
Effects of Contaminated Soil on the Survival and Growth Performance of European (Populus tremula L.) and Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones Based on Stand Density

Abstract
:1. Introduction
2. Materials and methods
2.1. Soil Materials
2.2. Plant Materials
2.3. Experimental Design
2.4. Statistical Analysis
3. Results
3.1. Survival Scenario
3.2. Absolute Height
3.3. Diameter
3.4. Dry Biomass of Stem
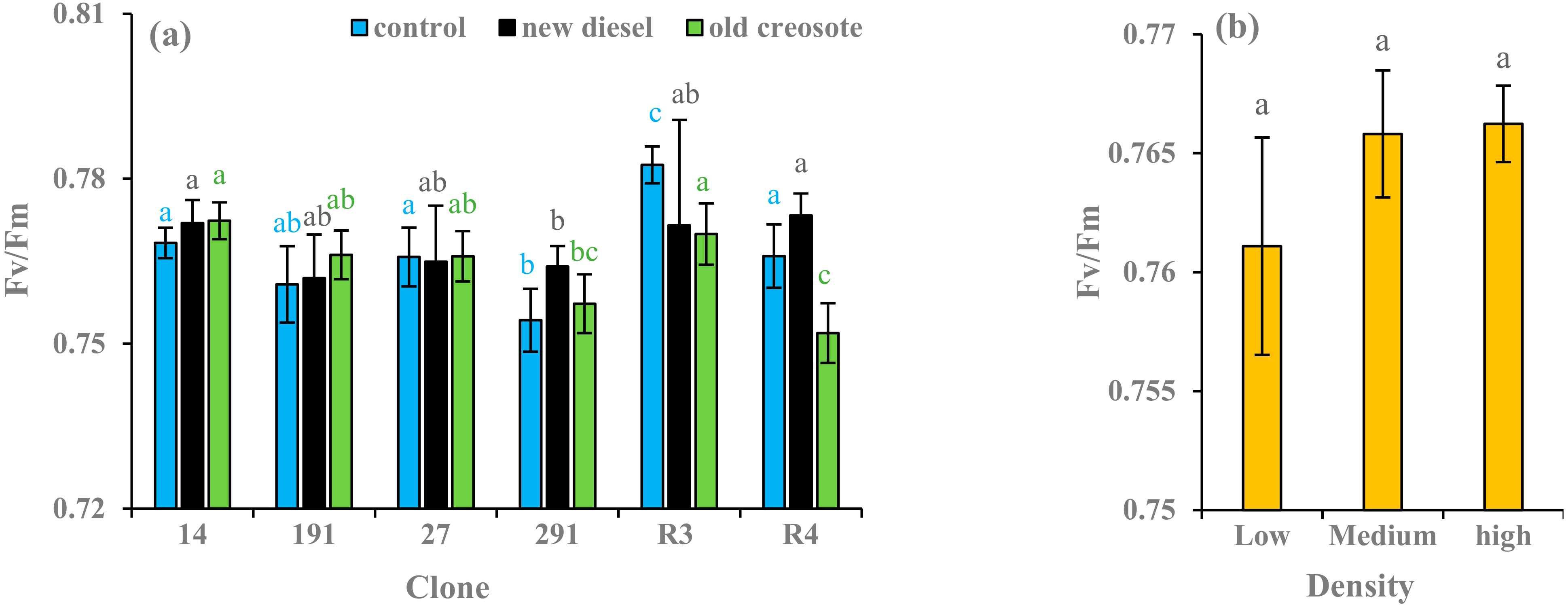
3.5. Chlorophyll Fluorescence (Fv/Fm)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cook, R.L.; Hesterberg, D. Comparison of trees and grasses for rhizoremediation of petroleum hydrocarbons. Int. J. Phytorem. 2013, 15, 844–860. [Google Scholar] [CrossRef]
- Plohl, K.; Leskovsek, H.; Bricelj, M. Biological degradation of motor oil in water. Acta Chim. Slov. 2002, 49, 279–289. [Google Scholar]
- Wu, B.; Zhang, Y.; Zhang, X.X.; Cheng, S.P. Health risk assessment of polycyclic aromatic hydrocarbons in the source water and drinking water of China: Quantitative analysis based on published monitoring data. Sci. Total Environ. 2012, 410, 112–118. [Google Scholar] [CrossRef]
- Van Liedekerke, M.; Prokop, G.; Rabl-Berger, S.; Kibblewhite, M.; Louwagie, G. Progress in the Management of Contaminated Soils in Europe. Joint Research Center. Institute for Environment and Sustainability. 2014. Available online: http://eusoils.;jrc.;ec.;europa.;eu/ESDB_Archive/eusoils_docs/other/EUR26376EN.;pdf (accessed on 1 January 2022).
