
The Modified Brière Equation and Its Applications



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Methods
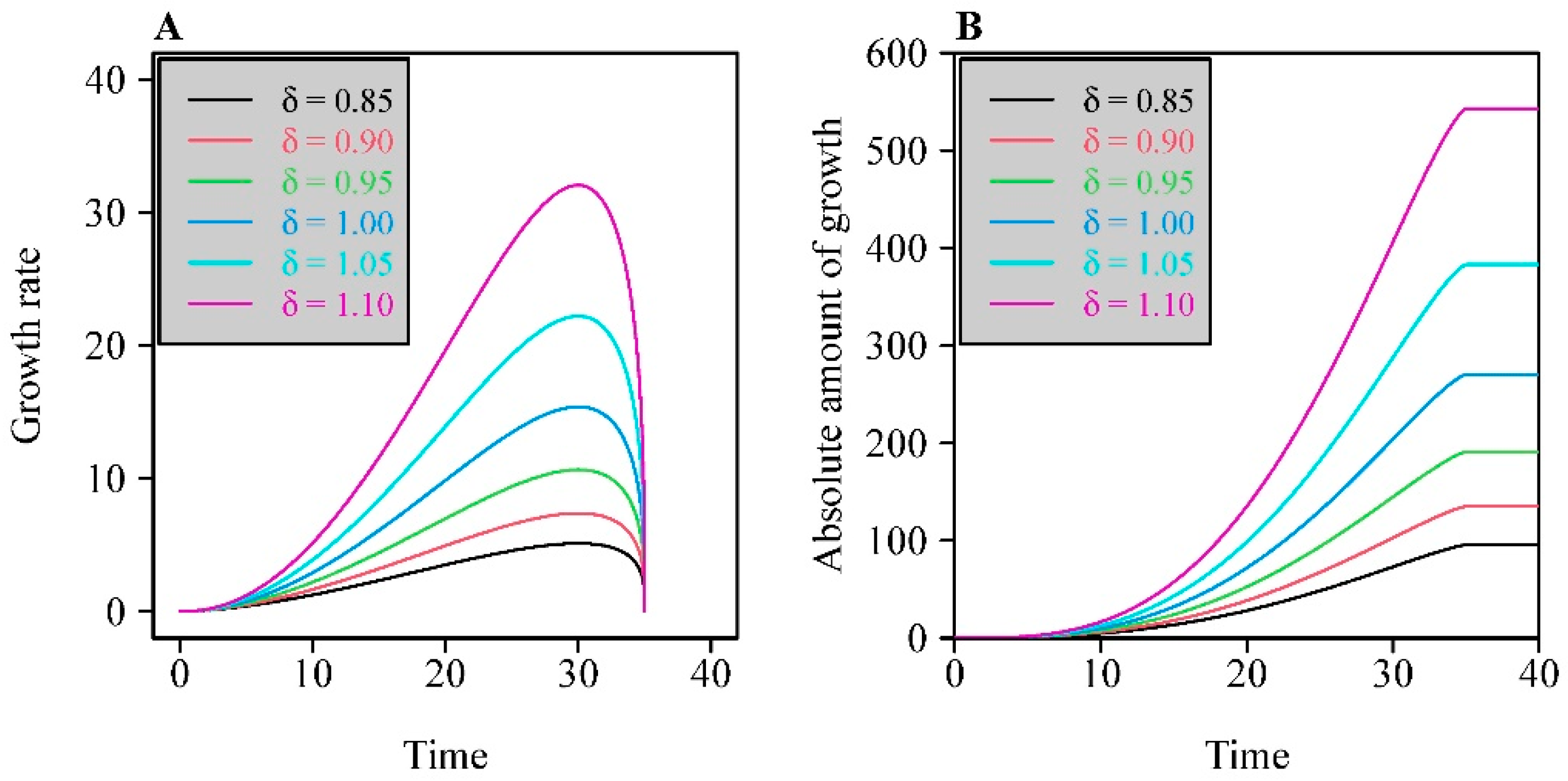
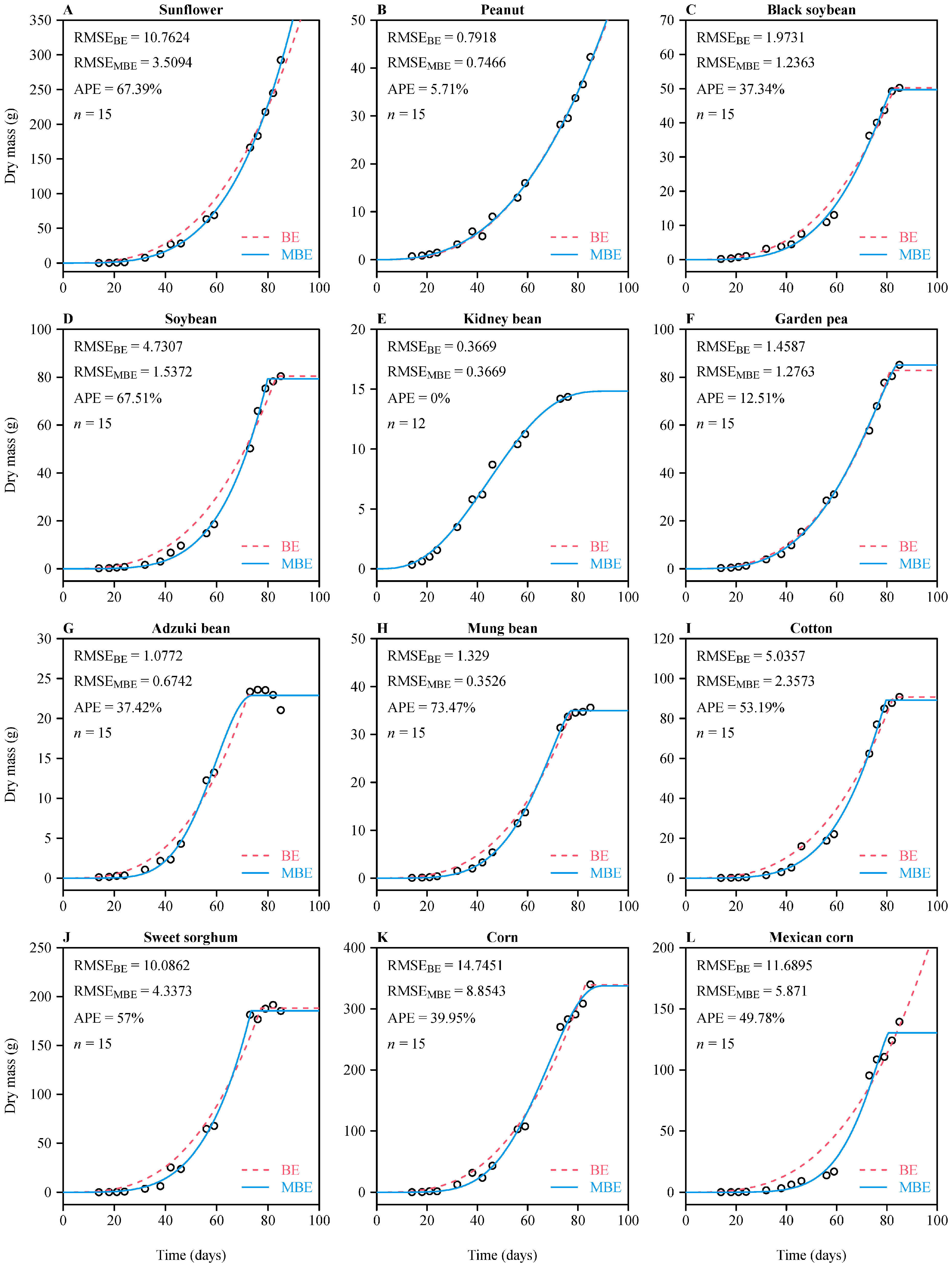
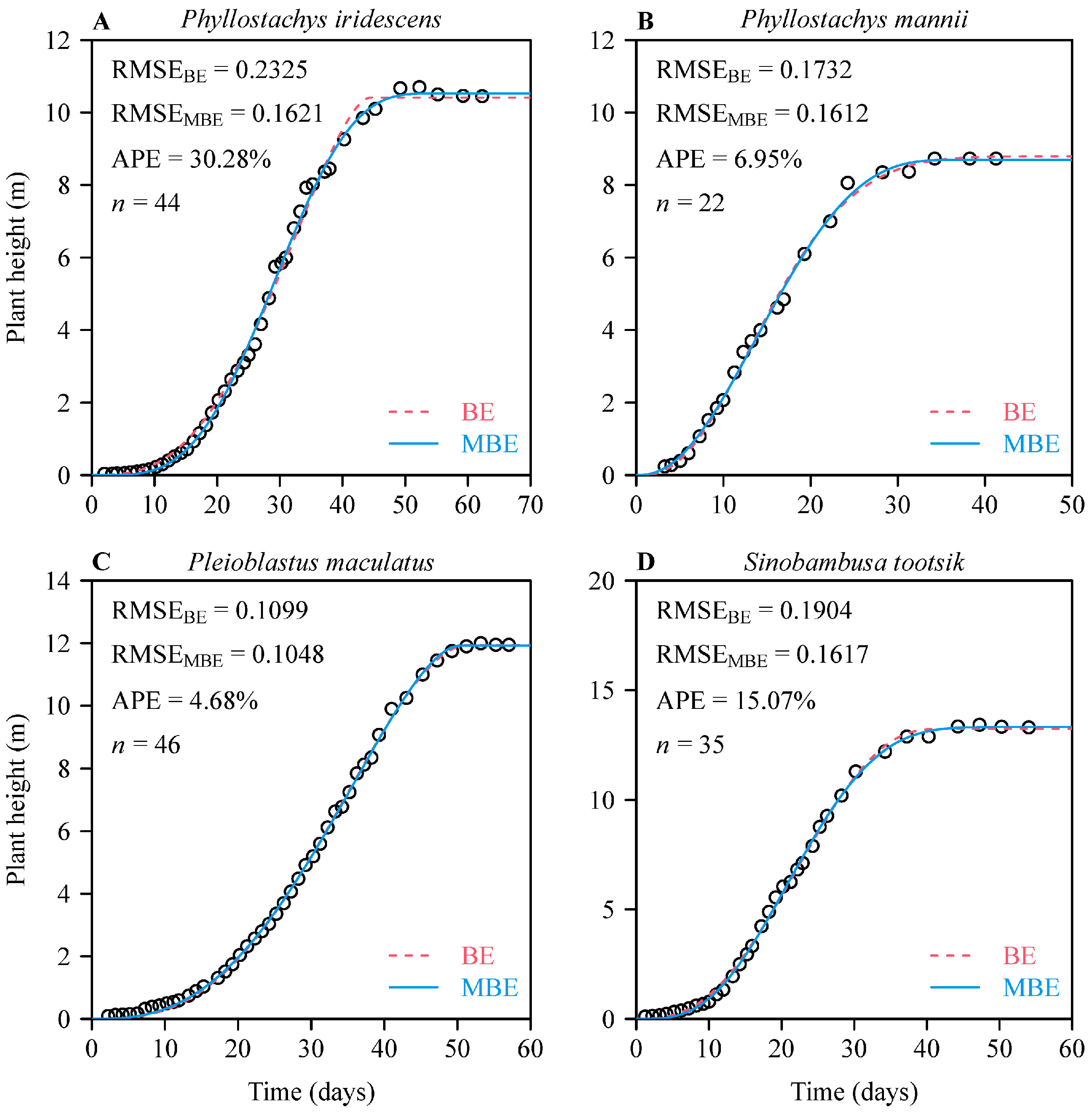
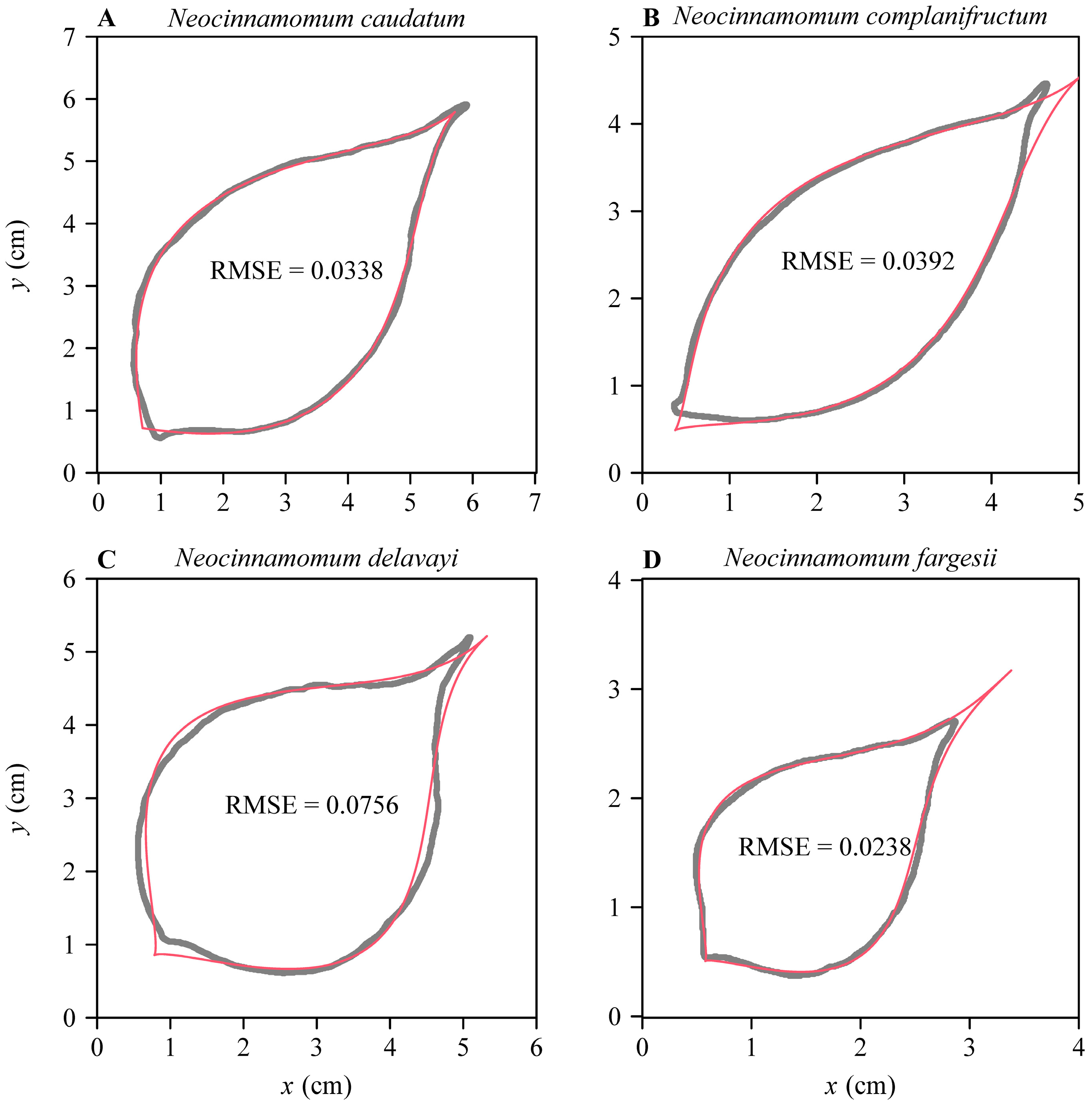
3. Results
4. Discussion
4.1. Elasticity in Curve Fitting of the Two Sigmoid Equations
4.2. Why We Did Not Compare the Two Sigmoid Equations with Other Equations
4.3. Reliability of Estimated Parameters in the Two Sigmoid Equations
4.4. Other Potential Applications of the Modified Brière Equation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Bertalanffy, L. Quantitative laws in metabolism and growth. Q. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef] [PubMed]
- West, G.B.; Brown, J.H.; Enquist, B.J. A general model for ontogenetic growth. Nature 2001, 413, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Goudriaan, J.; Lantinga, E.A.; Vos, J.; Spiertz, H.J. A