
Genetic and Morphological Diversity Assessment of Five Kalanchoe Genotypes by SCoT, ISSR and RAPD-PCR Markers

 ,
,  ,
,  ,
,  ,
, 
 ,
,  , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
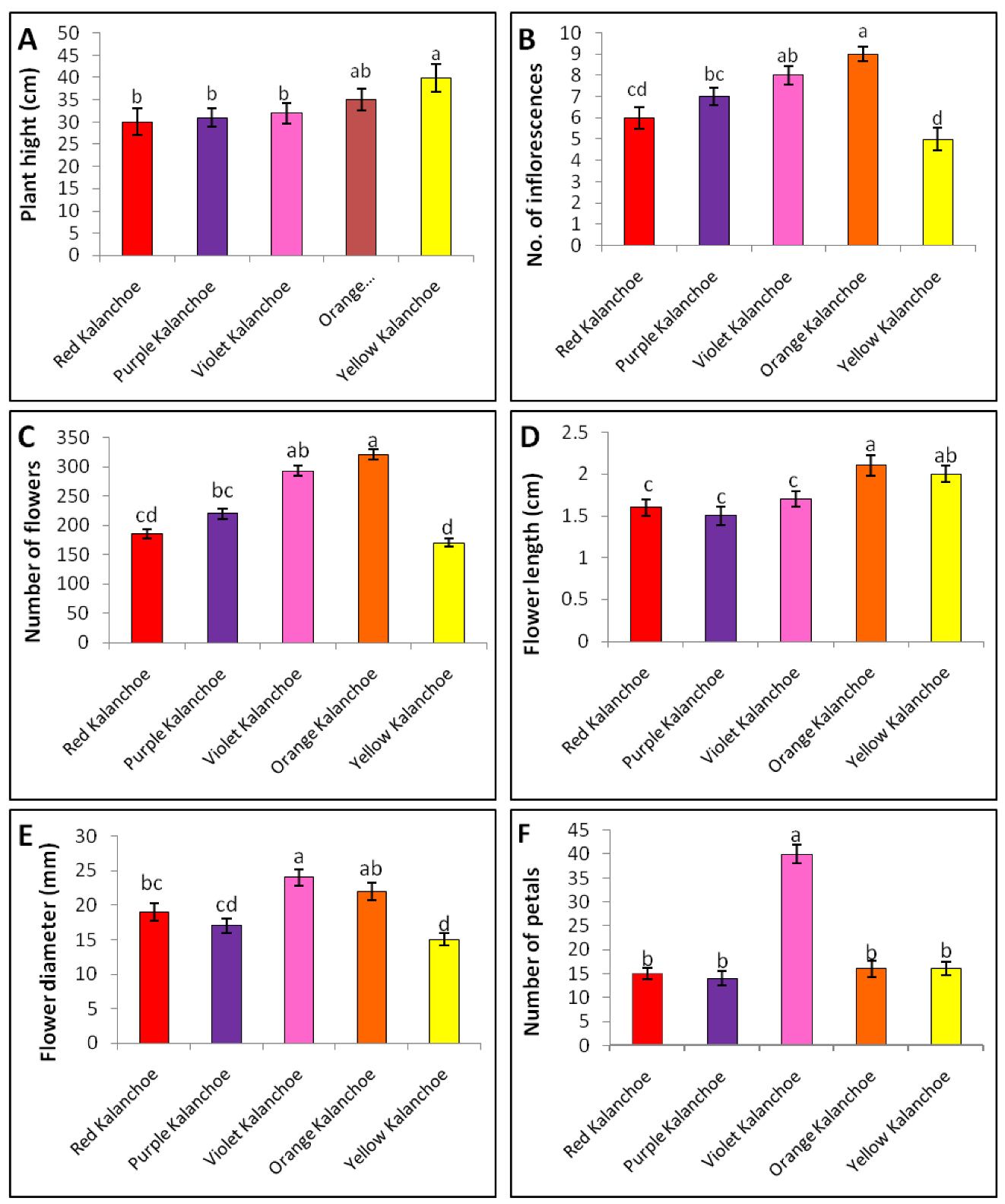
2.1. Morphological Polymorphism
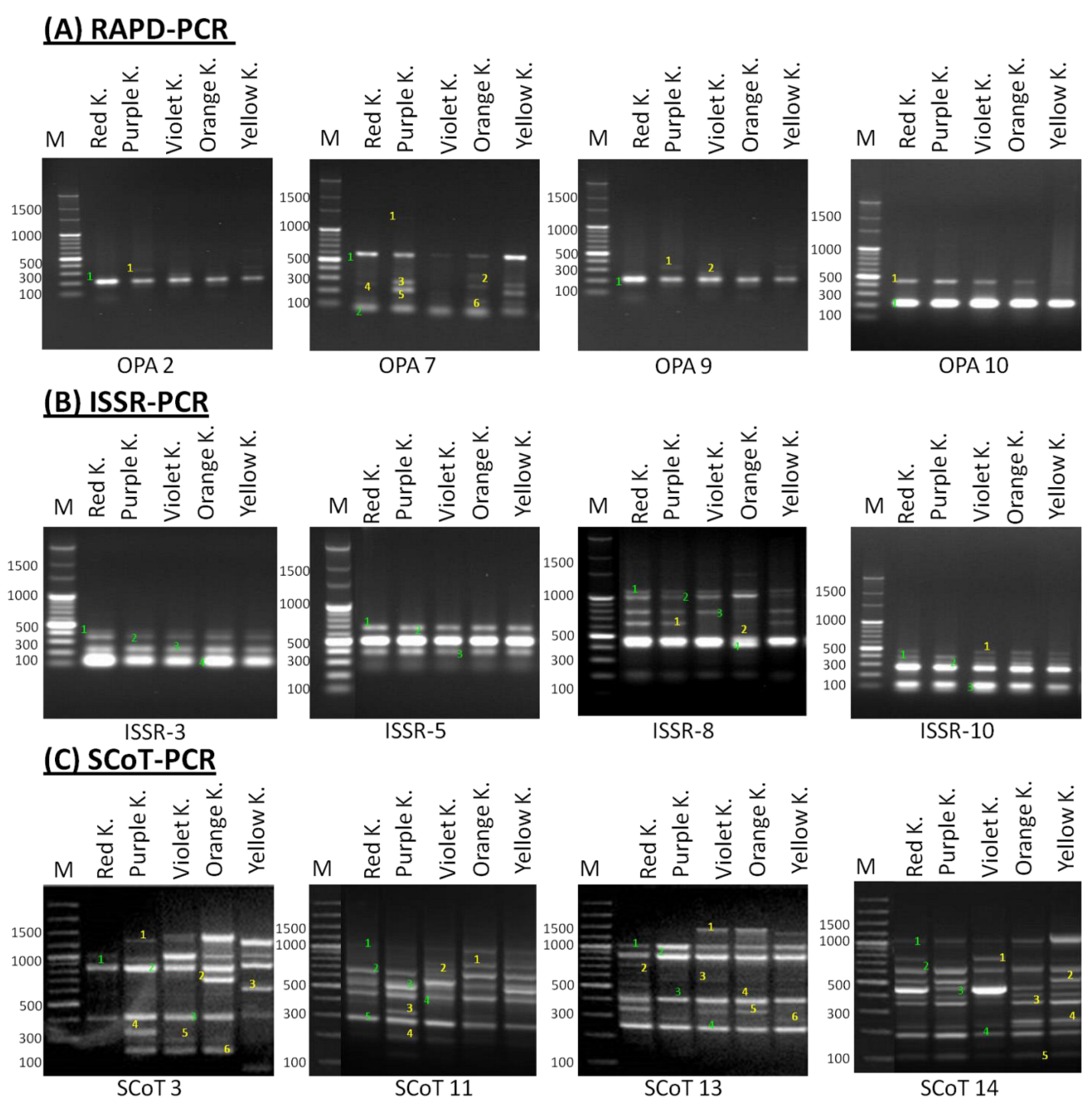
2.2. Molecular Polymorphism Analyses
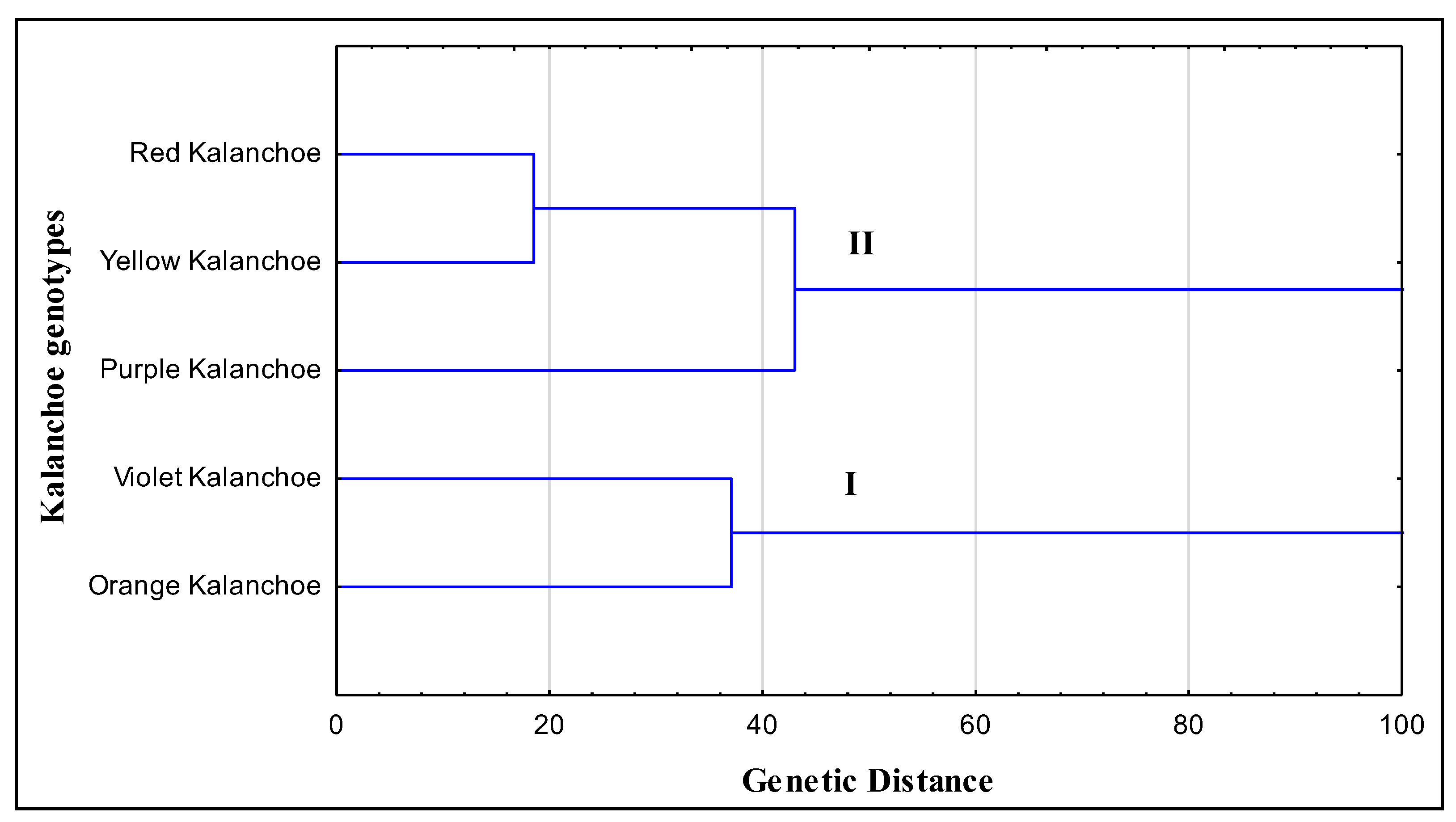
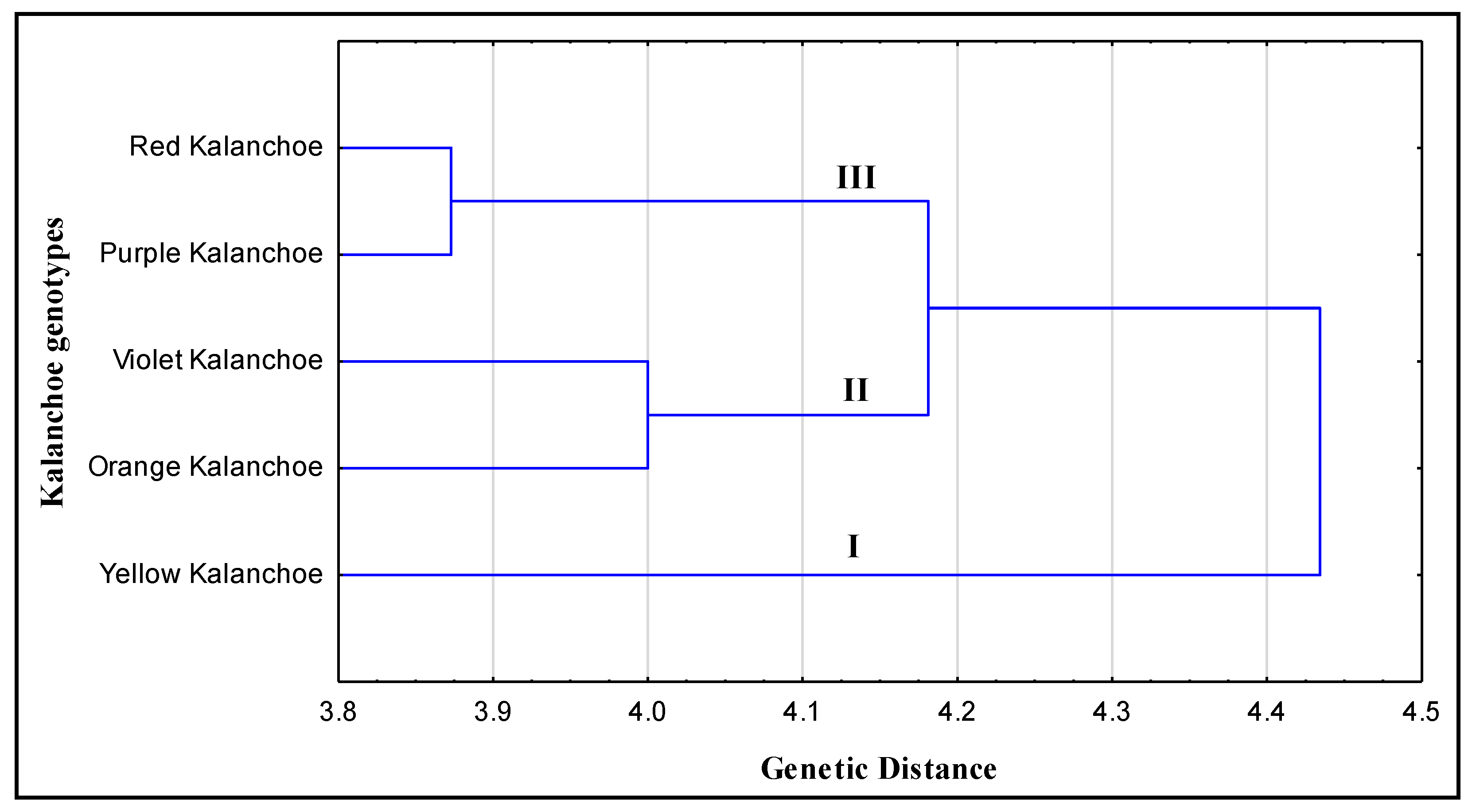
2.3. Phylogeny Analyses
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Morphological Polymorphism
4.3. Genomic DNA Extraction
4.4. Random Amplified Polymorphism DNA (RAPD-PCR)
4.5. Inter-Simple Sequence Repeats (ISSR-PCR)
4.6. Start Codon Targeted (SCoT) Amplification
4.7. Gel Electrophoresis
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lunkad, A.; Agrawal, M.; Kothawade, S. Anthelmintic activity of Bryophyllum pinnatum. J. Pharmacogn. Phytochem. 2016, 8, 21. [Google Scholar]
- Dole, J.M.; Wilkins, H.F.; Dole, J.M.; Wilkins, H.F. Kalanchoe; Pearson Prentice Hall: Hoboken, NJ, USA, 2005; pp. 629–635. [Google Scholar]
- Boiteau, P.; Allorge-Boiteau, L. Kalanchoe (Crassulacées) de Madagascar: Systématique, Écophysiologie et Phytochimie; KARTHALA Editions: Paris, France, 1995. [Google Scholar]
- Kluge, M.; Brulfert, J. Crassulacean acid metabolism in the genus Kalanchoë: Ecological, physiological and biochemical aspects. In Crassulacean Acid Metabolism; Springer: Berlin/Heidelberg, Germany, 1996; pp. 324–335. [Google Scholar]
