
Genome-Wide Identification and Expression Analysis of Dof Transcription Factors in Lotus (Nelumbo nucifera Gaertn.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Physiochemical Characteristics of NnDofs
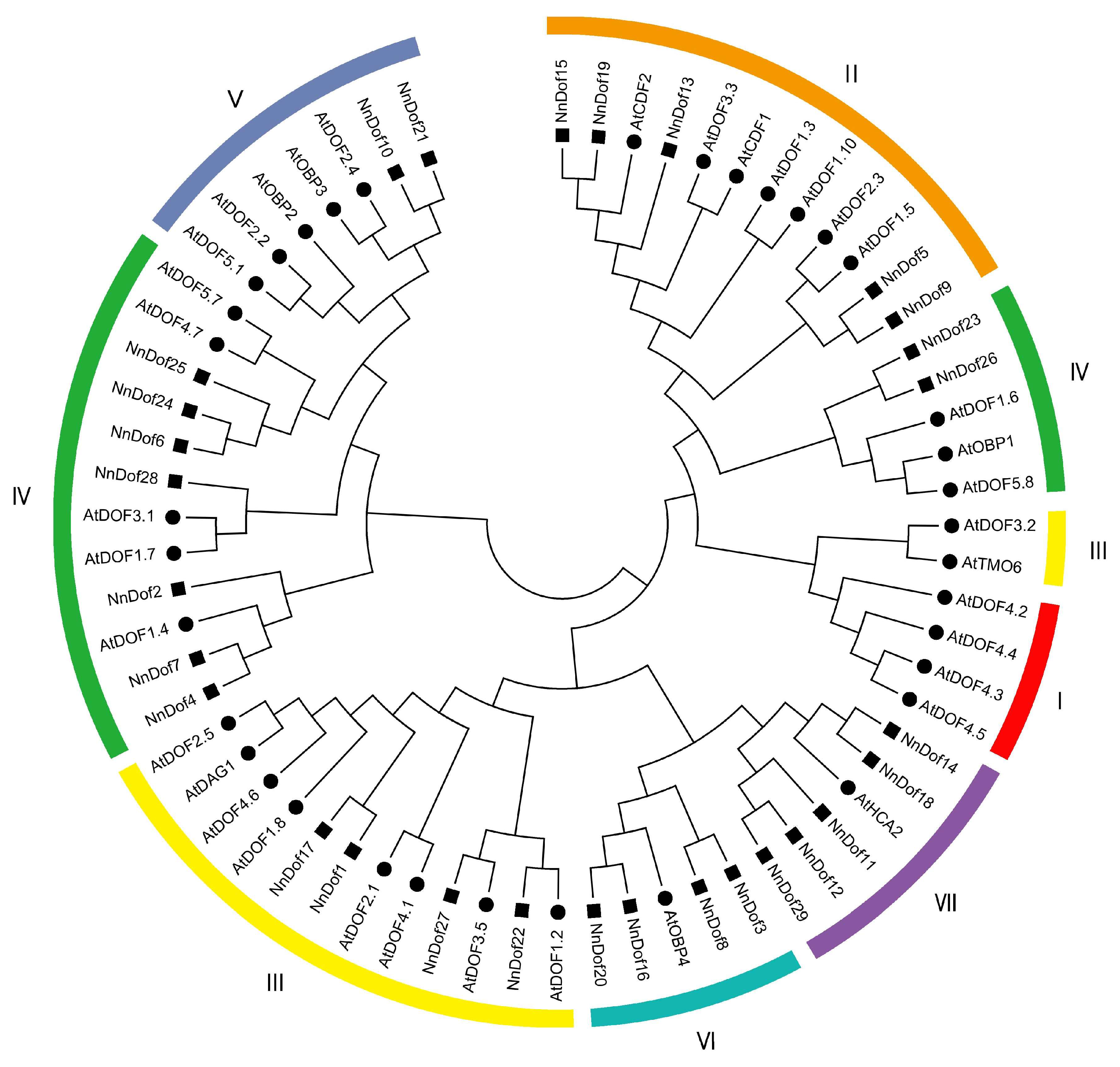
2.2. Phylogenetic Analysis and Classification of NnDofs
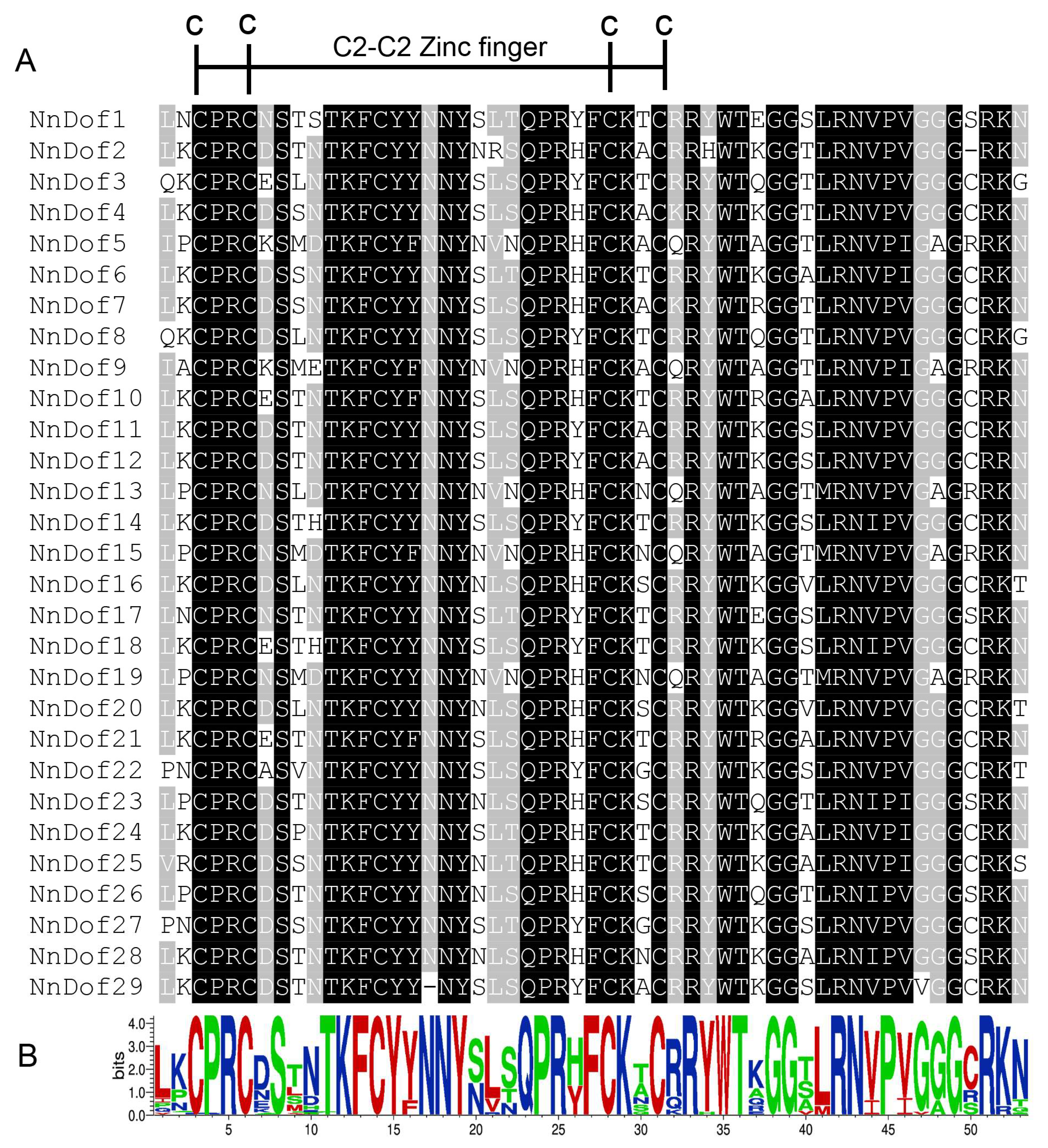
2.3. Gene Structure and Conserved Motifs of NnDofs
2.4. Promoter Analysis of NnDof Genes
2.5. Interaction Network Analysis of NnDofs
2.6. Chromosome Distribution, Gene Duplication and Synteny Analysis of NnDofs
2.7. Expression Patterns of NnDofs in Lotus
3. Discussion
3.1. Identification, Structural and Phylogenetic Analysis, and Evolutionary Characterization of NnDofs
3.2. Expression Profiling of NnDofs
4. Conclusions
5. Materials and Methods
5.1. Identification and Physical Properties Analysis of NnDofs
5.2. Conserved Domain, Conserved Structure, and Phylogenetic Analysis of NnDofs
5.3. Gene Duplication and Syntenic Analysis of NnDofs
5.4. Cis-Acting Element Analysis and Protein Interaction Network Prediction of NnDofs
5.5. Expression Patterns Analysis of NnDofs by RNA-Seq Data
5.6. Stress Treatments and Quantitative Real-Time PCR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, J.; Pu, L.; Han, M.; Zhu, M.; Zhang, R.; Xiang, Y. Soil Salinization Research in China: Advances and Prospects. J. Geogr. Sci. 2014, 24, 943–960. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil Salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as One of the Tools for Its Alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative Stress and Antioxidant Defense Mechanisms in Plants under Salt Stress. In Plant Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019; pp. 191–205. [Google Scholar]
- Shah, W.H.; Rasool, A.; Saleem, S.; Mushtaq, N.U.; Tahir, I.; Hakeem, K.R.; Rehman, R.U. Understanding the Integrated Pathways and Mechanisms of Transporters, Protein Kinases, and Transcription Factors in Plants under Salt Stress. Int. J. Genom. 2021, 2021, 5578727. [Google Scholar] [CrossRef]
- Golldack, D.; Lüking, I.; Yang, O. Plant Tolerance to Drought and Salinity: Stress Regulating Transcription Factors and Their Functional Significance in the Cellular Transcriptional Network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Boggon, T.J.; Shan, W.