
Identification and Expression Analysis of NAC Gene Family in Weeping Trait of Lagerstroemia indica


Abstract
:1. Introduction
2. Results
2.1. Identification of NAC Genes in L. indica
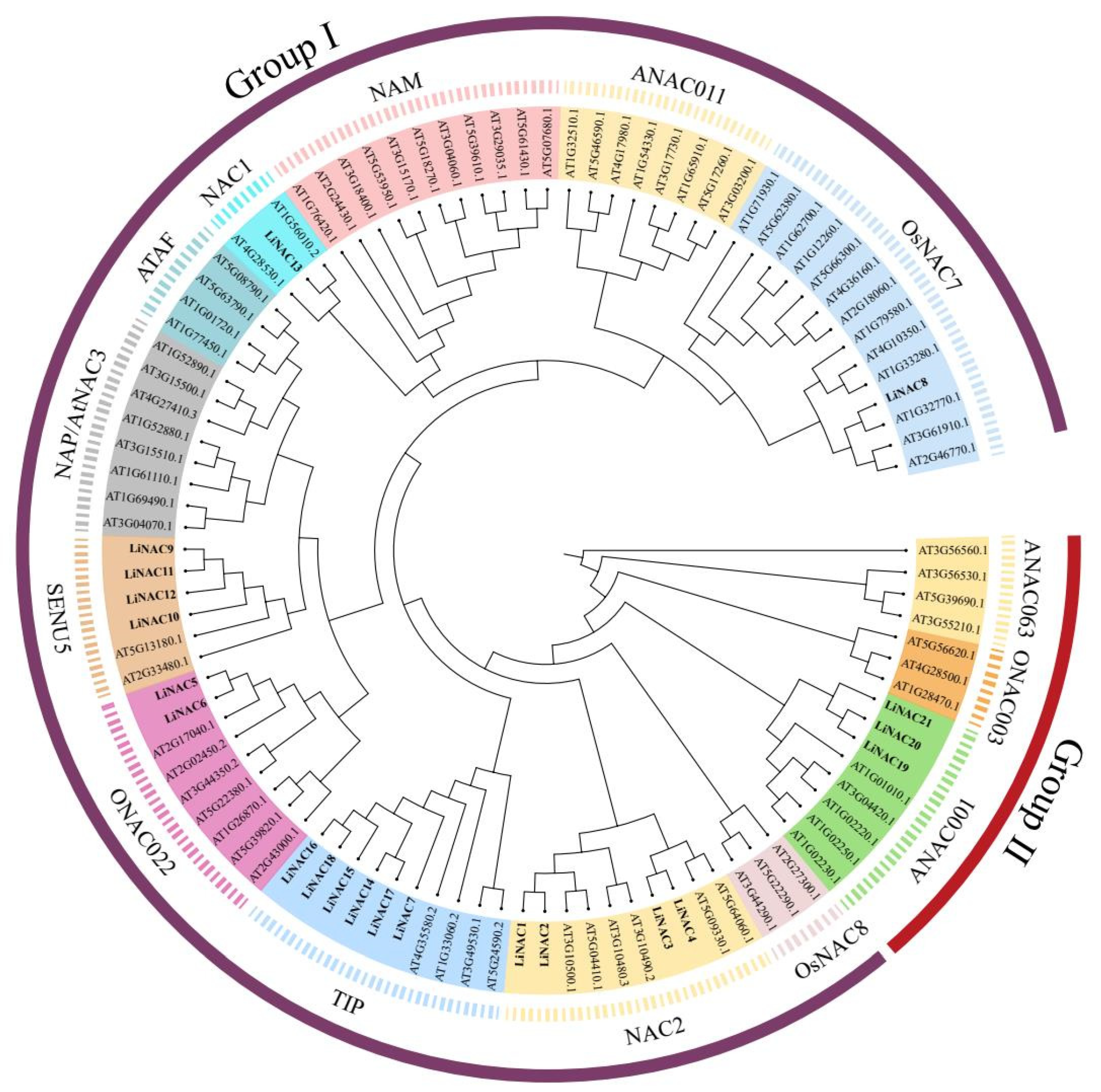
2.2. Phylogenetic Analysis and Classification of LiNAC Proteins
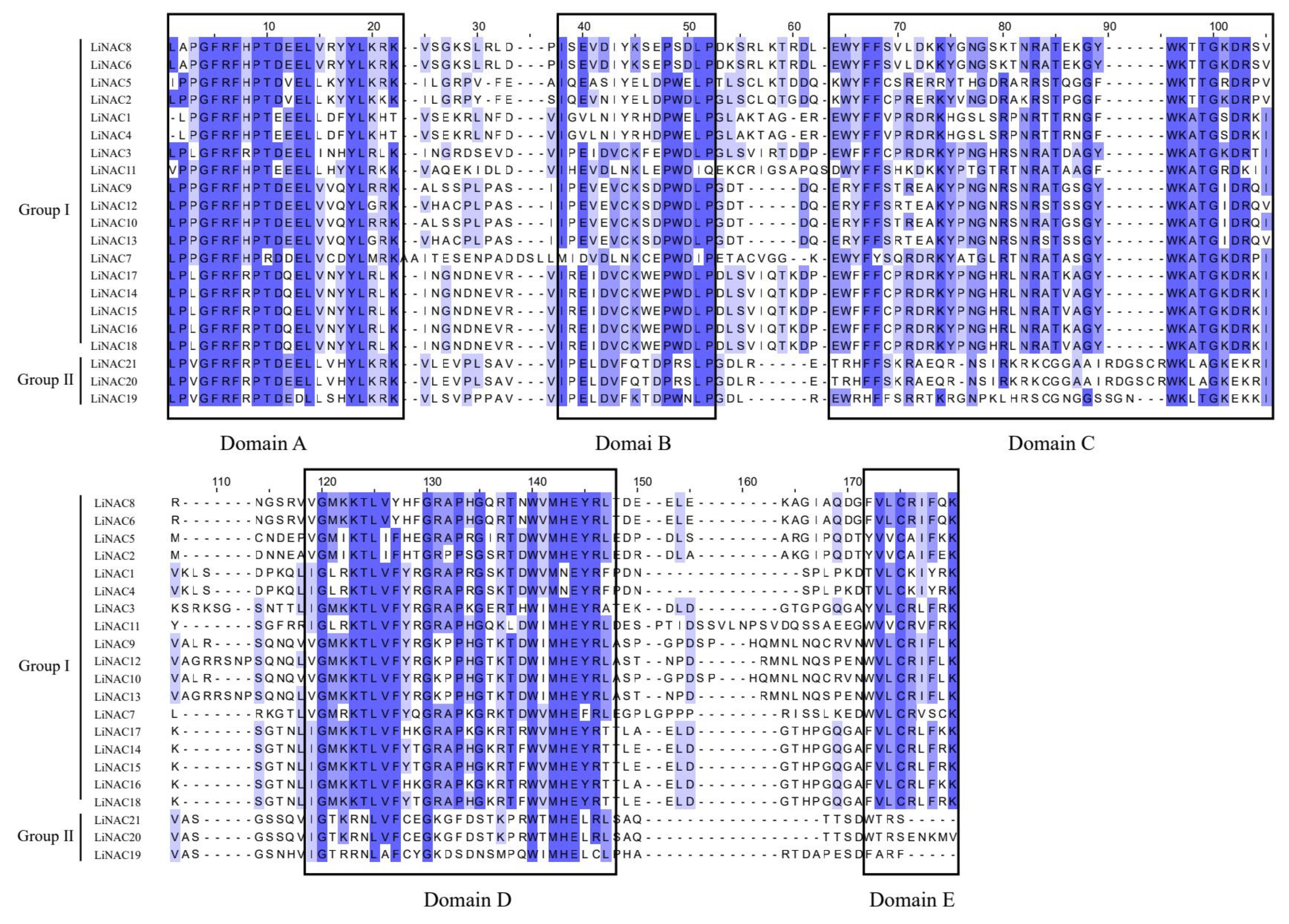
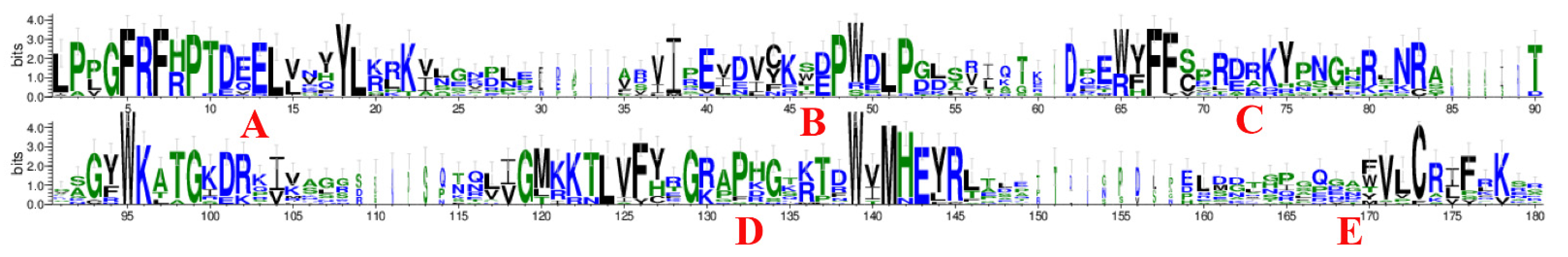
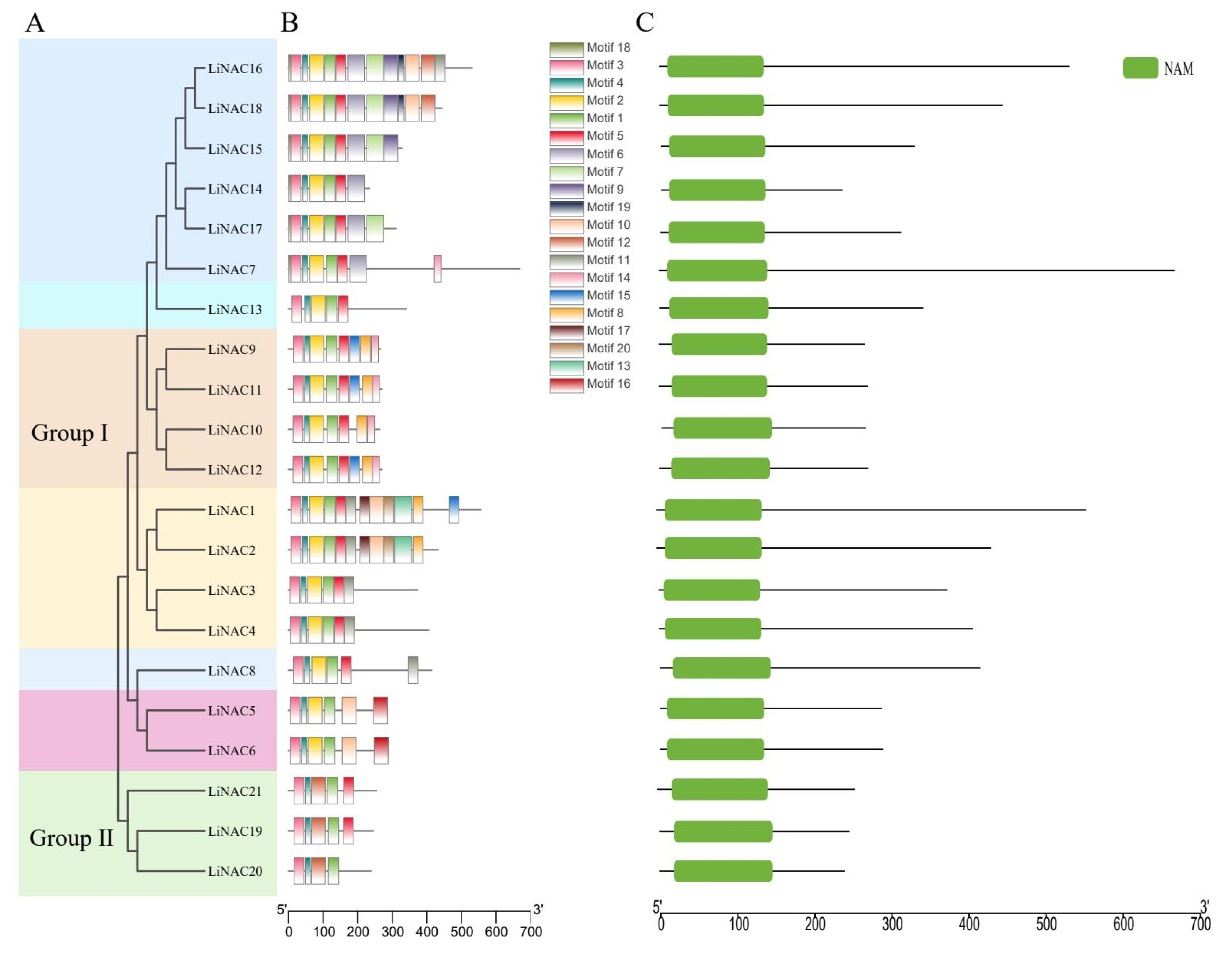
2.3. Conserved Sequence Alignment and Motif Analysis
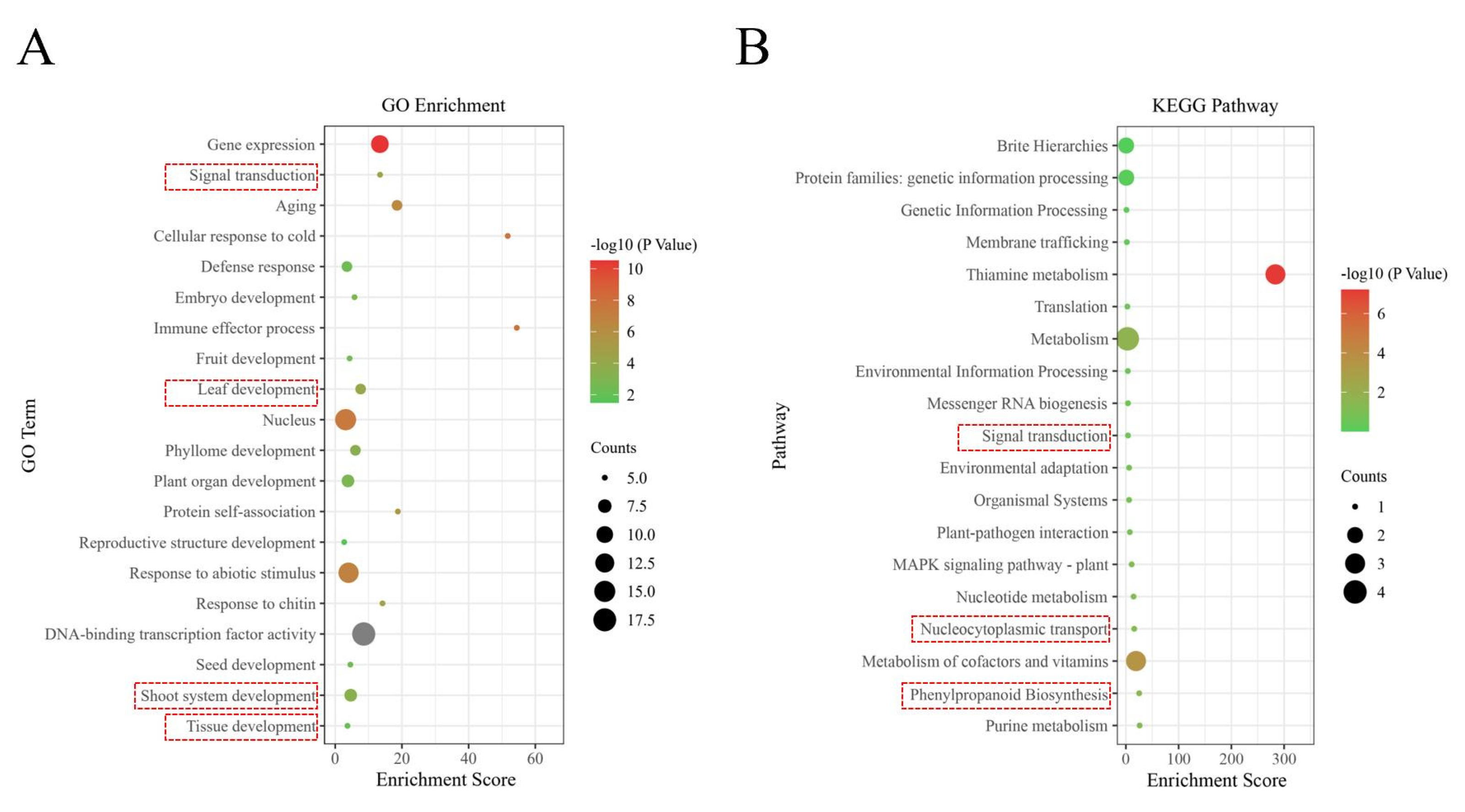
2.4. Annotation and Enrichment Analysis of LiNAC Genes
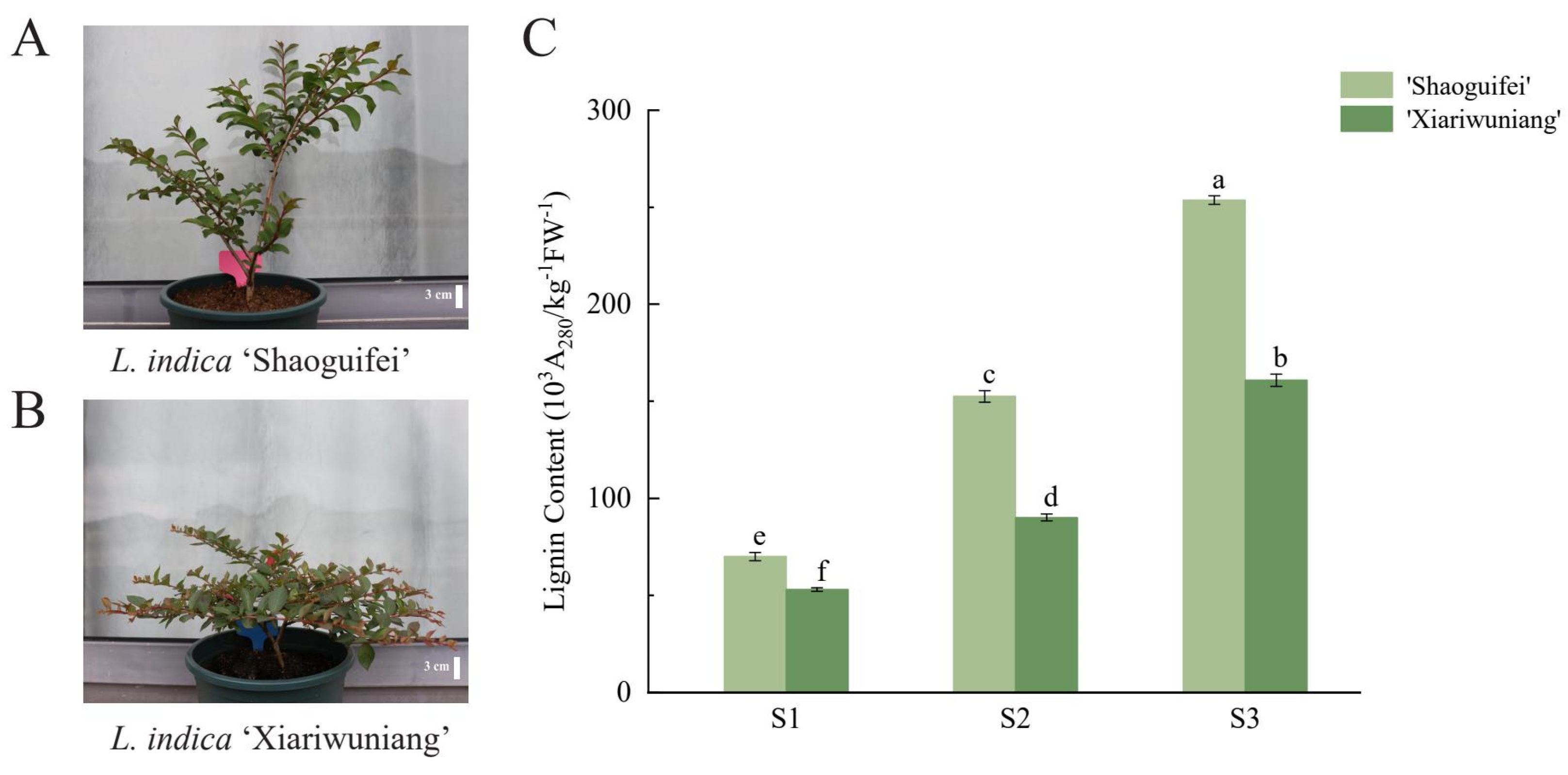
2.5. Determination and Analysis of Lignin Content
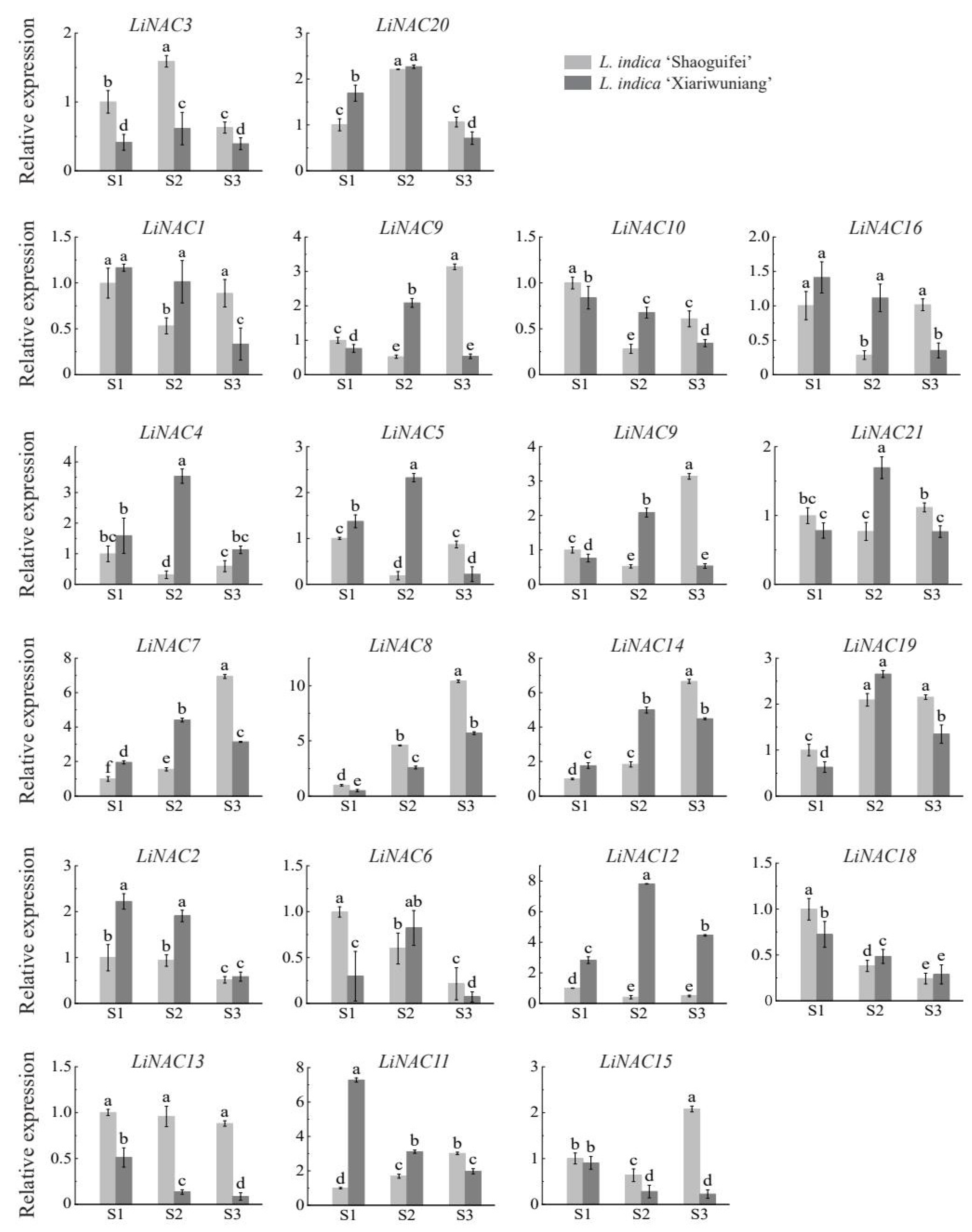
2.6. Expression Analysis of LiNAC Genes in Regulating the Weeping Trait
3. Discussion
3.1. The Characteristics of NAC Gene Family in L. indica
3.2. LiNACs Are Closely Associated with the Weeping Trait of L. indica
4. Materials and Methods
4.1. Identification and Sequence Analysis of NAC Genes from L. indica
4.2. Characterization of LiNAC Proteins
4.3. Sequence Alignment and Phylogenetic Tree Construction
4.4. Annotation and Enrichment Analysis in GO and KEGG Databases
4.5. Determination of Lignin Content
4.6. Plant Materials, RNA Extraction, and qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, J.L.; Hollender, C.A. Branching out: New insights into the genetic regulation of shoot architecture in trees. Curr. Opin. Plant Biol. 2019, 47, 73–80. [Google Scholar] [CrossRef]