- Möller, M.; Cronk, Q. Evolution of morphological novelty: A phylogenetic analysis of growth patterns in Streptocarpus (Gesneriaceae). Evolution 2001, 55, 918–929. [Google Scholar] [CrossRef]
- Hassanin, A.A.; Saad, A.M.; Bardisi, E.A.; Salama, A.; Sitohy, M.Z. Transfer of Anthocyanin Accumulating Delila and Rosea1 Genes from the Transgenic Tomato Micro-Tom Cultivar to Moneymaker Cultivar by Conventional Breeding. J. Agric. Food Chem. 2020, 68, 10741–10749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hong, Y.; Liao, Y.; Tian, K.; Sun, H.; Liu, X.; Tang, Y.; Hassanin, A.A.; Abdelnour, S.A.; Suthikrai, W.; et al. Dietary Lasia spinosa Thw. Improves Growth Performance in Broilers. Front. Nutr. 2022, 8, 775223. [Google Scholar] [CrossRef]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Hassanin, A.A.; Eldomiaty, A.S.; Ujjan, J.A.; Al-Mushhin, A.A.M.; Alrashidi, A.A.; Saad, A.M.; Sakit Alhaithloul, H.A.; El-Saadony, M.T.; Awad, M.F.; Sitohy, M.Z. Assessment of the R2R3 MYB gene expression profile during tomato fruit development using in silico analysis, quantitative and semi-quantitative RT-PCR. Saudi J. Biol. Sci. 2022. [Google Scholar] [CrossRef]
- Krzymińska, A.; Gawłowska, M.; Wolko, B.; Bocianowski, J. Genetic diversity of ornamental Allium species and cultivars assessed with isozymes. J. Appl. Genet. 2008, 49, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Abeyot, T.; Sentayehu, A.; Taye, K.; Weyessa, G. Genetic diversity analysis for quality attributes of some promising Coffea arabica germplasm collections in southwestern Ethiopia. Sci. J. Biol. Sci. 2011, 11, 236–244. [Google Scholar]
