
Foliar Functional Traits of Resource Island-Forming Nurse Tree Species from a Semi-Arid Ecosystem of La Guajira, Colombia


Abstract
:1. Introduction
2. Results
2.1. Physiological Traits in Leaves
2.2. Foliar Biochemical Traits
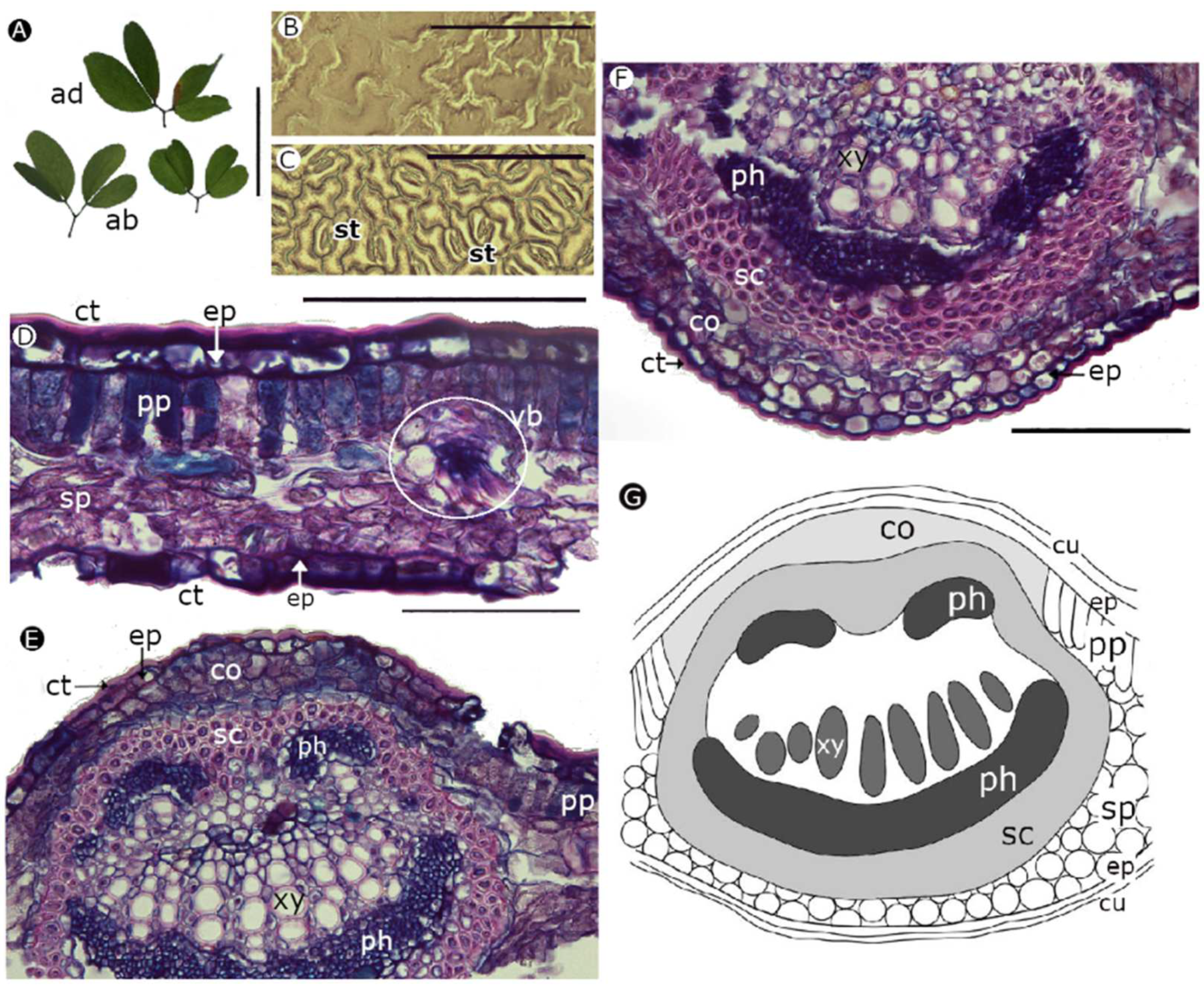
2.3. Foliar Anatomical Traits
3. Discussion
4. Materials and Methods
4.1. Study Area and Plant Material
4.2. Physiological Traits
4.3. Biochemical Traits
4.3.1. Photosynthetic Pigments
4.3.2. Proline Content
4.3.3. Total Sugar Content
4.4. Anatomical Characteristics
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Gu, D.; Wang, Q.; Mallik, A. Non-convergent transpiration and stomatal conductance response of a dominant desert species in central Asia to climate drivers at leaf, branch and whole plant scales. J. Agric. Meteorol. 2018, 74, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Pereira de Oliveira, A.C.; Nunes, A.; Garcia Rodrigues, R.; Branquinho, C. The response of plant functional traits to aridity in a tropical dry forest. Sci. Total Environ. 2020, 747, 141177. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Liu, T.; Duan, L.; Tong, X.; Ji, H.; Zhang, L.; Singh, V.P. A comparative study of three stomatal conductance models for estimating evapotranspiration in a dune ecosystem in a semi-arid region. Sci. Total Environ. 2022, 802, 149937. [Google Scholar] [CrossRef]
- le Polain de Waroux, Y.; Lambin, E.F. Monitoring degradation in arid and semi-arid forests and woodlands: The case of the argan woodlands (Morocco). Appl. Geogr. 2012, 32, 777–786. [Google Scholar] [CrossRef]
- Figueiredo-Lima, K.V.; Falcão, H.M.; Melo-de-Pinna, G.F.; Albacete, A.; Dodd, I.C.; Lima, A.L.; Santos, M.G. Leaf phytohormone levels and stomatal control in an evergreen woody species under semiarid environment in a Brazilian seasonally dry tropical forest. Plant Growth Regul. 2018, 85, 437–445. [Google Scholar] [CrossRef]
- Aponte, M. Organización espacial de la región geográfica de la Alta Guajira colombiana. Perpectiva Geográfica 2010, 15, 157–176. [Google Scholar] [CrossRef]
- Jaramillo, C.; Sepulchre, P.; Cardenas, D.; Correa-Metrio, A.; Moreno, J.E.; Trejos, R.; Vallejos, D.; Hoyos, N.; Martínez, C.; Carvalho, D.; et al. Drastic Vegetation Change in the Guajira Peninsula (Colombia) During the Neogene. Paleoceanogr. Paleoclimatol. 2020, 35, 3933. [Google Scholar] [CrossRef]