- Agnello, A.C.; Bagard, M.; Van Hullebusch, E.D.; Esposito, G.; Huguenot, D. Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci. Total Environ. 2016, 563, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Paccassoni, F.; Kalnina, D.; Piga, L. Comparative studies of oil product regulation in contaminated soil for several industrialized countries. Mater. Sci. Eng. 2017, 251, 012066. [Google Scholar]
- Camille, D.H. The Use of Aspens for Phytoremediation: Effects of Contamination by Petroleum Hydrocarbons, Planting Density, and Clone Type on the Establishment of European Aspens (Populus tremula L.) and Hybrid Aspens (Populus tremula L. × Populus tremuloides Michx.). Master’s Thesis, Agriculture, Environment et Territoire, Vetagro Sup.,, Lempdes, France, 2015. [Google Scholar]
- Zalensy, R.S., Jr.; Bauer, E.O.; Richard, R.B.; Zalensy, J.A.; Kunzman, J.; Rog, C.J.; Riemenschneider, D.E. Clonal variation in survival and growth of hybrid poplar and Salix in an in–situ trial on soils heavily contaminated with petroleum hydrocarbons. Int. J. Phytorem. 2005, 7, 177–197. [Google Scholar]
- Sharma, H.D.; Reddy, K.R. Geoenvironmental Engineering: Site Remediation, Waste Containment, and Emerging Waste Management Technologies; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2004. [Google Scholar]
- Fasani, E.; Manara, A.; Martini, F.; Furini, A.; DalCorso, G. The potential of genetic engineering of plants for the remediation of soils contaminated with heavy metals. Plant Cell Environ. 2018, 41, 1201–1232. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E. Phytoremediation: Annual review. Plant Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef] [PubMed]
- Greger, M. Influence of willow (Salix viminalis L.) roots on soil metal chemistry: Effects of clones with varying metal uptake potential. In Biogeochemistry of Trace Elements in the Rhizosphere; Huang, P.M., Gobran, G.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 301–312. [Google Scholar]
- Salam, M.M.A.; Kaipiainen, E.; Mohsin, M.; Villa, A.; Kuittinen, S.; Pulkkinen, P.; Pelkonen, P.; Mehtätalo, L.; Pappinen, A. Effects of Contaminated Soil on the Growth Performance of Young Salix (Salix Schwerinii E. L. Wolf) and the Potential for Phytoremediation of Heavy Metals. J. Environ. Manag. 2016, 183, 467–477. [Google Scholar] [CrossRef]
- Sivaram, A.K.; Logeshwaran, P.; Subashchandrabose, S.R.; Lockington, R.; Naidu, R.; Megharaj, M. Comparison of plants with C3 and C4 carbon fixation pathways for remediation of polycyclic aromatic hydrocarbon contaminated soils. Sci. Rep. 2018, 8, 2100. [Google Scholar] [CrossRef]
- Mohsin, M.; Kuittinen, S.; Salam, M.M.A.; Peräniemi, S.; Laine, S.; Pulkkinen, P.; Kaipiainen, E.; Vepsäläinen, J.; Pappinen, A. Chelate-assisted phytoextraction: Growth and ecophysiological responses by Salix schwerinii EL Wolf grown in artificially polluted soils. J. Geochem. Explor. 2019, 205, 106335. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J.; Salam, M.M.A.; Ma, C.; Chen, G. Impacts of bamboo biochar on the phytoremediation potential of Salix psammophila grown in multi–metals contaminated paddy soil. Int. J. Phytoremed. 2020, 23, 387–399. [Google Scholar] [CrossRef]