- Soleimani, M.H.; Talebi, M.; Sayed-Tabatabaei, B.E. Use of SRAP markers to assess genetic diversity and population structure of wild, cultivated, and ornamental pomegranates (Punica granatum L.) in different regions of Iran. Plant Syst. Evol. 2012, 298, 1141–1149. [Google Scholar] [CrossRef]
- Fortunato, F.L.G.; do Rêgo, E.R.; de Carvalho, M.G.; dos Santos, C.A.P.; do Rêgo, M.M. Genetic diversity in ornamental pepper plants. Comun. Sci. 2019, 10, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Ning, G.; Gao, L.; Bao, M. Genetic diversity of Dianthus accessions as assessed using two molecular marker systems (SRAPs and ISSRs) and morphological traits. Sci. Hortic. 2008, 117, 263–270. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, X.-Q.; Zhou, Y.-H.; Bai, S.-Q.; Liu, W. Assessing genetic diversity of Elymus sibiricus (Poaceae: Triticeae) populations from Qinghai-Tibet Plateau by ISSR markers. Biochem. Syst. Ecol. 2008, 36, 514–522. [Google Scholar] [CrossRef]
- Yan, J.; Bai, S.; Zhang, X.; You, M.; Zhang, C.; Li, D.; Zeng, Y. Genetic diversity of wild Elymus sibiricus germplasm from the Qinghai-Tibetan Plateau in China detected by SRAP markers. Acta Prataculturae Sin. 2010, 19, 173–183. [Google Scholar]
- Collard, B.C.; Mackill, D.J. Start codon targeted (SCoT) polymorphism: A simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol. Biol. Rep. 2009, 27, 86–93. [Google Scholar] [CrossRef]