- Hollender, C.A.; Pascal, T.; Tabb, A.; Hadiarto, T.; Srinivasan, C.; Wang, W.; Liu, Z.; Scorza, R.; Dardick, C. Loss of a highly conserved sterile alpha motif domain gene (WEEP) results in pendulous branch growth in peach trees. Proc. Natl. Acad. Sci. USA 2018, 115, e4690–e4699. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zeng, Y.; Yan, P.; He, C.; Zhang, J. Transcriptional and hormonal regulation of weeping trait in Salix matsudana. Genes 2017, 8, 359. [Google Scholar] [CrossRef] [Green Version]
- Hou, D.; Zhang, J.; Zheng, T.C.; Zhang, Q.X. A study about branch physical structure of upright/weeping Mei (Prunus mume Sieb. et Zucc). Adv. Ornament Hortic. China 2017, 2017, 552–559. [Google Scholar]
- Mao, T.Y.; Zhu, H.H.; Liu, Y.; Bao, M.Z.; Zhang, J.W.; Fu, Q.; Xiong, C.F.; Zhang, J. Weeping candidate genes screened using comparative transcriptomic analysis of weeping and upright progeny in an F1 population of Prunus mume. Physiol. Plant 2020, 170, 318–334. [Google Scholar] [CrossRef]
- Li, L.L.; Zhang, Y.C.; Zheng, T.C.; Zhuo, X.K.; Li, P.; Qiu, L.K.; Liu, W.C.; Wang, J.; Cheng, T.R.; Zhang, Q.X. Comparative gene expression analysis reveals that multiple mechanisms regulate the weeping trait in Prunus mume. Sci. Rep. 2021, 11, 2675. [Google Scholar] [CrossRef]
- Zheng, T.C.; Li, P.; Zhuo, X.K.; Liu, W.C.; Qiu, L.K.; Li, L.L.; Yuan, C.Q.; Sun, L.D.; Zhang, Z.Y.; Wang, J.; et al. The chromosome-level genome provides insight into the molecular mechanism underlying the tortuous-branch phenotype of Prunus mume. New Phytol. 2022, 235, 141–156. [Google Scholar] [CrossRef]