flexible sigmoid function of determinate growth. Ann. Bot. 2003, 91, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, A.M.; Gorshkov, V.G.; Li, B.-L. Ontogenetic growth: Models and theory. Ecol. Model. 2004, 176, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Men, X.; Sandhu, H.S.; Chakraborty, A.; Li, B.; Ouyang, F.; Sun, Y.; Ge, F. The “general” ontogenetic growth model is inapplicable to crop growth. Ecol. Model. 2013, 266, 1–9. [Google Scholar] [CrossRef]
- Shi, P.; Ishikawa, T.; Sandhu, H.S.; Hui, C.; Chakraborty, A.; Jin, X.; Tachihara, K.; Li, B. On the 3/4-exponent von Bertalanffy equation for ontogenetic growth. Ecol. Model. 2014, 276, 23–28. [Google Scholar] [CrossRef]
- Verhulst, P.-F. Recherches mathématiques sur la loi d’accroissement de la population. Nouv. Mémoires L’académie R. Sci. Belles-Lett. Brux. 1845, 18, 14–54. [Google Scholar]
- Thornley, J.H.M.; Shepherd, J.J.; France, J. An open-ended logistic-based growth function: Analytical solutions and the power-law logistic model. Ecol. Model. 2007, 204, 531–534. [Google Scholar] [CrossRef]
- Shi, P.; Fan, M.; Ratkowsky, D.A.; Huang, J.; Wu, H.; Chen, L.; Fang, S.; Zhang, C. Comparison of two ontogenetic growth equations for animals and plants. Ecol. Model. 2017, 349, 1–10. [Google Scholar] [CrossRef]
- Paine, C.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: An update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Uvarov, B.P. Insects and climate. Trans. Entomol. Soc. 1931, 79, 1–232. [Google Scholar] [CrossRef]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Shi, P.; Reddy, G.V.P.; Chen, L.; Ge, F. Comparison of thermal performance equations in describing temperature-dependent developmental rates of insects: (I) empirical models. Ann. Entomol. Soc. Am. 2016, 109, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Ratkowsky, D.A.; Reddy, G.V.P. Empirical model with excellent statistical properties for describing temperature-dependent developmental rates of insects and mites. Ann. Entomol. Soc. Am. 2017, 110, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Briére, J.-F.; Pracros, P.; Le Roux, A.-Y.; Pierre, J.-S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Voorend, W.; Lootens, P.; Nelissen, H.; Roldán-Ruiz, I.; Inzé, D.; Muylle, H. LEAF-E: A tool to analyze grass leaf growth using function fitting. Plant Methods 2014, 10, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, P.; Chen, L.; Hui, C.; Grissino-Mayer, H.D. Capture the time when plants reach their maximum body size by using the beta sigmoid growth equation. Ecol. Model. 2016, 320, 177–181. [Google Scholar] [CrossRef]
- Cao, L.; Shi, P.; Li, L.; Chen, G. A new flexible sigmoidal growth model. Symmetry 2019, 11, 204. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 30 May 2022).