- Bertan, I.; de Carvalho, F.I.; de Oliveira, A.C. Parental selection strategies in plant breeding programs. J. Crop Sci. Biotechnol. 2007, 10, 211–222. [Google Scholar]
- Ceballos, H.; Iglesias, C.A.; Pérez, J.C.; Dixon, A.G. Cassava breeding: Opportunities and challenges. Plant Mol. Biol. 2004, 56, 503–516. [Google Scholar] [CrossRef]
- Huang, C.-H.; Chu, C.-Y. Inheritance of leaf and flower morphologies in Kalanchoe spp. Euphytica 2017, 213, 4. [Google Scholar] [CrossRef]
- Agarwal, M.; Shrivastava, N.; Padh, H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008, 27, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasution, F.; Theanhom, A.A.; Bhuyar, P.; Chumpookam, J. Genetic diversity evaluation in wild Muntingia calabura L. based on Random Amplified Polymorphic DNA (RAPD) markers. Gene Rep. 2021, 25, 101335. [Google Scholar] [CrossRef]
- Sadhu, S.; Jogam, P.; Thampu, R.K.; Abbagani, S.; Penna, S.; Peddaboina, V. High efficiency plant regeneration and genetic fidelity of regenerants by SCoT and ISSR markers in chickpea (Cicer arietinum L.). Plant Cell Tissue Organ Cult. 2020, 141, 465–477. [Google Scholar] [CrossRef]
- Eslahi, M.; Safaie, N.; Saidi, A.; Shams-Bakhsh, M. cDNA-AFLP analysis of plant defense genes expressed in wheat (cv. Chamran) infected with Mycosphaerella graminicola. J. Agric. Sci. Technol. 2021, 23, 699–710. [Google Scholar]
- Leipold, M.; Tausch, S.; Hirtreiter, M.; Poschlod, P.; Reisch, C. Sampling for conservation genetics: How many loci and individuals are needed to determine the genetic diversity of plant populations using AFLP? Conserv. Genet. Resour. 2020, 12, 99–108. [Google Scholar] [CrossRef]
