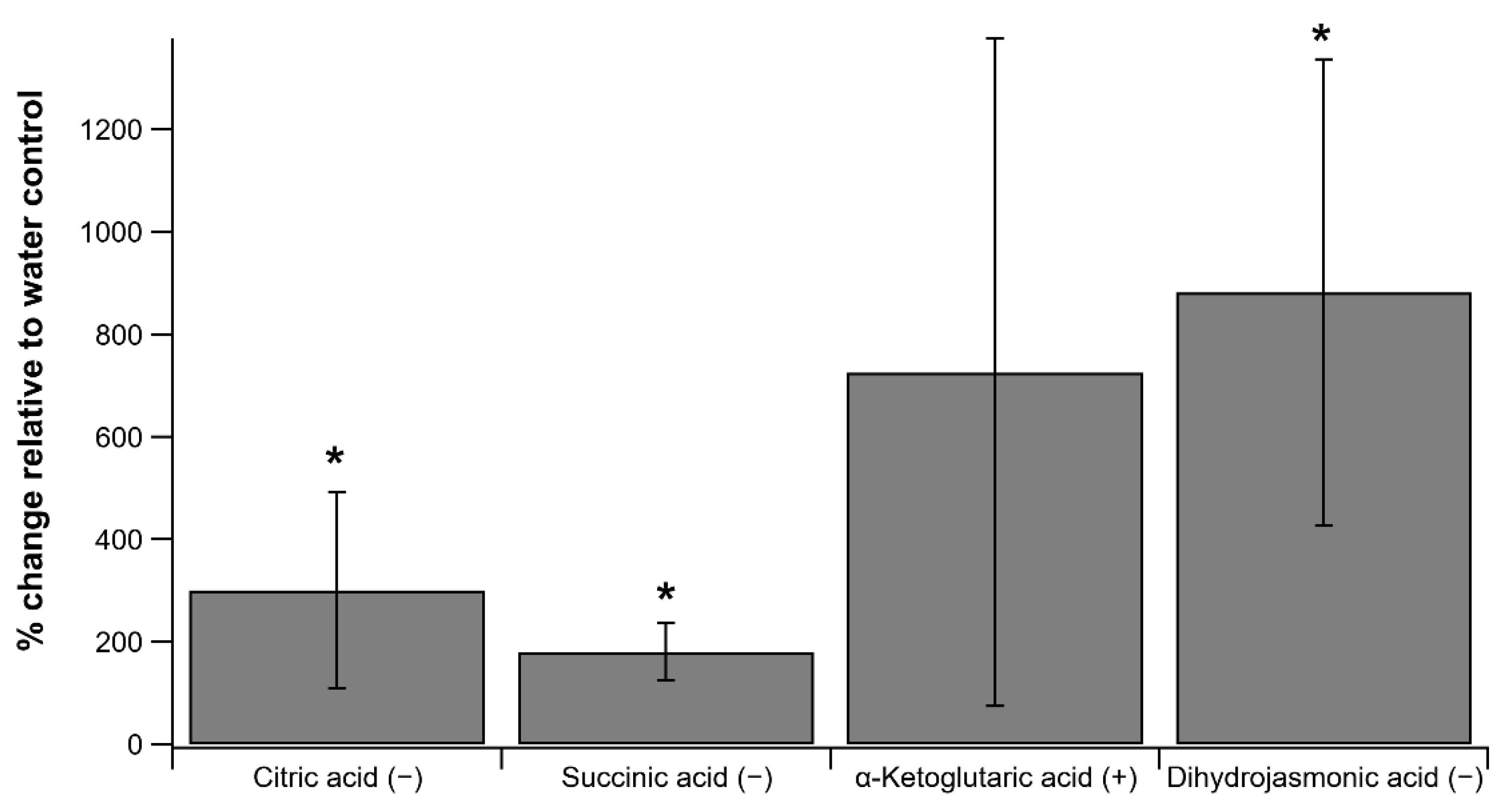
2.1. Leaf Concentrations of TCA Cycle Intermediates and Jasmonates
2.2. Dynamic Xylem Solution Injection (DXSI) System for Continuous Whole Tree Injections of [13C2]acetate and [13C]formate
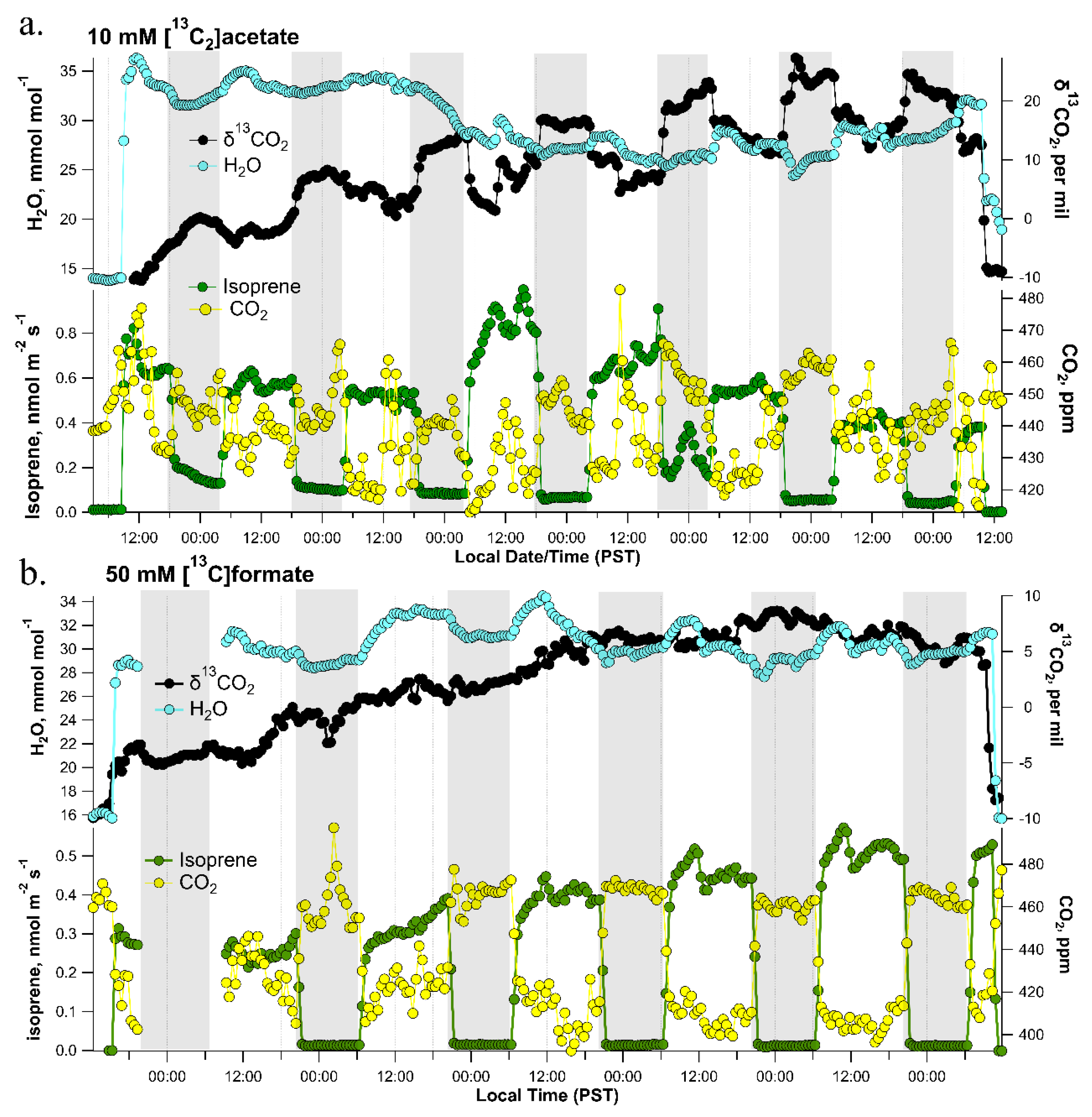
2.3. Light-Dependence of Headspace δ13CO2 of [13C2]acetate and [13C]formate Treated Detached Poplar Branches
3. Discussion
3.1. Dynamic Xylem Solution Injection (DXSI)
3.2. Transport and Metabolism of C1 and C2 Organic Acids in Detached Branches and Whole Trees
4. Materials and Methods
4.1. Plant Material
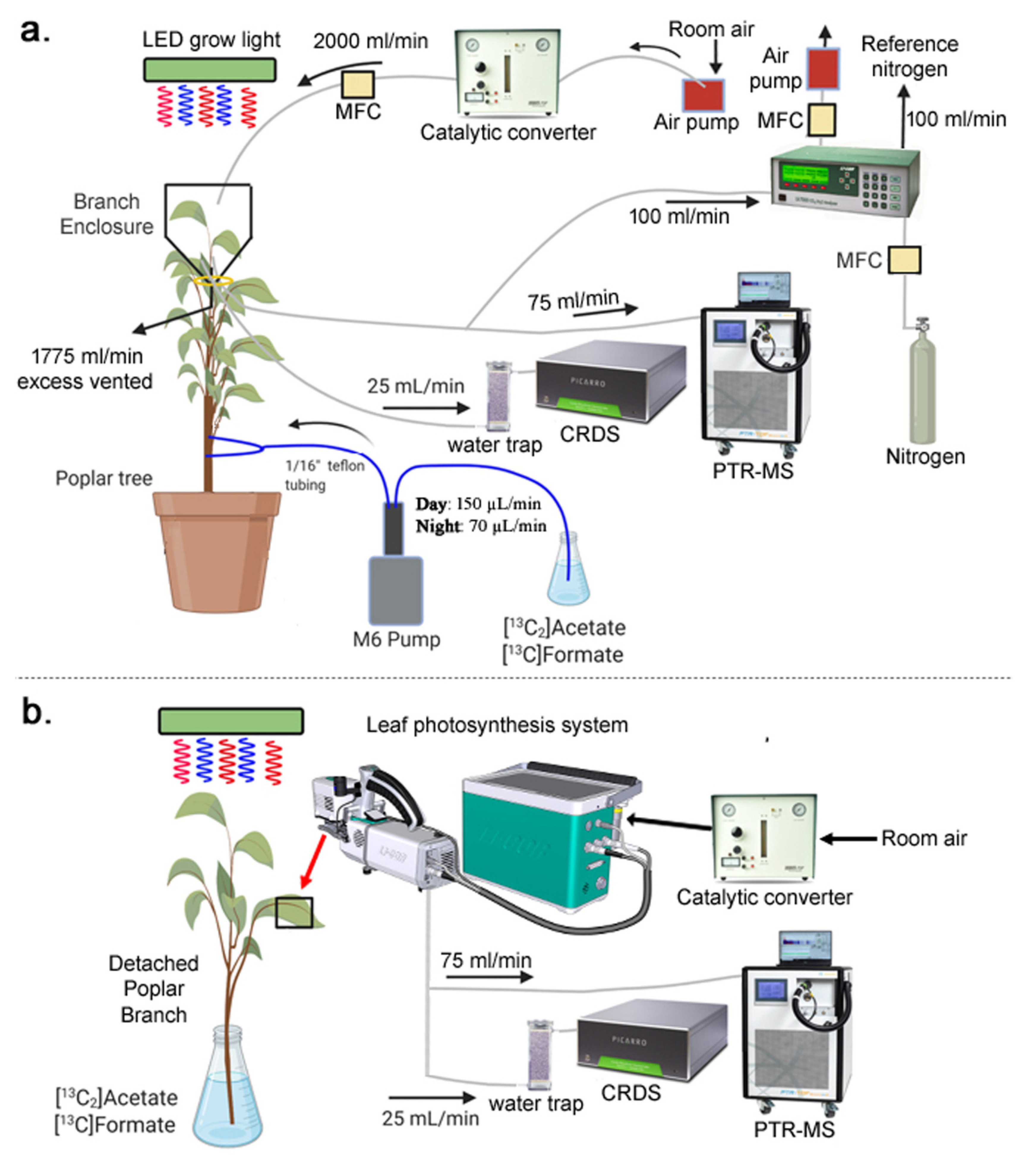
4.2. Dynamic Xylem Solution Injection (DXSI) System
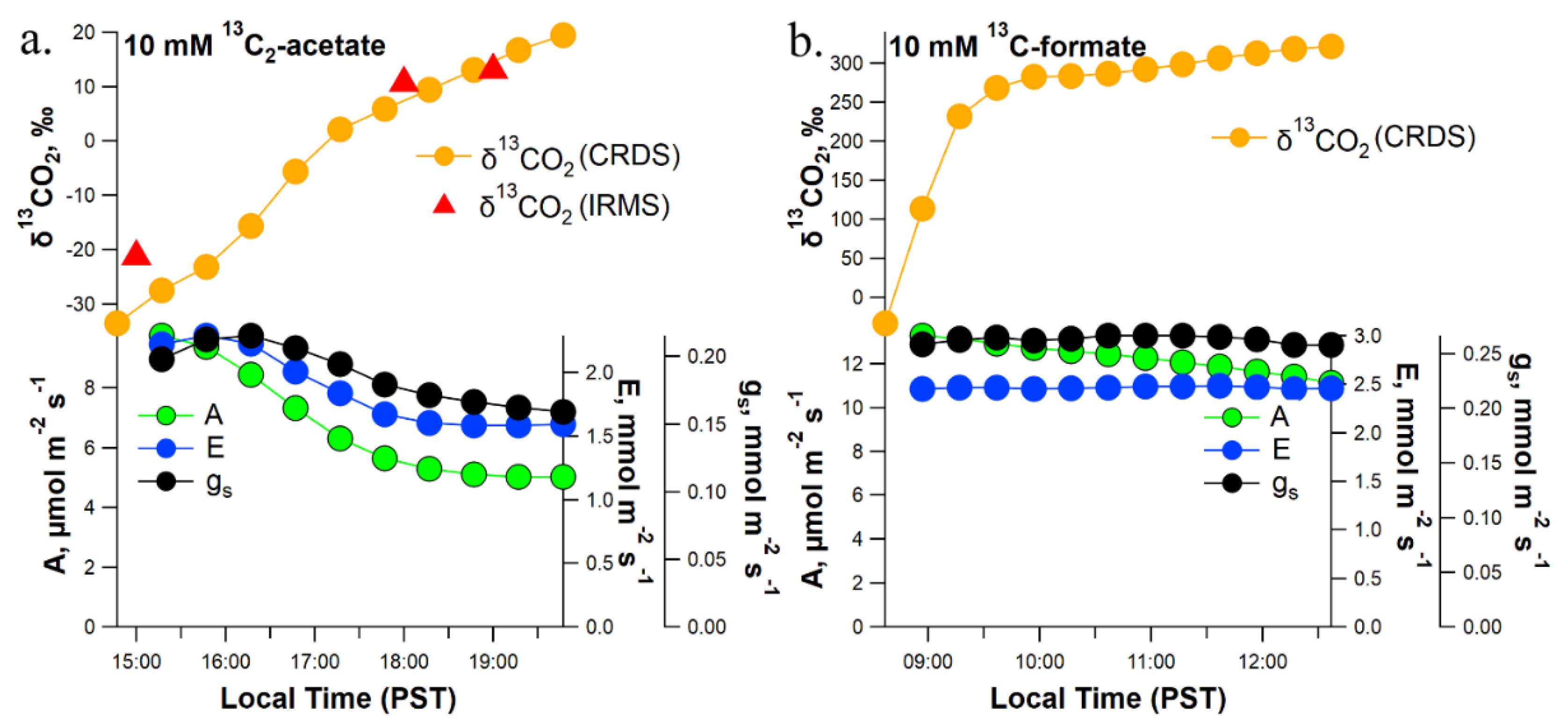
4.3. Detached Branch Labeling
4.4. Relative Quantification of Leaf TCA Cycle and Jasmonate in Acetate Treated Leaves
4.5. Branch Gas Exchange Analysis during Whole Plant DXSI Experiments
4.6. Leaf Gas Exchange Analysis under Constant Environmental Conditions from Detached Branches Pretreated with [13C2]acetate or [13C]formate Solutions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Leary, B.M.; Asao, S.; Millar, A.H.; Atkin, O.K. Core principles which explain variation in respiration across biological scales. New Phytol. 2019, 222, 670–686. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, G.C.; Dahal, K.; Alber, N.A.; Chadee, A. Photosynthesis, respiration and growth: A carbon and energy balancing act for alternative oxidase. Mitochondrion 2020, 52, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Méndez, A.; Miernyk, J.A.; Randall, D.D. Regulation of pyruvate dehydrogenase complex activity in plant cells. Eur. J. Biochem. 