-S.; Santagata, S.; Myers, S.C.; Shapiro, L. Implication of Tubby Proteins as Transcription Factors by Structure-Based Functional Analysis. Science 1999, 286, 2119–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Malviya, N.; Kushwaha, H.; Nasim, J.; Bisht, N.C.; Singh, V.K.; Yadav, D. Insights into Structural and Functional Diversity of Dof (DNA Binding with One Finger) Transcription Factor. Planta 2015, 241, 549–562. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H.-Y. Functional Analysis of a Calcium-Binding Transcription Factor Involved in Plant Salt Stress Signaling. FEBS Lett. 2006, 580, 5251–5256. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Suzuki, K.; Ohtsuki, N.; Tsujimoto, Y.; Fujimura, T.; Shinshi, H. Identification of Genes of the Plant-Specific Transcription-Factor Families Cooperatively Regulated by Ethylene and Jasmonate in Arabidopsis Thaliana. J. Plant Res. 2006, 119, 407–413. [Google Scholar] [CrossRef]
- Yanagisawa, S.; Izui, K. Molecular Cloning of Two DNA-Binding Proteins of Maize That Are Structurally Different but Interact with the Same Sequence Motif. J. Biol. Chem. 1993, 268, 16028–16036. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, Y.; Zhang, C.; Zhang, T.; Hu, T.; Ye, J.; Zhang, J.; Wang, T.; Li, H.; Ye, Z. Genome-wide Analysis of Plant-specific Dof Transcription Factor Family in Tomato. J. Integr. Plant Biol. 2013, 55, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Zhu, J.; Zhang, X. Genome-Wide Identification and Characterization of the Dof Gene Family in Cassava (Manihot esculenta). Gene 2019, 687, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, X.; Huan, L.; Sun, M.; Liu, L.; Chen, X.; Gao, D.; Li, L. Genome-Wide Analysis of Dof Family Genes and Their Expression during Bud Dormancy in Peach (Prunus Persica). Sci. Hortic. 2017, 214, 18–26. [Google Scholar] [CrossRef]
- Wu, Z.; Cheng, J.; Cui, J.; Xu, X.; Liang, G.; Luo, X.; Chen, X.; Tang, X.; Hu, K.; Qin, C. Genome-Wide Identification and Expression Profile of Dof Transcription Factor Gene Family in Pepper (Capsicum Annuum L.). Front. Plant Sci. 2016, 7, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagisawa, S. Structure, Function, and Evolution of the Dof Transcription Factor Family. In Plant Transcription Factors; Elsevier: Amsterdam, The Netherlands, 2016; pp. 183–197. [Google Scholar]
- Fang, Z.; Jiang, W.; He, Y.; Ma, D.; Liu, Y.; Wang, S.; Zhang, Y.; Yin, J. Genome-Wide Identification, Structure Characterization, and Expression Profiling of Dof Transcription Factor Gene Family in Wheat (Triticum Aestivum L.). Agronomy 2020, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, S. The Dof Family of Plant Transcription Factors. Trends Plant Sci. 2002, 7, 555–560. [Google Scholar] [CrossRef]