- Shi, P.; Ridland, P.M.; Ratkowsky, D.A.; Li, Y. IPEC: Root Mean Square Curvature Calculation, R package version 1.0.3; CRAN: Vienna, Austria, 2022; Available online: https://cran.r-project.org/web/packages/IPEC/index.html (accessed on 24 May 2022).
- Quinn, B.K. A critical review of the use and performance of different function types for modeling temperature-dependent development of arthropod larvae. J. Therm. Biol. 2017, 63, 65–77. [Google Scholar] [CrossRef]
- Nelder, J.A.; Mead, R. A simplex method for function minimization. Comput. J. 1965, 7, 308–313. [Google Scholar] [CrossRef]
- Yu, X.; Shi, P.; Schrader, J.; Niklas, K.J. Nondestructive estimation of leaf area for 15 species of vines with different leaf shapes. Amer. J. Bot. 2020, 107, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Reddy, G.V.P.; Schrader, J.; Guo, X.; Li, Y.; Jiao, Y.; Shi, P. A nondestructive method of calculating the wing area of insects. Ecol. Evol. 2022, 12, e8792. [Google Scholar] [CrossRef] [PubMed]
- Shipley, B.; Vu, T.-T. Dry matter content as a measure of dry matter concentration in plants and their parts. New Phytol. 2002, 153, 359–364. [Google Scholar] [CrossRef]
- Shi, P.; Gielis, J.; Quinn, B.K. ‘biogeom’: Biological Geometries, R package version 1.0.6.; CRAN: Vienna, Austria, 2022; Available online: https://cran.r-project.org/web/packages/biogeom/index.html (accessed on 30 May 2022).
- Shi, P.; Gielis, J.; Quinn, B.K.; Niklas, K.J.; Ratkowsky, D.A.; Schrader, J.; Ruan, H.; Wang, L.; Niinemets, Ü. ‘biogeom’: An R package for simulating and fitting natural shapes. Ann. N. Y. Acad. Sci. 2022; in press. [Google Scholar] [CrossRef]
- Spiess, A.-N.; Neumeyer, N. An evaluation of R squared as an inadequate measure for nonlinear models in pharmacological and biochemical research: A Monte Carlo approach. BMC Pharmacol. 2010, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Gompertz, B. On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies. Philos. Trans. R. Soc. Lond. 1825, 115, 513–583. [Google Scholar]
- Lobry, J.R.; Rosso, L.; Flandrois, J.P. A FORTRAN subroutine for the determination of parameter confidence limits in non-linear models. Binary 1991, 3, 86–93. [Google Scholar]
- Rosso, L.; Lobry, J.R.; Flandrois, J.P. An unexpected correlation between cardinal temperatures of microbial growth highlighted by a new model. J. Theor. Biol. 1993, 162, 447–463. [Google Scholar] [CrossRef]
- Shi, P.; Yu, K.; Niklas, K.J.; Schrader, J.; Song, Y.; Zhu, R.; Li, Y.; Wei, H.; Ratkowsky, D.A. A general model for describing the ovate leaf shape. Symmetry 2021, 13, 1524. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Quinn, B.K.; Shi, P. The Modified Brière Equation and Its Applications. Plants 2022, 11, 1769. https://doi.org/10.3390/plants11131769
Jin J, Quinn BK, Shi P. The Modified Brière Equation and Its Applications. Plants. 2022; 11(13):1769. https://doi.org/10.3390/plants11131769
Chicago/Turabian StyleJin, Jun, Brady K. Quinn, and Peijian Shi. 2022. "The Modified Brière Equation and Its Applications" Plants 11, no. 13: 1769. https://doi.org/10.3390/plants11131769