2003, 270, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Florez-Sarasa, I.; Fernie, A.R.; Gupta, K.J. Does the alternative respiratory pathway offer protection against the adverse effects resulting from climate change? J. Exp. Bot. 2020, 71, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Jardine, K.J.; Fernandes de Souza, V.; Oikawa, P.; Higuchi, N.; Bill, M.; Porras, R.; Niinemets, U.; Chambers, J.Q. Integration of C₁ and C₂ metabolism in trees. Int. J. Mol. Sci. 2017, 18, 2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, P.L.; Ramberg, H.; Baack, R.D.; Markwell, J.; Osterman, J.C. Formate dehydrogenase in Arabidopsis thaliana: Overexpression and subcellular localization in leaves. Plant Sci. 2002, 163, 1137–1145. [Google Scholar] [CrossRef]
- Kim, J.-M.; To, T.K.; Matsui, A.; Tanoi, K.; Kobayashi, N.I.; Matsuda, F.; Habu, Y.; Ogawa, D.; Sakamoto, T.; Matsunaga, S.; et al. Acetate-mediated novel survival strategy against drought in plants. Nat. Plants 2017, 3, 17097. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, R.A.; Lei, J.; Afseth, C.A.; Castanha, C.; Wistrom, C.M.; Mortimer, J.C.; Jardine, K. Are Methanol-Derived Foliar Methyl Acetate Emissions a Tracer of Acetate-Mediated Drought Survival in Plants? Plants 2021, 10, 411. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Ashrafi, M.; Azimi-Moqadam, M.R.; MohseniFard, E.; Shekari, F.; Jafary, H.; Moradi, P.; Pucci, M.; Abate, G.; Mastinu, A. Physiological and Molecular Aspects of Two Thymus Species Differently Sensitive to Drought Stress. BioTech 2022, 11, 8. [Google Scholar] [CrossRef]