- Salam, M.M.A.; Mohsin, M.; Pulkkinen, P.; Pelkonen, P.; Pappinen, A. Effects of Soil Amendments on the Growth Response and Phytoextraction Capability of a Willow Variety (S. viminalis × S. schwerinii × S. dasyclados) Grown in Contaminated Soils. Ecotoxicol. Environ. Safe 2019, 171, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Pilipović, A.; Zalesny, R.S., Jr.; Rončević, S.; Nikolić, N.; Orlović, S.; Beljin, J.; Katanić, M. Growth, physiology, and phytoextraction potential of poplar and willow established in soils amended with heavy metal contaminated, dredged river sediments. J. Environ. Manag. 2019, 239, 352–365. [Google Scholar] [CrossRef]
- Iori, V.; Zacchini, M.; Pietrini, F. Growth, physiological response and phytoremoval capability of two willow clones exposed to ibuprofen under hydroponic culture. J. Hazard. Mater. 2013, 262, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, A.; Salvatori, E.; Di Re, S.; Fusaro, L.; Nervo, G.; Manes, F. Natural and commercial Salix clones differ in their ecophysiological response to Zn stress. Photosynthetica 2016, 54, 56–64. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Pietrini, F.; Zacchini, M.; Iori, V.; Pietrosanti, L.; Ferretti, M.; Massacci, A. Spatial distribution of cadmium in leaves and its impact on photosynthesis: Examples of different strategies in Salix and poplar varieties. Plant Biol. 2010, 12, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Yu, Q. Can physiological and anatomical characters be used for selecting high yielding hybrid aspen clones? Silva Fenn. 2001, 35, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Pulkkinen, P. Genotype–environment interaction and stability in growth of aspen hybrid clones. For. Ecol. Manag. 2003, 173, 25–35. [Google Scholar] [CrossRef]
- Zacchini, M.; Iori, V.; Mugnozza, G.S.; Pietrini, F.; Massacci, A. Cadmium accumulation and tolerance in Populus nigra and Salix Alba. Biol. Plant. 2011, 55, 383–386. [Google Scholar] [CrossRef]
- Korhonen, K.T.; Ihalainen, A.; Viiri, H.; Heikkinen, J.; Henttonen, H.M.; Hotanen, J.-P.; Mäkelä, H.; Nevalainen, S.; Pitkänen, J. Suomen metsät 2004–2008 ja niiden kehitys 1921–2008. Metsätiueteen Aikakauskirja 2013, 3, 269–333. (In Finnish) [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Beuker, E.; Viherä-Aarnio, A.; Hynynen, J. Site index models with density effect for hybrid aspen (Populus tremula L. × P. tremuloides Michx.) plantations in southern Finland. For. Ecol. Manag. 2021, 480, 118669. [Google Scholar] [CrossRef]
- Hynynen, J.; Ahtikoski, A.; Eskelinen, T. Viljelyhaavikon tuotos ja kasvatuksen kannattavuus. Metsätieteen Aikakauskirja 2004, 1, 113–116. (In Finnish) [Google Scholar] [CrossRef]