- Lijavetzky, D.; Carbonero, P.; Vicente-Carbajosa, J. Genome-Wide Comparative Phylogenetic Analysis of the Rice and Arabidopsis Dof Gene Families. BMC Evol. Biol. 2003, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Risueno, M.Á.; Martínez, M.; Vicente-Carbajosa, J.; Carbonero, P. The Family of DOF Transcription Factors: From Green Unicellular Algae to Vascular Plants. Mol. Genet. Genom. 2007, 277, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Ruta, V.; Longo, C.; Lepri, A.; De Angelis, V.; Occhigrossi, S.; Costantino, P.; Vittorioso, P. The DOF Transcription Factors in Seed and Seedling Development. Plants 2020, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.-H.; Kim, S.; Lee, H.-A.; Choi, D.; Yeom, S.-I. Genome-Wide Analysis of Dof Transcription Factors Reveals Functional Characteristics during Development and Response to Biotic Stresses in Pepper. Sci. Rep. 2016, 6, 33332. [Google Scholar] [CrossRef] [Green Version]
- Corrales, A.-R.; Carrillo, L.; Lasierra, P.; Nebauer, S.G.; Dominguez-Figueroa, J.; Renau-Morata, B.; Pollmann, S.; Granell, A.; Molina, R.-V.; Vicente-Carbajosa, J.; et al. Multifaceted Role of Cycling DOF Factor 3 (CDF3) in the Regulation of Flowering Time and Abiotic Stress Responses in Arabidopsis. Plant Cell Environ. 2017, 40, 748–764. [Google Scholar] [CrossRef] [PubMed]
- Goralogia, G.S.; Liu, T.-K.; Zhao, L.; Panipinto, P.M.; Groover, E.D.; Bains, Y.S.; Imaizumi, T. CYCLING DOF FACTOR 1 Represses Transcription through the TOPLESS Co-repressor to Control Photoperiodic Flowering in Arabidopsis. Plant J. 2017, 92, 244–262. [Google Scholar] [CrossRef]
- Xu, X.; Li, F.; Wang, Y.; Tang, S.; Dai, Q.; Zhu, S.; Liu, T. Identification of Dof Transcription Factors in Ramie (Boehmeria Nivea L. Gaud) and Their Expression in Response to Different Nitrogen Treatments. 3 Biotech 2018, 8, 496. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Lin, J.; Li, Z.; Lin, Z.; Ma, Y.; Wang, Y.; Ming, R. Allele Specific Expression of Dof Genes Responding to Hormones and Abiotic Stresses in Sugarcane. PLoS ONE 2020, 15, e0227716. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, N.; Deng, X.; Liu, D.; Li, M.; Cui, D.; Hu, Y.; Yan, Y. Genome-Wide Analysis of Wheat DNA-Binding with One Finger (Dof) Transcription Factor Genes: Evolutionary Characteristics and Diverse Abiotic Stress Responses. BMC Genom. 