- Thakur, M.; Sharma, V.; Chauhan, A. Genetic fidelity assessment of long term in vitro shoot cultures and regenerated plants in Japanese plum cvs Santa Rosa and Frontier through RAPD, ISSR and SCoT markers. S. Afr. J. Bot. 2021, 140, 428–433. [Google Scholar] [CrossRef]
- Luo, C.; He, X.-H.; Chen, H.; Ou, S.-J.; Gao, M.-P. Analysis of diversity and relationships among mango cultivars using start codon targeted (SCoT) markers. Biochem. Syst. Ecol. 2010, 38, 1176–1184. [Google Scholar] [CrossRef]
- Gorji, A.M.; Poczai, P.; Polgar, Z.; Taller, J. Efficiency of arbitrarily amplified dominant markers (SCoT, ISSR and RAPD) for diagnostic fingerprinting in tetraploid potato. Am. Potato J. 2011, 88, 226–237. [Google Scholar] [CrossRef]
- Raza, S.H.A.; Hassanin, A.A.; Dhshan, A.I.M.; Abdelnour, S.A.; Khan, R.; Mei, C.; Zan, L. In silico genomic and proteomic analyses of three heat shock proteins (HSP70, HSP90-α, and HSP90-β) in even-toed ungulates. Electron. J. Biotechnol. 2021, 53, 61–70. [Google Scholar] [CrossRef]
- Fathy, D.M.; Eldomiaty, A.S.; El-Fattah, H.I.A.; Mahgou, E.-S.I.; Hassanin, A.A. Morphological, Biochemical and Molecular Characterization of Rhizobia of Faba Bean Plants Grown in North Nile Delta Egypt. Pak. J. Biol. Sci. 2021, 24, 672–679. [Google Scholar] [CrossRef]
- Fang, E.F.; Hassanien, A.A.E.; Wong, J.H.; Bah, C.S.F.; Soliman, S.S.; Ng, T.B. Isolation of a New Trypsin Inhibitor from the Faba Bean (Vicia faba cv. Giza 843) with Potential Medicinal Applications. Protein Pept. Lett. 2011, 18, 64–72. [Google Scholar] [CrossRef]