- Guerra, F.; Gainza, F.; Pérez, R.; Zamudio., F. Phytoremediation of heavy metals using poplars (Populus spp.): A glimpse of the plant responses to copper, cadmium and zinc stress. In Handbook of Phytoremediation; Nova Science: New York, NY, USA, 2011; Ivan, A., Golubev, Eds.; Nova Science: New York, NY, USA, 2011; pp. 387–413. [Google Scholar]
- Aprill, W.; Sims, R.C. Evaluation of the use of prairie grasses for stimulating polycyclic aromatic hydrocarbon treatment in soil. Chemosphere 1990, 20, 253–265. [Google Scholar] [CrossRef]
- Hynynen, J.; Karlsson, K. Intensive management of hybrid aspen in Finland. Management and utilization of broadleaved tree species in Nordic and Baltic countries—Birch, aspen and alder. In Proceedings of the Workshop, Vantaa, Finland, 16–18 May 2001. [Google Scholar]
- Rytter, L. A management regime for hybrid aspen stands combining conventional forestry techniques with early biomass harvests to exploit their rapid early growth. For. Ecol. Manag. 2006, 236, 422–426. [Google Scholar] [CrossRef]
- Salt, D.E.; Blaylock, M.; Kumar, N.P.B.A.; Dushenkov, V.; Ensley, B.D.; Chet, I.; Raskin, I. Phytoremediation: A Novel Strategy for the Removal of Toxic Metals from the Environment Using Plants. Nat. Biotech. 1995, 13, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Shrive, S.C.; McBride, R.A.; Gordon, A.M. Photosynthetic and Growth Responses of Two Broad-Leaf Tree Species to Irrigation with Municipal Landfill Leachate. J. Environ. Qual. 1994, 23, 534–542. [Google Scholar] [CrossRef]
- Justin, M.Z.; Pajk, N.; Zupanc, V.; Zupančič, M. Phytoremediation of landfill leachate and compost wastewater by irrigation of Populus and Salix: Biomass and growth response. Waste Manag. 2010, 30, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Salam, M.M.A.; Mohsin, M.; Rasheed, F.; Ramzan, M.; Zafar, Z.; Pulkkinen, P. Assessment of European and hybrid aspen clones efficiency based on height growth and removal percentage of petroleum hydrocarbons—A field trial. Environ. Sci. Pollut. Res. 2020, 27, 45555–45567. [Google Scholar] [CrossRef] [PubMed]
- Marmiroli, M.; Pietrini, F.; Maestri, E.; Zacchini, M.; Marmiroli, N.; Massacci, A. Growth, physiological and molecular traits in Salicaceae trees investigated for phytoremediation of heavy metals and organics. Tree Physiol. 2011, 31, 1319–1334. [Google Scholar] [CrossRef] [Green Version]
- Salam, M.M.A.; Mohsin, M.; Kaipiainen, E.; Villa, A.; Kuittinen, S.; Pulkkinen, P.; Pelkonen, P.; Pappinen, A. Biomass growth variation and phytoextraction potential of four Salix varieties grown in contaminated soil amended with lime and wood ash. Int. J. Phytoremed. 2019, 21, 1329–1340. [Google Scholar] [CrossRef]
- Van Dillewijn, P.; Couselo, J.L.; Corredoira, E.; Delgado, A.; Wittich, R.M.; Ballester, A.; Ramos, J.L. Bioremediation of 2, 4, 6-trinitrotoluene by bacterial nitroreductase expressing transgenic aspen. Environ. Sci. Technol. 2008, 42, 7405–7410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangoi, L. Understanding plant density effects on maize growth and development: An important issue to maximize grain yield. Cienc. Rural 2001, 31, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Søndergaard, M. Seasonal variations in the loosely sorbed phosphorus fraction of the sediment of a shallow and hypertrophic lake. Environ. Geol. Water Sci. 1988, 11, 115–121. [Google Scholar] [CrossRef]