2020, 21, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yuan, L.; Liu, X.; Chen, X.; Wang, X. Evolution Analysis of Dof Transcription Factor Family and Their Expression in Response to Multiple Abiotic Stresses in Malus Domestica. Gene 2018, 639, 137–148. [Google Scholar] [CrossRef]
- Ramirez-Parra, E.; Perianez-Rodriguez, J.; Navarro-Neila, S.; Gude, I.; Moreno-Risueno, M.A.; del Pozo, J.C. The Transcription Factor OBP4 Controls Root Growth and Promotes Callus Formation. New Phytol. 2017, 213, 1787–1801. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Li, S.; Zhu, Y.; Zhao, Q.; Zhu, D.; Yu, J. ZmDof3, a Maize Endosperm-Specific Dof Protein Gene, Regulates Starch Accumulation and Aleurone Development in Maize Endosperm. Plant Mol. Biol. 2017, 93, 7–20. [Google Scholar] [CrossRef]
- Wu, J.; Chen, L.; Chen, M.; Zhou, W.; Dong, Q.; Jiang, H.; Cheng, B. The DOF-Domain Transcription Factor ZmDOF36 Positively Regulates Starch Synthesis in Transgenic Maize. Front. Plant Sci. 2019, 10, 465. [Google Scholar] [CrossRef] [PubMed]
- Fornara, F.; Panigrahi, K.C.; Gissot, L.; Sauerbrunn, N.; Rühl, M.; Jarillo, J.A.; Coupland, G. Arabidopsis DOF Transcription Factors Act Redundantly to Reduce CONSTANS Expression and Are Essential for a Photoperiodic Flowering Response. Dev. Cell 2009, 17, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Liang, W.; Liu, Z.; Wang, Y.; Zhao, Y.; Ijaz, B.; Hua, J. Overexpression of GhDof1 Improved Salt and Cold Tolerance and Seed Oil Content in Gossypium Hirsutum. J. Plant Physiol. 2017, 218, 222–234. [Google Scholar] [CrossRef]
- Zhou, Y.; Cheng, Y.; Wan, C.; Li, J.; Yang, Y.; Chen, J. Genome-Wide Characterization and Expression Analysis of the Dof Gene Family Related to Abiotic Stress in Watermelon. PeerJ 2020, 8, e8358. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Wang, J.; Chen, X.; Wang, F.; Peng, P.; Zhou, Y.; Miao, Y.; Zhang, Y.; Gao, Y.; Qi, Y. Rice Os DOF 15 Contributes to Ethylene-inhibited Primary Root Elongation under Salt Stress. New Phytol. 2019, 223, 798–813. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Zhang, C.; Shu, W.; Ye, Z.; Li, H.; Zhang, Y. The Transcription Factor SlDof22 Involved in Ascorbate Accumulation and Salinity Stress in Tomato. Biochem. Biophys. Res. Commun. 2016, 474, 736–741. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, F.-L.; Zheng, X.-W.; Hu, M.-L.; Dong, C.; Diao, Y.; Wang, Y.-W.; Xie, K.-Q.; Hu, Z.-L. Comparative Population Genomics Reveals Genetic Divergence and Selection in Lotus, Nelumbo Nucifera. BMC Genom. 2020, 21, 146. [Google Scholar] [CrossRef]
