
Complete Genome Sequence of a Novel Monopartite Mastrevirus, Soybean Geminivirus B, Isolated from Soybean (Glycine max (L.) Merrill)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soybean Samples
2.2. Library Preparation and RNA-Sequencing
2.3. Bioinformatics Analyses to Identify Virus-Associated Contigs
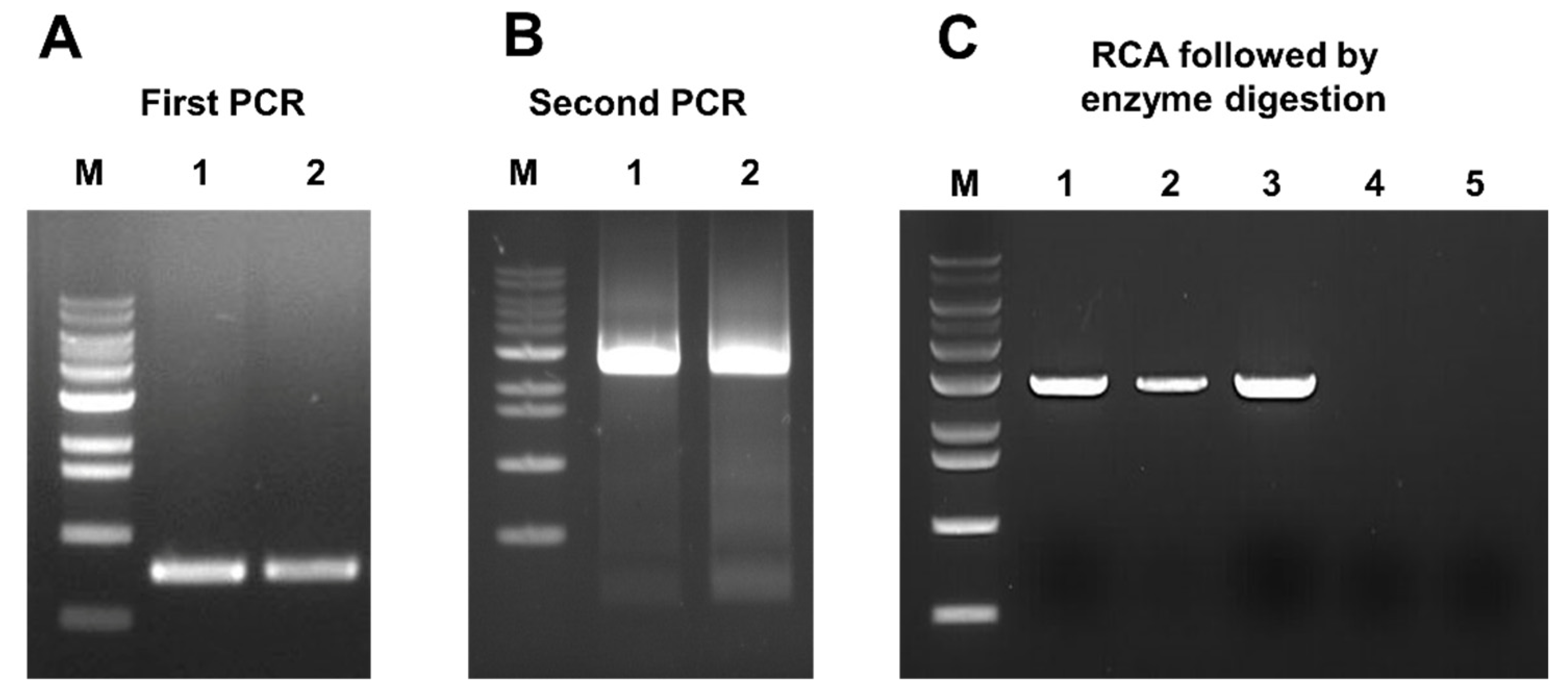
2.4. Complete Genome Sequencing of SGVB
2.5. Prediction of Open Reading Frames (ORFs), Conserved Domains, and Stem-Loop Structure
2.6. Pairwise Sequence Alignment and Visualization
2.7. Construction of Phylogenetic Trees
2.8. Construction of Full-Length Infectious Clones for SGVB
2.9. Plant Materials, Growth Conditions, and Agroinoculation
3. Results
3.1. Identification of a Novel DNA Virus in Soybean Using RNA-Sequencing
3.2. Complete Genome Sequencing of SGVB
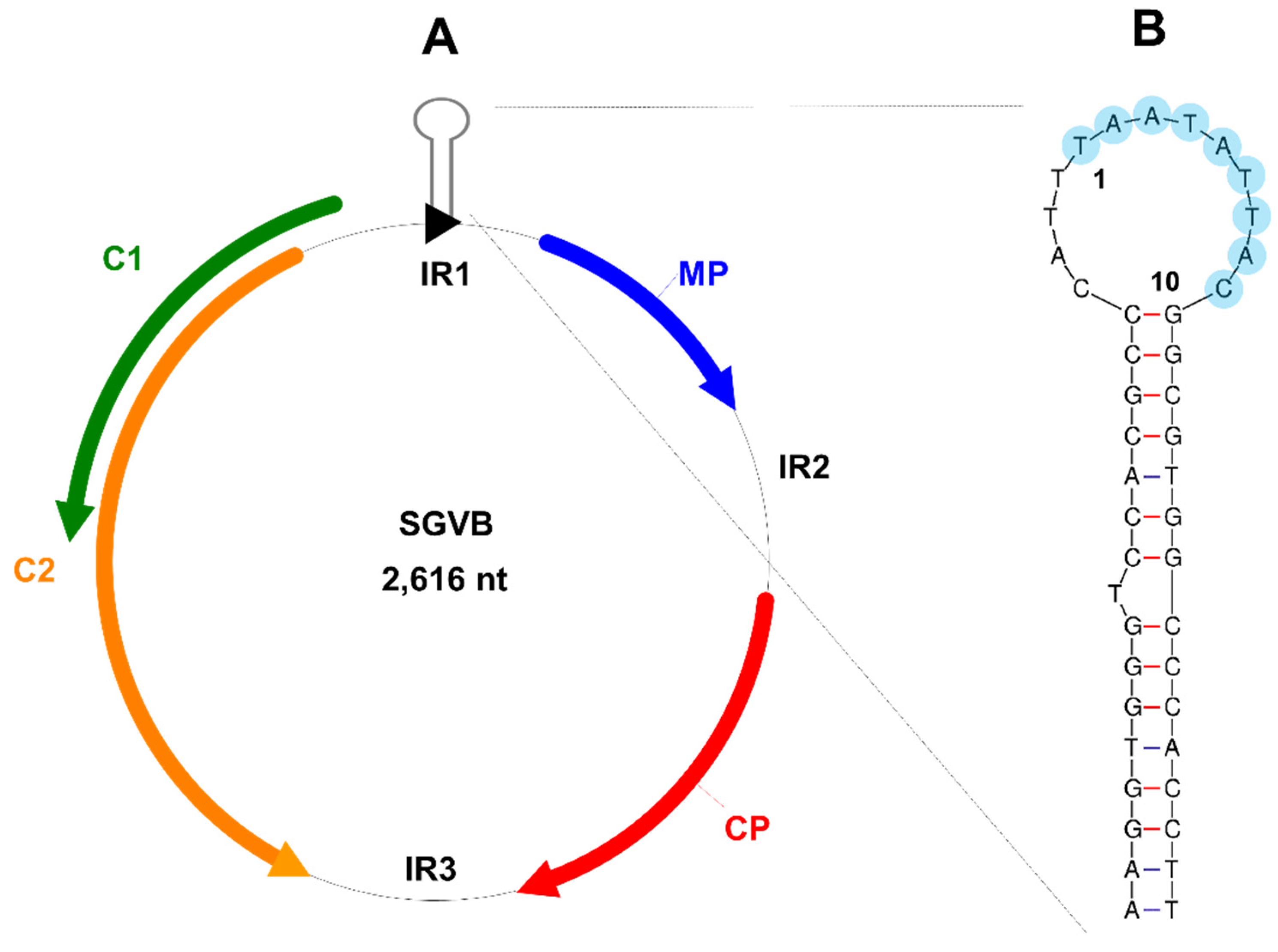
3.3. Genome Organization of SGVB
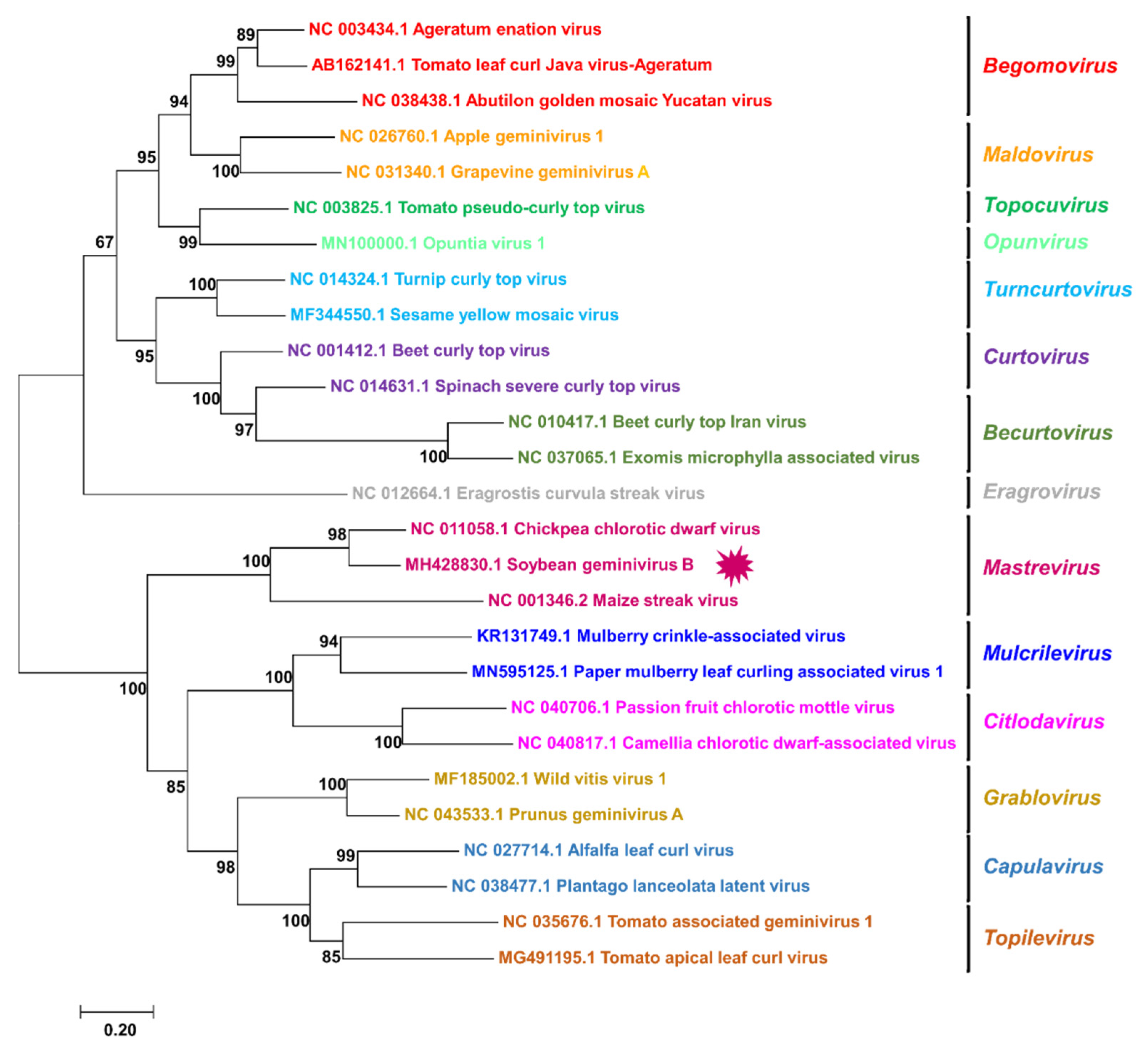
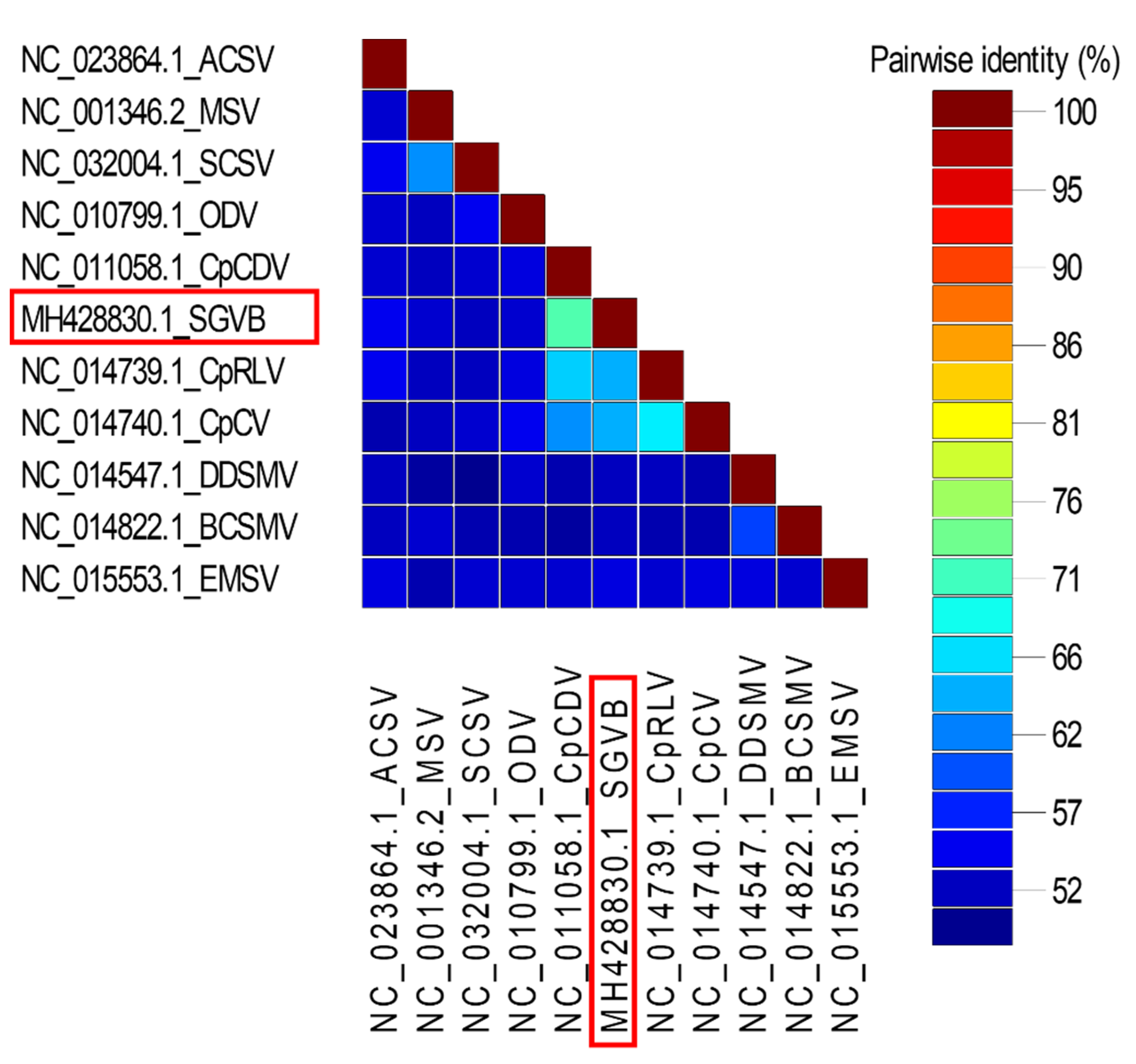
3.4. Phylogenetic Relationship of SGVB with Other Geminiviruses
3.5. Generation of Infectious Clones for SGVB and Disease Symptoms Caused by SGVB
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varma, A.; Malathi, V. Emerging geminivirus problems: A serious threat to crop production. Ann. Appl. Biol. 2003, 142, 145–164. [Google Scholar] [CrossRef]