- Li, Y.M.; Chaney, R.; Brewer, E.; Roseberg, R.; Angle, J.S.; Baker, A.; Reeves, R.; Nelkin, J. Development of a technology for commercial phytoextraction of nickel: Economic and technical considerations. Plant and Soil 2003, 249, 107–115. [Google Scholar] [CrossRef]
- Chaney, R.L.; Angle, J.S.; Broadhurst, C.L.; Peters, C.A.; Tappero, R.V.; Sparks, D.L. Improved understanding of hyperaccumulation yields commercial phytoextraction and phytomining technologies. J. Environ. Qual. 2007, 36, 1429–1443. [Google Scholar] [CrossRef] [Green Version]
- Chaney, R.L.; Li, Y.M.; Angle, J.S.; Baker, A.J.M.; Reeves, R.D.; Brown, S.L.; Homer, F.A.; Malik, M.; Chin, M. Improving metal hyperaccumulator wild plants to develop commercial phytoextraction systems: Approaches and progress. In Phytoremediation of Contaminated Soil and Water; Terry, N., Bañuelos, G.S., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 131–160. [Google Scholar]
- Sinkkonen, A. Density-dependent chemical interference—an extension of the biological response model. J. Chem. Ecol. 2001, 27, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- San Emeterio, L.; Damgaard, C.; Cannals, R.M. Modelling the combined effect of chemical interference and resource competition on the individual growth of two herbaceous populations. Plant Soil 2007, 292, 95–103. [Google Scholar] [CrossRef]
- Hansi, M.; Weidenhamer, J.; Sinkkonen, A. Plant growth responses to inorganic environmental contaminants are density-dependent: Experiments with copper sulfate, barley and lettuce. Environ. Pollut. 2014, 184, 443–448. [Google Scholar] [CrossRef]
- Xu, M.; Natural Resources Institute, Finland. Phytoremediation with aspen trees in Luumäki. Unpublished Manuscript. 2013. [Google Scholar]
- Mukherjee, S.; Juottonen, H.; Siivonen, P.; Lloret Quesada, C.; Tuomi, P.; Pulkkinen, P.; Yrjälä, K. Spatial patterns of microbial diversity and activity in an aged creosote–contaminated site. ISME J. 2014, 98, 2131–2142. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Mäntylä, N.; Salonen, M. Rooting of Hybrid Clones of Populus tremula L. × P. tremuloides Michx. by Stem Cuttings Derived from Micropropagated Plants. Scand. J. For. Res. 2001, 16, 238–245. [Google Scholar] [CrossRef]
- Evlard, A.; Sergeant, K.; Ferrandis, S.; Printz, B.; Renaut, J.; Guignard, C.; Paul, R.; Hausman, J.F.; Campanella, B. Physiological and Proteomic Responses of Different Salix Varieties (Salix fragilis × alba) Exposed to Dredged Sediment Contaminated by HMs. Int. J. Phytorem. 2014, 16, 1148–1169. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. A Language and Environment for Statistical Computing, Vienna (Austria): R Foundation for Statistical Computing. 2021. Available online: https://www.;r-project.;org/ (accessed on 7 May 2021).