- Joshi, C.P.; Thammannagowda, S.; Fujino, T.; Gou, J.Q.; Avci, U.; Haigler, C.H.; McDonnell, L.M.; Mansfield, S.D.; Mengesha, B.; Carpita, N.C.; et al. Perturbation of wood cellulose synthesis causes pleiotropic effects in transgenic aspen. Mol. Plant 2011, 4, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.K.; Kjaersgaard, T.; Nielsen, M.M.; Galberg, P.; Petersen, K.; O’Shea, C.; Skriver, K. The Arabidopsis thaliana NAC transcription factor family: Structure-function relationships and determinants of ANAC019 stress signaling. Biochem. J. 2010, 426, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Souer, E.; Van, H.A.; Kloos, D.; Mol, J.; Koes, R. The no apical meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Melo, B.P.; Lourenço-Tessutti, I.T.; Fraga, O.T.; Pinheiro, L.B.; de Jesus Lins, C.B.; Morgante, C.V.; Engler, J.A.; Reis, P.A.B.; Grossi-de-Sá, M.F.; Fontes, E.P.B. Contrasting roles of GmNAC065 and GmNAC085 in natural senescence, plant development, multiple stresses and cell death responses. Sci. Rep. 2021, 11, 11178. [Google Scholar] [CrossRef]
- Hu, P.; Zhang, K.M.; Yang, C.P. BpNAC012 positively regulates abiotic stress responses and secondary wall biosynthesis. Plant Physiol. 2019, 179, 700–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.Q.; Richardson, E.A.; Ye, Z.H. Two NAC domain transcription factors, SND1 and NST1, function redundantly in regulation of secondary wall synthesis in fibers of Arabidopsis. Planta 2007, 225, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Tusong, K.; Mamat, A.; Xu, J. Genome-wide investigation of NAC family genes potentially related to SCW biogenesis in korla pear fruit. Plant Mol. Biol. Rep. 2022. [Google Scholar] [CrossRef]