- Ramesh, S.; Sahu, P.; Prasad, M.; Praveen, S.; Pappu, H. Geminiviruses and plant hosts: A closer examination of the molecular arms race. Viruses 2017, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Zerbini, F.M.; Briddon, R.W.; Idris, A.; Martin, D.P.; Moriones, E.; Navas-Castillo, J.; Rivera-Bustamante, R.; Roumagnac, P.; Varsani, A. ICTV virus taxonomy profile: Geminiviridae. J. Gen. Virol. 2017, 98, 131–133. [Google Scholar] [CrossRef]
- Straub, S.C.; Pfeil, B.E.; Doyle, J.J. Testing the polyploid past of soybean using a low-copy nuclear gene? Is Glycine (Fabaceae: Papilionoideae) an auto-or allopolyploid? Mol. Phylogenetics Evol. 2006, 39, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.H.; Whitham, S.A. Control of virus diseases in soybeans. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 90, pp. 355–390. [Google Scholar]
- Fernandes, F.R.; Cruz, A.; Faria, J.; Zerbini, F.; Aragão, F.J. Three distinct begomoviruses associated with soybean in central Brazil. Arch. Virol. 2009, 154, 1567–1570. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Seo, E.-Y.; Cho, S.; Kim, J.; Chung, J.; Lim, H.-J.; Gotoh, T.; Hammond, J.; Lim, H.-S. A 2014 nationwide survey of the distribution of soybean mosaic virus (SMV), soybean yellow mottle mosaic virus (SYMMV) and soybean yellow common mosaic virus (SYCMV) major viruses in South Korean soybean fields, and changes from 2012 Isolate prevalence. J. Fac. Agric. Kyushu Univ. 2015, 60, 339–347. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Yoon, Y.-N.; Yun, H.-T.; Baek, I.; Lim, S.; Moon, J.; Lee, S.-H. First report of Bean common mosaic virus infecting soybean in South Korea. Plant Dis. 2015, 99, 1189. [Google Scholar] [CrossRef]