- Fletcher, J.S.; Beevers, H. Acetate metabolism in cell suspension cultures. Plant Physiol. 1970, 45, 765–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Yang, H.; Pangestu, F.; Nikolau, B.J. Failure to Maintain Acetate Homeostasis by Acetate-Activating Enzymes Impacts Plant Development. Plant Physiol. 2020, 182, 1256–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kok, B. On the interrelation of respiration and photosynthesis in green plants. Biochim. Biophys. Acta 1949, 3, 625–631. [Google Scholar] [CrossRef]
- Garcia, S.; Jardine, K.; Souza, V.; Souza, R.; Duvoisin, S., Jr.; Gonçalves, J. Reassimilation of Leaf Internal CO2 Contributes to Isoprene Emission in the Neotropical Species Inga edulis Mart. Forests 2019, 10, 472. [Google Scholar] [CrossRef] [Green Version]
- Heskel, M.A.; Atkin, O.K.; Turnbull, M.H.; Griffin, K.L. Bringing the Kok effect to light: A review on the integration of daytime respiration and net ecosystem exchange. Ecosphere 2013, 4, art98. [Google Scholar] [CrossRef]
- Yin, X.; Niu, Y.; van der Putten, P.E.L.; Struik, P.C. The Kok effect revisited. New Phytol. 2020, 227, 1764–1775. [Google Scholar] [CrossRef]
- Tcherkez, G.; Gauthier, P.; Buckley, T.N.; Busch, F.A.; Barbour, M.M.; Bruhn, D.; Heskel, M.A.; Gong, X.Y.; Crous, K.Y.; Griffin, K.; et al. Leaf day respiration: Low CO2 flux but high significance for metabolism and carbon balance. New Phytol. 2017, 216, 986–1001. [Google Scholar] [CrossRef]
- Tcherkez, G.; Gauthier, P.; Buckley, T.N.; Busch, F.A.; Barbour, M.M.; Bruhn, D.; Heskel, M.A.; Gong, X.Y.; Crous, K.; Griffin, K.L.; et al. Tracking the origins of the Kok effect, 70 years after its discovery. New Phytol. 2017, 214, 506–510. [Google Scholar] [CrossRef] [Green Version]