- Fang, E.F.; Hassanien, A.A.E.; Wong, J.H.; Bah, C.S.F.; Soliman, S.S.; Ng, T.B. Purification and Modes of Antifungal Action by Vicia faba cv. Egypt Trypsin Inhibitor. J. Agric. Food Chem. 2010, 58, 10729–10735. [Google Scholar] [CrossRef]
- Farrag, M.E.; Soliman, S.S.; Amer, I.M.; Heakel, R.M.; Hassanin, A.A. Analysis of Genetic Diversity among a Population of Canola Genotypes As Reveled By ISSR-PCR and Their Associations to Seed Yield and Oil Content. Ann. Agric. Sci. Moshtohor 2019, 57, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Hassanin, A.A.; Haidar Abbas Raza, S.; Ahmed Ujjan, J.; Aysh Alrashidi, A.; Sitohy, B.M.; Al-surhanee, A.A.; Saad, A.M.; Mohamed Al-Hazani, T.; Osman Atallah, O.; Al Syaad, K.M.; et al. Emergence, evolution, and vaccine production approaches of SARS-CoV-2 virus: Benefits of getting vaccinated and common questions. Saudi J. Biol. Sci. 2022, 29, 1981–1997. [Google Scholar] [CrossRef]
- Vieira, E.A.; de Carvalho, F.I.F.; de Oliveira, A.C.; Benin, G.; Zimmer, P.D.; da Silva, J.A.G.; Martins, A.F.; Bertan, I.; da Silva, G.O.; Schmidt, D.A.M. Comparison among pedigree, morphological and molecular distance measures in oats (Avena sativa) in experiments with and without fungicide. Bragantia 2005, 64, 51–60. [Google Scholar] [CrossRef]