- Kim, S.-M.; Lee, J.-B.; Lee, Y.-H.; Choi, S.-H.; Choi, H.-S.; Park, J.-W.; Lee, J.-S.; Lee, G.-S.; Moon, J.-K.; Moon, J.-S. First report of Soybean dwarf virus on soybean (Glycine max) in Korea. Res. Plant Dis. 2006, 12, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-C.; Kim, M.-K.; Kwak, H.-R.; Choi, H.-S.; Kim, J.-S.; Park, C.-Y.; Lee, S.-H.; Cha, B. First report of clover yellow vein virus on Glycine max in Korea. Plant Dis. 2014, 98, 1283. [Google Scholar] [CrossRef]
- Nam, M.; Park, S.-J.; Kim, Y.-J.; Kim, J.-S.; Park, C.-Y.; Lee, J.-S.; Choi, H.-S.; Kim, J.-S.; Kim, H.-G.; Lee, S.-H. First report of Peanut stunt virus on Glycine max in Korea. Plant Pathol. J. 2012, 28, 330. [Google Scholar] [CrossRef]
- Yoon, Y.; Jo, Y.; Cho, W.; Choi, H.; Jang, Y.; Lee, Y.; Bae, J.; Lee, B. First report of Tomato spotted wilt virus infecting soybean in Korea. Plant Dis. 2018, 102, 461. [Google Scholar] [CrossRef]
- Jang, Y.; Jo, Y.; Cho, W.; Choi, H.; Yoon, Y.; Lim, S.; Lee, Y.; Bae, J.; Lee, B. First report of bean common mosaic necrosis virus infecting soybean in Korea. Plant Dis. 2018, 102, 2051. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Yoon, Y.N.; Jang, Y.-W.; Choi, H.; Lee, Y.-H.; Kim, S.