- Shen-Miller, J. Sacred lotus, the long-living fruits of China Antique. Seed Sci. Res. 2002, 12, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhu, L.; Pan, C.; Xu, L.; Liu, Y.; Ke, W.; Yang, P. Transcriptomic Analysis of the Regulation of Rhizome Formation in Temperate and Tropical Lotus (Nelumbo nucifera). Sci. Rep. 2015, 5, 13059. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.R.; Gautam, L.N.S.; Adhikari, D.; Karki, R. A Comprehensive Review on Chemical Profiling of Nelumbo Nucifera: Potential for Drug Development. Phytother. Res. 2017, 31, 3–26. [Google Scholar] [CrossRef]
- Sohn, D.-H.; Kim, Y.-C.; Oh, S.-H.; Park, E.-J.; Li, X.; Lee, B.-H. Hepatoprotective and Free Radical Scavenging Effects of Nelumbo Nucifera. Phytomedicine 2003, 10, 165–169. [Google Scholar] [CrossRef]
- Liu, L.; White, M.J.; MacRae, T.H. Transcription Factors and Their Genes in Higher Plants: Functional Domains, Evolution and Regulation. Eur. J. Biochem. 1999, 262, 247–257. [Google Scholar] [CrossRef]
- Zou, H.-F.; Zhang, Y.-Q.; Wei, W.; Chen, H.-W.; Song, Q.-X.; Liu, Y.-F.; Zhao, M.-Y.; Wang, F.; Zhang, B.-C.; Lin, Q. The Transcription Factor AtDOF4. 2 Regulates Shoot Branching and Seed Coat Formation in Arabidopsis. Biochem. J. 2013, 449, 373–388. [Google Scholar] [CrossRef]
- Skirycz, A.; Jozefczuk, S.; Stobiecki, M.; Muth, D.; Zanor, M.I.; Witt, I.; Mueller-Roeber, B. Transcription Factor AtDOF4; 2 Affects Phenylpropanoid Metabolism in Arabidopsis Thaliana. New Phytol. 2007, 175, 425–438. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.-Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R. Arabidopsis Transcription Factors: Genome-Wide Comparative Analysis among Eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Qu, L.-J.; Zhu, Y.-X. Transcription Factor Families in Arabidopsis: Major Progress and Outstanding Issues for Future Research. Curr. Opin. Plant Biol. 2006, 9, 544–549. [Google Scholar] [CrossRef]
- Lynch, M.; Force, A. The Probability of Duplicate Gene Preservation by Subfunctionalization. Genetics 2000, 154, 459–473. [Google Scholar] [CrossRef]