- Vargas-Hernández, B.Y.; Núñez-Muñoz, L.; Calderón-Pérez, B.; Xoconostle-Cázares, B.; Ruiz-Medrano, R. The NAC transcription factor ANAC087 induces aerial rosette development and leaf senescence in Arabidopsis. Front. Plant Sci. 2022, 13, 818107. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, J.Y.; Xu, P.; Zhang, R.; Li, L.G. Intron-mediated alternative splicing of wood-associated NAC transcription factor1B regulates cell wall thickening during fiber development in Populus species. Plant Physiol. 2014, 164, 765–776. [Google Scholar] [CrossRef] [Green Version]
- Zhong, R.Q.; Lee, C.; Haghighat, M.; Ye, Z.H. Xylem vessel-specific SND5 and its homologs regulate secondary wall biosynthesis through activating secondary wall NAC binding elements. New Phytol. 2021, 231, 1496–1509. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, D.W.; Zhou, B.; Wang, J.P.; Li, R.H.; Jiang, T.B. Over-expression of poplar NAC15 gene enhances wood formation in transgenic tobacco. BMC Plant Biol. 2020, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Wang, W.Q.; Zeng, J.K.; Zhang, J.; Grierson, D.; Li, X.; Yin, X.R.; Chen, K.S. A NAC transcription factor, EjNAC1, affects lignification of loquat fruit by regulating lignin. Postharvest Biol. Tec. 2015, 102, 25–31. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Luan, Y.T.; Shi, W.B.; Tang, Y.H.; Huang, X.Q.; Tao, J. Melatonin enhances stem strength by increasing lignin content and secondary cell wall thickness in herbaceous peony. J. Exp. Bot. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, X.K.; Zheng, T.C.; Li, S.Z.; Zhang, Z.Y.; Zhang, M.; Zhang, Y.C.; Ahmad, S.; Sun, L.D.; Wang, J.; Cheng, T.R.; et al. Identification of the PmWEEP locus controlling weeping traits in Prunus mume through an integrated genome-wide association study and quantitative trait