- Sauer, A.; Heise, K.P. On the light dependence of Fatty Acid synthesis in spinach chloroplasts. Plant Physiol. 1983, 73, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, P.P.G.; Saenz, N.; Griffin, K.L.; Way, D.; Tcherkez, G. Is the Kok effect a respiratory phenomenon? Metabolic insight using 13C labeling in Helianthus annuus leaves. New Phytol. 2020, 228, 1243–1255. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Fedorin, D.N.; Dobychina, M.A.; Igamberdiev, A.U. Regulation of expression of the mitochondrial and peroxisomal forms of citrate synthase in maize during germination and in response to light. Plant Sci. 2018, 272, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Plain, C.; Gerant, D.; Maillard, P.; Dannoura, M.; Dong, Y.; Zeller, B.; Priault, P.; Parent, F.; Epron, D. Tracing of recently assimilated carbon in respiration at high temporal resolution in the field with a tuneable diode laser absorption spectrometer after in situ 13CO2 pulse labelling of 20-year-old beech trees. Tree Physiol. 2009, 29, 1433–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinzer, F.C.; Brooks, J.R.; Domec, J.C.; Gartner, B.L.; Warren, J.M.; Woodruff, D.R.; Bible, K.; Shaw, D.C. Dynamics of water transport and storage in conifers studied with deuterium and heat tracing techniques. Plant Cell Environ. 2006, 29, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Seeger, S.; Weiler, M. Temporal dynamics of tree xylem water isotopes: In situ monitoring and modeling. Biogeosciences 2021, 18, 4603–4627. [Google Scholar] [CrossRef]