- Jourda, C.; Cardi, C.; Mbéguié-A.-Mbéguié, D.; Bocs, S.; Garsmeur, O.; D’Hont, A.; Yahiaoui, N. Expansion of Banana (Musa acuminata) Gene Families Involved in Ethylene Biosynthesis and Signalling after Lineage-Specific Whole-Genome Duplications. New Phytol. 2014, 202, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Koszul, R.; Fischer, G. A Prominent Role for Segmental Duplications in Modeling Eukaryotic Genomes. Comptes Rendus Biol. 2009, 332, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Yin, H.; Li, L.; Wang, R.; Wu, J.; Wu, J.; Zhang, S. Different Modes of Gene Duplication Show Divergent Evolutionary Patterns and Contribute Differently to the Expansion of Gene Families Involved in Important Fruit Traits in Pear (Pyrus bretschneideri). Front. Plant Sci. 2018, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Qin, G.; Gu, H.; Qu, L.-J. Dof5.6/HCA2, a Dof Transcription Factor Gene, Regulates Interfascicular Cambium Formation and Vascular Tissue Development in Arabidopsis. Plant Cell 2009, 21, 3518–3534. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Yanagisawa, S. Sequential Activation of Two Dof Transcription Factor Gene Promoters during Vascular Development in Arabidopsis Thaliana. Plant Physiol. Biochem. 2007, 45, 623–629. [Google Scholar] [CrossRef]
- Rymen, B.; Kawamura, A.; Schäfer, S.; Breuer, C.; Iwase, A.; Shibata, M.; Ikeda, M.; Mitsuda, N.; Koncz, C.; Ohme-Takagi, M. ABA Suppresses Root Hair Growth via the OBP4 Transcriptional Regulator. Plant Physiol. 2017, 173, 1750–1762. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.-h.; Han, Y.-c.; Xiao, Y.-y.; Kuang, J.-f.; Fan, Z.-q.; Chen, J.-y.; Lu, W.-j. The banana fruit Dof transcription factor MaDof23 acts as a repressor and interacts with MaERF9 in regulating ripening-related genes. J. Exp. Bot. 2016, 67, 2263–2275. [Google Scholar] [CrossRef] [Green Version]
- Lohani, N.; Babaei, S.; Singh, M.B.; Bhalla, P.L. Genome-Wide in Silico Identification and Comparative Analysis of Dof Gene Family in Brassica napus. Plants 2021, 10, 709. [Google Scholar] [CrossRef]
- Jiao, Z.; Tang, D.; Fan, K.; Zhang, Q.; Liu, M.-Y.; Ruan, J. Genome-wide identification of the DNA-binding one zinc finger (Dof) transcription factor gene family and their potential functioning in nitrogen remobilization in tea plant (Camellia sinensis L.). Sci. Hortic. 2022, 292, 110615. [Google Scholar] [CrossRef]