locus mapping. Hort. Res. 2021, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.Q.; Feng, L.; Wu, J.Y.; Ye, Y.J.; Zheng, T.C.; Cai, M.; Cheng, T.R.; Wang, J.; Zhang, Q.X.; Pan, H.T. Transcriptome analysis of the genes regulating phytohormone and cellular patterning in Lagerstroemia plant architecture. Sci. Rep. 2018, 8, 15162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.X.; Tian, M. Investigation on crape myrtle varieties in China. Adv. Ornament Hortic. China 2008, 2008, 654–658. [Google Scholar]
- Zhu, Q.K.; Zou, J.X.; Zhu, M.L.; Liu, Z.B.; Feng, P.C.; Fan, G.T.; Wang, W.J.; Liao, H. In silico analysis on structure and DNA binding mode of AtNAC1, a NAC transcription factor from Arabidopsis thaliana. J. Mol. Model. 2014, 20, 2117. [Google Scholar] [CrossRef]
- Guo, T.T.; Mao, X.G.; Zhang, H.; Zhang, Y.; Fu, M.D.; Sun, Z.F.; Kuai, P.; Lou, Y.G.; Fang, Y.D. Lamin-like proteins negatively regulate plant immunity through NAC with transmembrane motif1-like 9 and nonexpresser of pr genes1 in Arabidopsis thaliana. Mol. Plant 2017, 10, 1334–1348. [Google Scholar] [CrossRef] [Green Version]
- Li, W.H.; Zeng, Y.L.; Yin, F.L.; Wei, R.; Mao, X.F. Genome-wide identification and comprehensive analysis of the NAC transcription factor family in sunflower during salt and drought stress. Sci. Rep. 2021, 11, 19865. [Google Scholar] [CrossRef]
- Wang, X.; Basnayake, B.M.; Zhang, H.; Li, G.; Li, W.; Virk, N.; Mengiste, T.; Song, F. The Arabidopsis ATAF1, a NAC transcription factor, is a negative regulator of defense responses against necrotrophic fungal and bacterial pathogens. Mol. Plant Microbe Interact. 2009, 22, 1227–1238. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.B.; Qi, G.; Kong, Y.Z.; Kong, D.J.; Gao, Q.; Zhou, G.K. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [Green Version]