-M.; Choi, S.Y.; Lee, B.C.; Cho, W.K. Soybean Viromes in the Republic of Korea Revealed by RT-PCR and Next-Generation Sequencing. Microorganisms 2020, 8, 1777. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494. [Google Scholar] [CrossRef]
- Letunic, I.; Copley, R.R.; Pils, B.; Pinkert, S.; Schultz, J.; Bork, P. SMART 5: Domains in the context of genomes and networks. Nucleic Acids Res. 2006, 34, D257–D260. [Google Scholar] [CrossRef] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Choi, H.; Kim, S.-M.; Kim, S.-L.; Lee, B.C.; Cho, W.K. The pepper virome: Natural co-infection of diverse viruses and their quasispecies. BMC Genom. 2017, 18, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenele, R.S.; Lamas, N.S.; Lacorte, C.; Lacerda, A.L.M.; Varsani, A.; Ribeiro, S.G. A novel geminivirus identified in tomato and cleome plants sampled in Brazil. Virus Res. 2017, 240, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Al Rwahnih, M.; Alabi, O.J.; Westrick, N.M.; Golino, D.; Rowhani, A. Description of a novel monopartite geminivirus and its defective subviral genome in grapevine. Phytopathology 2016, 107, 240–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, P.; Navarro, B.; Zhang, Z.; Wang, H.; Lu, M.; Xiao, H.; Wu, Q.; Zhou, X.; Di Serio, F.; Li, S. Identification and characterization of a novel geminivirus with a monopartite genome infecting apple trees. J. Gen. Virol. 2015, 96, 2411–2420. [Google Scholar] [CrossRef]