- Hassanin, A.A.; Soliman, S.S.A.; Ismail, T.A.; Amin, M.K.A. The role of slmyb gene in tomato fruit development. Zagazig J. Agric. Res. 2017, 44, 969–988. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Xie, L.; Hassanin, A.A.; Zuo, E.; Lu, Y. The Potential of CRISPR/Cas9 Gene Editing as a Treatment Strategy for Inherited Diseases. Front. Cell Dev. Biol. 2021, 9, 699597. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Hassanin, A.A.; Pant, S.D.; Bing, S.; Sitohy, M.Z.; Abdelnour, S.A.; Alotaibi, M.A.; Al-Hazani, T.M.; Abd El-Aziz, A.H.; Cheng, G.; et al. Potentials, prospects and applications of genome editing technologies in livestock production. Saudi J. Biol. Sci. 2022, 29, 1928–1935. [Google Scholar] [CrossRef]
- Traoré, L.T.; Kuligowska, K.; Lütken, H.; Müller, R. Stigma development and receptivity of two Kalanchoë blossfeldiana cultivars. Acta Physiol. Plant. 2014, 36, 1763–1769. [Google Scholar] [CrossRef]
- Scobeyeva, V.; Omelchenko, D.; Dyakov, L.; Konovalov, A.; Speranskaya, A.; Krinitsina, A. Comparison of some plant DNA extraction methods. Russ. J. Genet. 2018, 54, 576–586. [Google Scholar] [CrossRef]
- Moreno, S.; Martín, J.P.; Ortiz, J.M. Inter-simple sequence repeats PCR for characterization of closely related grapevine germplasm. Euphytica 1998, 101, 117–125. [Google Scholar] [CrossRef]
- Maniatis, T. Molecular Cloning. Decontamination of Dilute Solutions of Ethidium Bromide. 1989. Available online: https://www.cabdirect.org/cabdirect/abstract/19901616061 (accessed on 16 June 2022).
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Norušis, M.J. SPSS for Windows, release 6.0; Prentice Hall (Higher Education Division, Pearson Education): Hoboken, NJ, USA, 1993.