- Powers, E.M.; Marshall, J.D. Pulse labeling of dissolved 13C-carbonate into tree xylem: Developing a new method to determine the fate of recently fixed photosynthate. Rapid Commun. Mass Spectrom. 2011, 25, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Churchland, C.; Weatherall, A.; Briones, M.J.I.; Grayston, S.J. Stable-isotope labeling and probing of recent photosynthates into respired CO2, soil microbes and soil mesofauna using a xylem and phloem stem-injection technique on Sitka spruce (Picea sitchensis). Rapid Commun. Mass Spectrom. 2012, 26, 2493–2501. [Google Scholar] [CrossRef]
- Doccola, J.J.; Wild, P.M. Tree Injection as an Alternative Method of Insecticide Application. In Insecticides—Basic and Other Applications; InTech: Rijeka, Croatia, 2012; pp. 61–78. [Google Scholar]
- Navarro, C.; Fernández-Escobar, R.; Benlloch, M. A Low-pressure, Trunk-injection Method for Introducing Chemical Formulations into Olive Trees. J. Am. Soc. Hort. Sci. 1992, 117, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Reil, W.O.; Beutel, J. A pressure machine for injecting trees. Calif. Agric. 1976, 30, 4–5. [Google Scholar]
- Gauthier, P.P.G.; Battle, M.O.; Griffin, K.L.; Bender, M.L. Measurement of gross photosynthesis, respiration in the light, and mesophyll conductance using H218O labeling. Plant Physiol. 2018, 177, 62–74. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Velikova, V.; Di Marco, G. Respiration in the light measured by 12CO2 emission in 13CO2 atmosphere in maize leaves. Funct. Plant Biol. 2001, 28, 1103. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Busch, F.A. Changes in the chloroplastic CO2 concentration explain much of the observed Kok effect: A model. New Phytol. 