- Anyasi, R.O.; Atagana, H.I. Profiling of plants at petroleum contaminated site for phytoremediation. Int. J. Phytoremed. 2018, 20, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Close, D.; Beadle, C.; Brown, P. The physiological basis of containerised tree seedling “transplant shock”: A review. Aust. For. 2004, 68, 112–120. [Google Scholar] [CrossRef]
- Baker, J. The effect of oil on plants. Environ. Pollut. 1970, 1, 27–44. [Google Scholar] [CrossRef]
- Tang, J.; Lu, X.; Sun, Q.; Zhu, W. Aging effect of petroleum hydrocarbons in soil under different attenuation conditions. Agric. Ecosyst. Environ. 2011, 149, 109–117. [Google Scholar] [CrossRef]
- Qin, Y.; Shi, X.; Wang, Z.; Pei, C.; Cao, M.; Luo, J. Influence of Planting Density on the Phytoremediation Efficiency of Festuca arundinacea in cd-Polluted Soil. Bull. Environ. Contam. Toxicol. 2021, 107, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Garnica, E.A. Adaptability and Development of Populus tremula L. × Populus tremuloides Michx. and Finnish Native Populus Tremula on Polluted Soils by PAHs and Sodium Chloride. Master’s Thesis, Novia University of Applied Sciences, Vaasa, Finland, 2014. [Google Scholar]
- Haider, F.U.; Ejaz, M.; Cheema, S.A.; Khan, M.I.; Zhao, B.; Liqun, C.; Salim, M.A.; Naveed, M.; Khan, N.; Núñez-Delgado, A.; et al. Phytotoxicity of petroleum hydrocarbons: Sources, impacts and remediation strategies. Environ. Res. 2021, 197, 111031. [Google Scholar] [CrossRef]
- Nguemté, P.M.; Wafo, G.D.; Djocgoue, P.F.; Noumsi, I.K.; Ngnien, A.W. Potentialities of Six Plant Species on Phytoremediation Attempts of Fuel Oil–Contaminated Soils. Water Air Soil Pollut. 2018, 229, 1–18. [Google Scholar]
- Shirdam, R.; Zand, A.; Bidhendi, G.; Mehrdadi, N. Phytoremediation of hydrocarbon–contaminated soils with emphasis on the effect of petroleum hydrocarbons on the growth of plant species. Phytoprotection 2008, 89, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Sun, T.; Song, Y.; Ackland, M.L.; Liu, Y. Strategies for enhancing the phytoremediation of cadmium-contaminated agricultural soils by Solanum nigrum L. Environ. Pollut. 2011, 159, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.H.; An, S.Q.; Wu, B.F. Density-dependent root morphology and root distribution in the submerged plant Vallisneria natans. Environ. Exp. Bot. 2006, 57, 195–200. [Google Scholar] [CrossRef]
- Jiang, J.H.; Zhou, C.F.; An, S.Q. Sediment type, population density and their combined effect greatly charge the short-time growth of two common submerged macrophytes. Ecol. Eng. 2008, 34, 79–90. [Google Scholar] [CrossRef]
- Liu, H.; Meng, F.; Tong, Y.; Chi, J. Effect of plant density on phytoremediation of polycyclic aromatic hydrocarbons contaminated sediments with Vallisneria spiralis. Ecol. Eng. 2014, 73, 380–385. [Google Scholar] [CrossRef]
- Xie, Y.; Yu, D. The significance of lateral roots in phosphorus (P) acquisition of water hyacinth (Eichhornia crassipes). Aquat. Bot. 2003, 75, 311–321. [Google Scholar] [CrossRef]
- Sinkkonen, A. A model describing chemical interference caused by decomposing residues at different densities of growing plants. Plant Soil 2003, 250, 315–322. [Google Scholar] [CrossRef]