- Rokach, L.; Maimon, O. Clustering methods. In Data Mining and Knowledge Discovery Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 321–352. [Google Scholar] [CrossRef]
- Weiß, C.H. Statistica, Version 8; Statsoft, Inc.: Tulsa, OK, USA, 2007. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Type | Primer | Number of Loci | Monomorphic Loci | Polymorphic Loci | Unique Loci | Polymorphism (%) | Percentage Fidelity of the RAPD/SCoT |
|---|---|---|---|---|---|---|---|
| RAPD | OPA 2 | 2 | 1 | 1 | 0 | 50% | 4/5 × 100− 15/38 × 100 = 80 − 39.47 = 40.53% |
| OPA 7 | 8 | 2 | 6 | 2 | 75% | ||
| OPA 9 | 3 | 1 | 2 | 2 | 66% | ||
| OPA 10 | 2 | 1 | 1 | 0 | 50% | ||
| Average | 3.75 | 1.25 | 2.5 | 1 | 60.25% | ||
| Total | 15 | 5 | 10 | 4 | |||
| ISSR | ISSR-3 | 4 | 4 | 0 | 0 | 0% | |
| ISSR-5 | 3 | 3 | 0 | 0 | 0% | ||
| ISSR-8 | 6 | 4 | 2 | 0 | 33% | ||
| ISSR-10 | 4 | 3 | 1 | 0 | 25% | ||
| Average | 4.25 | 3.5 | 0.75 | 0 | 15% | ||
| Total | 17 | 14 | 3 | 0 | |||
| SCoT | SCoT 3 | 9 | 3 | 6 | 3 | 66% | |
| SCoT 11 | 9 | 5 | 4 | 1 | 44% | ||
| SCoT 13 | 11 | 4 | 7 | 1 | 63% | ||
| SCoT 14 | 9 | 4 | 5 | 0 | 55% | ||
| Average | 9.5 | 4 | 5.5 | 1.25 | 57% | ||
| Total | 38 | 16 | 22 | 5 | |||
| Total number of loci | 70 | 35 | 35 | 9 |
| No. | RAPD Primers | ISSR Primers | SCoT Primers | |||
|---|---|---|---|---|---|---|
| Code | Sequence ′3–5′ | Code | Sequence ′3–5′ | Code | Sequence ′3–5′ | |
| 1 | OPA2 | TGCCGAGCTG | ISSR-1 | (ga) 6 gg | SCoT 2 | ACCATGGCTACCACCGGC |
| 2 | OPA7 | GAAACGGGTG | ISSR-2 | (cac)3 gc | SCoT 3 | ACGACATGGCGACCCACA |
| 3 | OPA9 | GGGTAACGCC | ISSR-3 | (gag) 3 gc | SCoT 4 | ACCATGGCTACCACCGCA |
| 4 | OPA10 | CTGCTGGGAC | ISSR-4 | cac (tcc) 5 | SCoT 5 | CAATGGCTACCACTAGCG |
| 5 | OPA18 | AGGTGACCGT | ISSR-5 | tgta (ca) 7 | SCoT 6 | CAATGGCTACCACTACAG |
| 6 | OPB5 | TGCGCCCTTC | ISSR-6 | tac (ca) 7 | SCoT 9 | ACAATGGCTACCACTGCC |
| 7 | OPC4 | CCGCATCTAC | ISSR-7 | (ag) 8 t | SCoT 11 | ACAATGGCTACCACTACC |
| 8 | OPC5 | GATGACCGCC | ISSR-8 | cgtc (ac) 7 | SCoT 12 | CAACAATGGCTACCACCG |
| 9 | OPC8 | TGGACCGGTG | ISSR-9 | tcga (ca) 7 | SCoT 13 | ACCATGGCTACCACGGCA |
| 10 | OPD5 | TGAGCGGACA | ISSR-10 | (ag) 8 ct | SCoT 14 | ACCATGGCTACCAGCGCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khayri, J.M.; Mahdy, E.M.B.; Taha, H.S.A.; Eldomiaty, A.S.; Abd-Elfattah, M.A.; Abdel Latef, A.A.H.; Rezk, A.A.; Shehata, W.F.; Almaghasla, M.I.; Shalaby, T.A.; et al. Genetic and Morphological Diversity Assessment of Five Kalanchoe Genotypes by SCoT, ISSR and RAPD-PCR Markers. Plants 2022, 11, 1722. https://doi.org/10.3390/plants11131722
Al-Khayri JM, Mahdy EMB, Taha HSA, Eldomiaty AS, Abd-Elfattah MA, Abdel Latef AAH, Rezk AA, Shehata WF, Almaghasla MI, Shalaby TA, et al. Genetic and Morphological Diversity Assessment of Five Kalanchoe Genotypes by SCoT, ISSR and RAPD-PCR Markers. Plants. 2022; 11(13):1722. https://doi.org/10.3390/plants11131722
Chicago/Turabian StyleAl-Khayri, Jameel M., Ehab M. B. Mahdy, Heba S. A. Taha, Ahmed S. Eldomiaty, Mohamed A. Abd-Elfattah, Arafat Abdel Hamed Abdel Latef, Adel A. Rezk, Wael F. Shehata, Mustafa I. Almaghasla, Tarek A. Shalaby, and et al. 2022. "Genetic and Morphological Diversity Assessment of Five Kalanchoe Genotypes by SCoT, ISSR and RAPD-PCR Markers" Plants 11, no. 13: 1722. https://doi.org/10.3390/plants11131722