2017, 214, 570–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tcherkez, G.; Boex-Fontvieille, E.; Mahé, A.; Hodges, M. Respiratory carbon fluxes in leaves. Curr. Opin. Plant Biol. 2012, 15, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Tcherkez, G.; Cornic, G.; Bligny, R.; Gout, E.; Ghashghaie, J. In vivo respiratory metabolism of illuminated leaves. Plant Physiol. 2005, 138, 1596–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tcherkez, G.; Mahé, A.; Gauthier, P.; Mauve, C.; Gout, E.; Bligny, R.; Cornic, G.; Hodges, M. In folio respiratory fluxomics revealed by 13C-isotopic labeling and H/D isotope effects highlight the noncyclic nature of the tricarboxylic acid “cycle” in illuminated leaves. Plant Physiol. 2009, 151, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Oliver, D.J.; Nikolau, B.J.; Wurtele, E.S. Acetyl-CoA—Life at the metabolic nexus. Plant Sci. 2009, 176, 597–601. [Google Scholar] [CrossRef]
- Schwer, B.; Bunkenborg, J.; Verdin, R.O.; Andersen, J.S.; Verdin, E. Reversible lysine acetylation controls the activity of the mitochondrial enzyme acetyl-CoA synthetase 2. Proc. Natl. Acad. Sci. USA 2006, 103, 10224–10229. [Google Scholar] [CrossRef] [Green Version]
- Kunze, M.; Pracharoenwattana, I.; Smith, S.M.; Hartig, A. A central role for the peroxisomal membrane in glyoxylate cycle function. Biochim. Biophys. Acta 2006, 1763, 1441–1452. [Google Scholar] [CrossRef] [Green Version]
- Beevers, H. Metabolic production of sucrose from fat. Nature 1961, 191, 433–436. [Google Scholar] [CrossRef]