- Fideghelli, C.; Sartori, A.; Grassi, F. Fruit tree size and architecture. Acta. Hort. 2003, 622, 279–293. [Google Scholar] [CrossRef]
- Weise, E.S.; Kiss, Z.J. Gravitropism of inflorescence stems in starch-deficient mutants of Arabidopsis. Int. J. Plant Sci. 1999, 160, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Hoson, T.; Wakabayashi, K. Role of the plant cell wall in gravity resistance. Phytochemistry 2015, 112, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q. Lignification: Flexibility, Biosynthesis and Regulation. Trends Plant Sci. 2016, 21, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, P.; Zheng, T.C.; Zhang, Q.X. Genomic insights into the important ornamental and stress resistance traits of Prunus mume. Sci. Hortic. 2022, 302, 111179. [Google Scholar] [CrossRef]
- Yang, S.D.; Seo, P.J.; Yoon, H.K.; Park, C.M. The Arabidopsis NAC transcription factor VNI2 integrates abscisic acid signals into leaf senescence via the COR/RD genes. Plant Cell 2011, 23, 2155–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated ortholog resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2018, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zheng, T.C.; Chen, Z.L.; Ju, Y.Q.; Zhang, H.; Cai, M.; Pan, H.T.; Zhang, Q.X. Reference gene selection for qRT-PCR analysis of flower development in Lagerstroemia indica and L. speciosa. PLoS ONE 2018, 13, e0195004. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcriptome Gene ID (Gene Name) | Amino Acid Number/aa | Molecular Weight/Da | Isoelectric Point | Percentage of the Amino Acids with Highest Content% | Aliphatic Index | Instability Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| TRINITY_DN1513_c0_g1_i2 (LiNAC1) | 566 | 61,920.54 | 4.6 | Ser(S)12.2% | 65.81 | 46.22 | −0.592 | Nucleus |
| TRINITY_DN1513_c0_g1_i4 (LiNAC2) | 433 | 48,711.53 | 4.49 | Ser(S)10.2% | 62.84 | 43.93 | −0.758 | Nucleus |
| TRINITY_DN1552_c0_g1_i1 (LiNAC3) | 373 | 41,299 | 4.58 | Ala(A)9.7% | 69.87 | 36.19 | −0.53 | Nucleus |
| TRINITY_DN1552_c0_g2_i1 (LiNAC4) | 406 | 44,609.42 | 4.42 | Gly(G)9.9% | 66.6 | 39.37 | −0.525 | Nucleus |
| TRINITY_DN17248_c0_g1_i1 (LiNAC5) | 286 | 32,581.02 | 8.41 | Leu(L)10.5% | 73.01 | 48.22 | −0.634 | Nucleus |
| TRINITY_DN17248_c0_g1_i5 (LiNAC6) | 288 | 32,682.93 | 8.67 | Leu(L)10.4% | 69.79 | 42.84 | −0.619 | Nucleus |
| TRINITY_DN1795_c0_g1_i3 (LiNAC7) | 668 | 74,336.43 | 5.15 | Ser(S)10.5% | 68.64 | 51.19 | −0.691 | Nucleus |
| TRINITY_DN2758_c0_g1_i5 (LiNAC8) | 414 | 47,164.68 | 5.39 | Leu(L)8.7% | 68.79 | 59.01 | −0.731 | Nucleus |
| TRINITY_DN419_c0_g1_i2 (LiNAC9) | 266 | 30,251.89 | 9.36 | Ser(S)10.9% | 57.89 | 52.67 | −0.85 | Nucleus |
| TRINITY_DN419_c0_g1_i4 (LiNAC10) | 264 | 29,841.31 | 8.9 | Ser(S)12.1% | 59.39 | 56.49 | −0.795 | Nucleus |
| TRINITY_DN419_c0_g1_i6 (LiNAC11) | 270 | 30,776.37 | 8.89 | Asn(N)& Ser (S) 9.3% | 60.63 | 47.02 | −0.905 | Nucleus |
| TRINITY_DN419_c0_g1_i8 (LiNAC12) | 270 | 30,781.24 | 8.89 | Ser(S)10.0% | 59.92 | 49.93 | −0.942 | Nucleus |
| TRINITY_DN4293_c0_g3_i1 (LiNAC13) | 341 | 37,460.37 | 6.89 | Ser(S)10.9% | 68.94 | 49.69 | −0.415 | Nucleus |