- Sinkkonen, A. Modelling the effect of autotoxicity on density-dependent phytotoxicity. J. Theor. Biol. 2007, 244, 218–227. [Google Scholar] [CrossRef]
- Chu, C.J.; Maestre, F.T.; Xiao, S.; Weiner, J.; Wang, Y.S.; Duan, Z.H.; Wang, G. Balance between facilitation and resource competition determines biomass-density relationships in plant populations. Ecol. Lett. 2008, 11, 1189–1197. [Google Scholar] [CrossRef]
- Chu, C.J.; Weiner, J.; Maestre, F.T.; Xiao, S.; Wang, Y.S.; Li, Q.; Yuan, J.L.; Zhao, L.Q.; Ren, Z.W.; Wang, G. Positive interactions can increase size inequality in plant populations. J. Ecol. 2009, 97, 1401–1407. [Google Scholar] [CrossRef]
- Sinkkonen, A.; Strömmer, R.; Penttinen, O.-P. Low toxicant concentrations decrease the frequency of fast-growing seedlings at high densities of annual baby’s breath (Gypsophila elegans). Environ. Pollut. 2008, 153, 523–525. [Google Scholar] [CrossRef] [PubMed]
- Sinkkonen, A.; Penttinen, O.-P.; Strömmer, R. Testing the homogenizing effect of low copper sulfate concentrations on the size distribution of Portulaca oleracea seedlings in vitro. Sci. Total Environ. 2009, 407, 4461–4464. [Google Scholar] [CrossRef] [PubMed]
- Sinkkonen, A.; Myyrä, M.; Penttinen, O.P.; Rantalainen, A.L. Selective toxicity at low doses: Experiments with three plant species and toxicants. Dose-Response 2011, 9, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Belz, R.G.; Sinkkonen, A. Selective toxin effects on faster and slower growing individuals in the formation of hormesis at the population level—A case study with Lactuca sativa and PCIB. Sci. Total Environ. 2016, 566, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.N.; Misra, M.; Sing, R. Chlorophyll Fluorescence in Plant Biology. In Biophysics; Misra, A.N., Ed.; IntechOpen: Shanghai, China, 2012; pp. 171–192. [Google Scholar]
- Zacchini, M.; Pietrini, F.; Mugnozza, G.S. Metal tolerance, accumulation and translocation in poplar and Salix clones treated with cadmium in hydroponics. Water Air Soil Pollut. 2009, 197, 23–34. [Google Scholar] [CrossRef]
- Nikolić, N.P.; Borišev, M.K.; Pajević, S.P.; Arsenov, D.D.; Župunski, M.D.; Orlović, S.S.; Pilipović, A.R. Photosynthetic response and tolerance of three willow species to cadmium exposure in hydroponic culture. Arch. Biol. Sci. 2015, 67, 1411–1420. [Google Scholar] [CrossRef]
- Stener, L.G.; Karlsson, B. Improvement of Populus tremula × P. tremuloides by phenotypic selection and clonal testing. For. Genet. 2004, 11, 13–27. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties of Soil | Unit | Soil Layer | References | |||||
|---|---|---|---|---|---|---|---|---|
| 0–10 cm | 5–10 cm | 10–50 cm | ||||||
| Minimum | Maximum | Mean | SE | |||||
| n = 4 | ||||||||
| pH | 5 a | 7.8 a | 5.94 | 0.31 | 6.1 b | 6.1 b | [37,49,50] | |
| n = 55 | ||||||||
| Total PAHs | ppm | 0.16 | 714 | 96.51 | 24.1 | 532 b | 164 b | |
| Naphthalene | 0.01 | 2.19 | 0.15 | 0.04 | ||||
| Acenaphthylene | 0.01 | 3.10 | 0.37 | 0.08 | ||||
| Acenaphthene | 0.01 | 7 | 0.23 | 0.13 | ||||
| Fluorene | 0.01 | 11 | 0.46 | 0.23 | ||||
| Phenanthrene | 0.01 | 22 | 0.87 | 0.42 | ||||