- Jo, Y.; Lian, S.; Chu, H.; Cho, J.K.; Yoo, S.-H.; Choi, H.; Yoon, J.-Y.; Choi, S.-K.; Lee, B.C.; Cho, W.K. Peach RNA viromes in six different peach cultivars. Sci. Rep. 2018, 8, 1844. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.; Choi, H.; Cho, J.K.; Yoon, J.-Y.; Choi, S.-K.; Cho, W.K. In silico approach to reveal viral populations in grapevine cultivar Tannat using transcriptome data. Sci. Rep. 2015, 5, 15841. [Google Scholar] [CrossRef] [Green Version]
- Rector, A.; Tachezy, R.; Van Ranst, M. A sequence-independent strategy for detection and cloning of circular DNA virus genomes by using multiply primed rolling-circle amplification. J. Virol. 2004, 78, 4993–4998. [Google Scholar] [CrossRef] [Green Version]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef]
- Johne, R.; Müller, H.; Rector, A.; Van Ranst, M.; Stevens, H. Rolling-circle amplification of viral DNA genomes using phi29 polymerase. Trends Microbiol. 2009, 17, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Huang, Q.; Wu, L.; Qian, Y. Identification and characterization of a maize-associated mastrevirus in China by deep sequencing small RNA populations. Virol. J. 2015, 12, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Zhang, S.; Yu, H.; Xuan, Z.; Yang, L.; Zhan, B.; Murilo Zerbini, F.; Cao, M. Identification and characterization of two novel geminiviruses associated with paper mulberry (Broussonetia papyrifera) leaf curl disease. Plant Dis. 2020, 104, 3010–3018. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Kim, S.-M.; Choi, H.; Yang, J.W.; Lee, B.C.; Cho, W.K. Sweet potato viromes in eight different geographical regions in Korea and two different cultivars. Sci. Rep. 2020, 10, 2588. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Song, W.; Kwak, H.-R.; Kim, J.-d.; Park, J.; Auh, C.-K.; Kim, D.-H.; Lee, K.-y.; Lee, S.; Choi, H.-S. Phylogenetic analysis and inflow route of Tomato yellow leaf curl virus (TYLCV) and Bemisia tabaci in Korea. Mol. Cells 2010, 30, 467–476. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Lai, Y.-C.; Lin, N.-S.; Hsu, Y.-H.; Tsai, H.-T.; Liao, J.-Y.; Hu, C.-C. A simplified method of constructing infectious clones of begomovirus employing limited restriction enzyme digestion of products of rolling circle amplification. J. Virol. Methods 2008, 147, 355–359. [Google Scholar] [CrossRef]
- Ferro, M.; Ramos-Sobrinho, R.; Xavier, C.; Zerbini, F.; Lima, G.; Nagata, T.; Assunção, I. New approach for the construction of infectious clones of a circular DNA plant virus using Gibson Assembly. J. Virol. Methods 2019, 263, 20–23. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Position | Size (aa) or (nt) | Homologous Protein | Homologous Virus | Accession No. | Coverage | Identity |
|---|---|---|---|---|---|---|---|