| TRINITY_DN894_c0_g1_i10 (LiNAC14) | 234 | 26,491.05 | 7.73 | Leu(L)11.1% | 75.47 | 29.93 | −0.671 | Nucleus |
| TRINITY_DN894_c0_g1_i11 (LiNAC15) | 328 | 37,132.65 | 4.83 | Asp(D)8.5% | 68.69 | 38.22 | −0.666 | Nucleus |
| TRINITY_DN894_c0_g1_i22 (LiNAC16) | 531 | 59,644.15 | 5.3 | Ser(S)8.7% | 71.62 | 48.59 | −0.63 | Nucleus |
| TRINITY_DN894_c0_g1_i31 (LiNAC17) | 311 | 34,893.12 | 4.92 | Pro(P)8.4% | 67.46 | 47.53 | −0.7 | Nucleus |
| TRINITY_DN894_c0_g1_i4 (LiNAC18) | 444 | 49,822.84 | 5.02 | Pro(P)8.1% | 69.19 | 47.22 | −0.703 | Nucleus |
| TRINITY_DN909_c0_g1_i1 (LiNAC19) | 245 | 27,455.47 | 9.93 | Arg(R)10.2% | 71.18 | 62.67 | −0.638 | Nucleus |
| TRINITY_DN909_c0_g1_i3 (LiNAC20) | 239 | 26,992.33 | 9.16 | Ser(S)11.7% | 63.18 | 58.10 | −0.743 | Nucleus |
| TRINITY_DN909_c0_g2_i1 (LiNAC21) | 255 | 28,368.91 | 9.3 | Ser(S)11.4% | 59.65 | 55.76 | −0.747 | Nucleus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, C.; Shang, L.; Zhang, G.; Wang, Q.; Ma, Q.; Hong, S.; Zhao, Y.; Yang, L. Identification and Expression Analysis of NAC Gene Family in Weeping Trait of Lagerstroemia indica. Plants 2022, 11, 2168. https://doi.org/10.3390/plants11162168
Gu C, Shang L, Zhang G, Wang Q, Ma Q, Hong S, Zhao Y, Yang L. Identification and Expression Analysis of NAC Gene Family in Weeping Trait of Lagerstroemia indica. Plants. 2022; 11(16):2168. https://doi.org/10.3390/plants11162168
Chicago/Turabian StyleGu, Cuihua, Linxue Shang, Guozhe Zhang, Qun Wang, Qingqing Ma, Sidan Hong, Yu Zhao, and Liyuan Yang. 2022. "Identification and Expression Analysis of NAC Gene Family in Weeping Trait of Lagerstroemia indica" Plants 11, no. 16: 2168. https://doi.org/10.3390/plants11162168