| Anthracene | 0.01 | 160 | 7.13 | 3.87 | ||||
| Fluoranthene | 0.01 | 213 | 21.89 | 6.09 | ||||
| Pyrene | 0.01 | 224 | 24.25 | 6.50 | ||||
| Benz(a)anthracene | 0.01 | 376 | 13.4 | 6.93 | ||||
| Chrysene | 0.01 | 100 | 11.35 | 2.86 | ||||
| Benzo(b)fluoranthene | 0.01 | 47.4 | 7.15 | 1.5 | ||||
| Benzo(k)fluoranthene | 0.01 | 22.9 | 4.12 | 0.87 | ||||
| Benzo[a]pyrene | 0.01 | 21 | 3.04 | 0.65 | ||||
| indeno(1,2,3-cd)pyrene | 0.01 | 5.62 | 0.98 | 0.18 | ||||
| Benzo[ghi]perylene | 0.01 | 5.61 | 0.67 | 0.14 | ||||
| Dibenz[a.h]anthracene | 0.01 | 2.26 | 0.44 | 0.08 | ||||
| C10–C21 | 10 | 580 | 99.84 | 20.44 | 556 b | 190 b | ||
| C22–C40 | 10 | 1780 | 338.8 | 67.06 | 2120 b | 688 b | ||
| C10–C40 | 20 | 2350 | 437.1 | 85.79 | 2675 b | 875.5 b | ||
| n = 30 | ||||||||
| C | % | 0.05 | 1.46 | 0.41 | 0.08 | |||
| N | 0.01 | 0.06 | 0.03 | 0.00 | ||||
| Ca | (mg kg–1) | 849 | 3020 | 1272 | 81.84 | |||
| Cu | 2.91 | 20.7 | 6.71 | 0.98 | ||||
| Fe | 4490 | 6950 | 5204 | 128.7 | ||||
| K | 393 | 690 | 510.6 | 13.37 | ||||
| Mg | 513 | 924 | 613.4 | 21.45 | ||||
| Ni | 1.25 | 5.69 | 2.10 | 0.21 | ||||
| P | 149 | 326 | 207.6 | 7.35 | ||||
| Pb | 2.81 | 15.4 | 5.07 | 0.56 | ||||
| Zn | 13.8 | 31.2 | 19.30 | 0.92 | ||||
| coarse sand | % | 2.03 | 28.74 | 10.53 | 1.29 | |||
| medium sand | 28.85 | 59.41 | 42.53 | 1.37 | ||||
| very fine sand | 18.26 | 52.55 | 40.06 | 1.66 | ||||
| Slit | 1.01 | 13.86 | 6.15 | 0.69 | ||||
| clay | 0.31 | 1.16 | 0.73 | 0.05 | ||||
| Variable | Factors | χ2 | DF (Degree of Freedom) | p-Value | Model |
|---|---|---|---|---|---|
| survival rate | clone | l2.3 | 5 | 0.0312 | 1 |
| soil treatment | 93.2 | 2 | <0.001 | ||
| density | 7.2 | 2 | 0.0272 | ||
| absolute height | clone | 106.99 | 5 | <0.001 | 2 |
| soil treatment | 15.05 | 2 | <0.001 | ||
| density | 35.65 | 2 | <0.001 | ||
| diameter | 423.22 | 1 | <0.001 | ||
| diameter | clone | 250.68 | 5 | <0.001 | 3 |
| soil treatment | 20.88 | 2 | <0.001 | ||
| density | 1.43 | 2 | 0.4893 | ||
| height | 425.7 | 1 | <0.001 | ||
| stem dry biomass | clone | 4.660 | 5 | 0.4587 | 4 |
| soil treatment | 47.886 | 2 | <0.001 | ||
| density | 52.850 | 2 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salam, M.M.A.; Ruhui, W.; Sinkkonen, A.; Pappinen, A.; Pulkkinen, P. Effects of Contaminated Soil on the Survival and Growth Performance of European (Populus tremula L.) and Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones Based on Stand Density. Plants 2022, 11, 1970. https://doi.org/10.3390/plants11151970
Salam MMA, Ruhui W, Sinkkonen A, Pappinen A, Pulkkinen P. Effects of Contaminated Soil on the Survival and Growth Performance of European (Populus tremula L.) and Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones Based on Stand Density. Plants. 2022; 11(15):1970. https://doi.org/10.3390/plants11151970
Chicago/Turabian StyleSalam, Mir Md Abdus, Wen Ruhui, Aki Sinkkonen, Ari Pappinen, and Pertti Pulkkinen. 2022. "Effects of Contaminated Soil on the Survival and Growth Performance of European (Populus tremula L.) and Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones Based on Stand Density" Plants 11, no. 15: 1970. https://doi.org/10.3390/plants11151970