| MP | 145–438 (sense) | 97 aa | Movementprotein | CpCDV | AIY33051.1 | 92% | 68.89% |
| CP | 705–1184 (sense) | 159 aa | Coatprotein | Chickpea redleaf virus 2 | QDA77210.1 | 100% | 89.94% |
| C1 | 2005–2496 (antisense) | 163 aa | RepB | CpCDV | AHF52852.1 | 69% | 71.93% |
| C2 | 1550–2431 (antisense) | 138 aa | RepA | CpCDV | AMN14229.1 | 100% | 77.47% |
| IR1 | 2497–2616, 1–144 | 264 nt | |||||
| IR2 | 439–704 | 266 nt | |||||
| IR3 | 1185–1449 | 265 nt |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.; Jo, Y.; Hong, J.; Chung, H.; Choi, S.; Kim, S.; Lee, J.; Moh, S.; Lee, B.; Cho, W.K. Complete Genome Sequence of a Novel Monopartite Mastrevirus, Soybean Geminivirus B, Isolated from Soybean (Glycine max (L.) Merrill). Plants 2022, 11, 1768. https://doi.org/10.3390/plants11131768
Choi H, Jo Y, Hong J, Chung H, Choi S, Kim S, Lee J, Moh S, Lee B, Cho WK. Complete Genome Sequence of a Novel Monopartite Mastrevirus, Soybean Geminivirus B, Isolated from Soybean (Glycine max (L.) Merrill). Plants. 2022; 11(13):1768. https://doi.org/10.3390/plants11131768
Chicago/Turabian StyleChoi, Hoseong, Yeonhwa Jo, Jinsung Hong, Hyunjung Chung, Sooyeon Choi, Sangmin Kim, Jeonghun Lee, Sanghyun Moh, Bongchoon Lee, and Won Kyong Cho. 2022. "Complete Genome Sequence of a Novel Monopartite Mastrevirus, Soybean Geminivirus B, Isolated from Soybean (Glycine max (L.) Merrill)" Plants 11, no. 13: 1768. https://doi.org/10.3390/plants11131768