- Liu, J.-H.; Peng, T.; Dai, W. Critical Cis-Acting Elements and Interacting Transcription Factors: Key Players Associated with Abiotic Stress Responses in Plants. Plant Mol. Biol. Report. 2014, 32, 303–317. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and Validation of Promoters and Cis-Acting Regulatory Elements. Plant Sci. 2014, 217–218. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Li, S.; Hussain, J.; Xu, X.; Yin, J.; Zhang, Y.; Chen, X.; Li, L. Isolation and functional characterization of a salt responsive transcriptional factor, LrbZIP from lotus root (Nelumbo nucifera Gaertn). Mol. Biol. Rep. 2013, 40, 4033–4045. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Shi, H.; Wang, Y.; Chen, S.; Deng, J.; Liu, Y.; Li, S.; Chan, Z. Comparative physiological analysis of lotus (Nelumbo nucifera) cultivars in response to salt stress and cloning of NnCIPK genes. Sci. Hortic. 2014, 173, 29–36. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, Y.; Zhao, S.; Li, L. Comprehensive analysis of evolutionary characterization and expression for monosaccharide transporter family genes in Nelumbo nucifera. Front. Ecol. Evol. 2021, 9, 537398. [Google Scholar] [CrossRef]
- Li, J.; Xiong, Y.; Li, Y.; Ye, S.; Yin, Q.; Gao, S.; Yang, D.; Yang, M.; Palva, E.T.; Deng, X. Comprehensive analysis and functional studies of WRKY transcription factors in Nelumbo nucifera. Int. J. Mol. Sci. 2019, 20, 5006. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Cao, J. Comparative Analysis of Dof Transcription Factor Family in Maize. Plant Mol. Biol. Report. 2015, 33, 1245–1258. [Google Scholar] [CrossRef]
- Li, H.; Yang, X.; Zhang, Y.; Gao, Z.; Liang, Y.; Chen, J.; Shi, T. Nelumbo Genome Database, an Integrative Resource for Gene Expression and Variants of Nelumbo Nucifera. Sci. Data 2021, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Wan, W.; Mao, H.; Yin, D.; Deng, X.; Yan, H.; Ren, L. Genome-Wide Identification and Expression Analysis of Dof Transcription Factors in Lotus (Nelumbo nucifera Gaertn.). Plants 2022, 11, 2057. https://doi.org/10.3390/plants11152057
Cao X, Wan W, Mao H, Yin D, Deng X, Yan H, Ren L. Genome-Wide Identification and Expression Analysis of Dof Transcription Factors in Lotus (Nelumbo nucifera Gaertn.). Plants. 2022; 11(15):2057. https://doi.org/10.3390/plants11152057
Chicago/Turabian StyleCao, Xiaohan, Wenyang Wan, Huimin Mao, Dandan Yin, Xianhui Deng, Huan Yan, and Liping Ren. 2022. "Genome-Wide Identification and Expression Analysis of Dof Transcription Factors in Lotus (Nelumbo nucifera Gaertn.)" Plants 11, no. 15: 2057. https://doi.org/10.3390/plants11152057