- Eastmond, P.J.; Germain, V.; Lange, P.R.; Bryce, J.H.; Smith, S.M.; Graham, I.A. Postgerminative growth and lipid catabolism in oilseeds lacking the glyoxylate cycle. Proc. Natl. Acad. Sci. USA 2000, 97, 5669–5674. [Google Scholar] [CrossRef] [Green Version]
- Füßl, M.; König, A.-C.; Eirich, J.; Hartl, M.; Kleinknecht, L.; Bohne, A.-V.; Harzen, A.; Kramer, K.; Leister, D.; Nickelsen, J.; et al. Dynamic light- and acetate-dependent regulation of the proteome and lysine acetylome of Chlamydomonas. Plant J. 2022, 109, 261–277. [Google Scholar] [CrossRef]
- Jardine, K.; Chambers, J.; Alves, E.G.; Teixeira, A.; Garcia, S.; Holm, J.; Higuchi, N.; Manzi, A.; Abrell, L.; Fuentes, J.D.; et al. Dynamic balancing of isoprene carbon sources reflects photosynthetic and photorespiratory responses to temperature stress. Plant Physiol. 2014, 166, 2051–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jardine, K.J.; Lei, J.; Som, S.; Souza, D.; Clendinen, C.S.; Mehta, H.; Handakumbura, P.; Bill, M.; Young, R.P. Light-Dependence of Formate (C1) and Acetate (C2) Transport and Oxidation in Poplar Trees. Plants 2022, 11, 2080. https://doi.org/10.3390/plants11162080
Jardine KJ, Lei J, Som S, Souza D, Clendinen CS, Mehta H, Handakumbura P, Bill M, Young RP. Light-Dependence of Formate (C1) and Acetate (C2) Transport and Oxidation in Poplar Trees. Plants. 2022; 11(16):2080. https://doi.org/10.3390/plants11162080
Chicago/Turabian StyleJardine, Kolby J., Joseph Lei, Suman Som, Daisy Souza, Chaevien S. Clendinen, Hardeep Mehta, Pubudu Handakumbura, Markus Bill, and Robert P. Young. 2022. "Light-Dependence of Formate (C1) and Acetate (C2) Transport and Oxidation in Poplar Trees" Plants 11, no. 16: 2080. https://doi.org/10.3390/plants11162080