- Araujo-carrillo, G.A.; Martínez-maldonado, F.E.; Deantonio-Florido, L.Y.; Gómez-Latorre, D.A. Agroclimatic zoning: A planning strategy for agricultural and livestock systems in Alta Guajira, Colombia Zonificación agroclimática: Una estrategia de planificación para sistemas agrícolas y pecuarios en la Alta Guajira, Colombia. Rev. Colomb. Cienc. Hortícolas 2019, 13, 435–447. [Google Scholar] [CrossRef]
- Bonanomi, G.; Rietkerk, M.; Dekker, S.C.; Mazzoleni, S. Islands of fertility induce co-occurring negative and positive plant-soil feedbacks promoting coexistence. Plant Ecol. 2008, 197, 207–218. [Google Scholar] [CrossRef]
- Celaya, H.; Castellanos, A.E. Nitrogen Mineralization on Arid and Semi-Arid Land Soil. Terra Latinoam. 2011, 29, 343–356. Available online: http://www.scielo.org.mx/pdf/tl/v29n3/2395-8030-tl-29-03-00343.pdf%0Ahttp://www.cabdirect.org/abstracts/20123049100.html (accessed on 27 August 2021).
- Moncayo-Riascos, M.C.; Gálvez-Cerón, A. Islas de fertilidad: Una revisión sistemática de su estructura y operación. Idesia 2018, 36, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Lozano, Y.M.; Hortal, S.; Armas, C.; Pugnaire, F.I. Complementarity in nurse plant systems: Soil drives community composition while microclimate enhances productivity and diversity. Plant Soil 2020, 450, 385–396. [Google Scholar] [CrossRef]
- Salazar, P.C.; Navarro-Cerrillo, R.M.; Grados, N.; Cruz, G.; Barrón, V.; Villar, R. Tree size and leaf traits determine the fertility island effect in Prosopis pallida dryland forest in Northern Peru. Plant Soil 2019, 437, 117–135. [Google Scholar] [CrossRef]
- Feyera, S.; Beck, E.; Lüttge, U. Exotic trees as nurse-trees for the regeneration of natural tropical forests. Trees-Struct. Funct. 2002, 16, 245–249. [Google Scholar] [CrossRef]
- Galindo, V.; Calle, Z.; Chará, J.; Armbrecht, I. Facilitation by pioneer shrubs for the ecological restoration of riparian forests in the Central Andes of Colombia. Restor. Ecol. 2017, 25, 731–737. [Google Scholar] [CrossRef]
- Gómez-Ruiz, P.A.; Lindig-Cisneros, R.; Vargas-Ríos, O. Facilitation among plants: A strategy for the ecological restoration of the high-andean forest (Bogotá, D.C.-Colombia). Ecol. Eng. 2013, 57, 267–275. [Google Scholar] [CrossRef]
- Guerra, A.; Scremin-Dias, E. Leaf traits, sclerophylly and growth habits in plant species of a semiarid environment. Rev. Bras. Bot. 2018, 41, 131–144. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Xu, L.; Chen, Z.; He, N. Variation in leaf morphological, stomatal, and anatomical traits and their relationships in temperate and subtropical forests. Sci. Rep. 2019, 9, 5803. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, L.H.; Oliveira, B.F.; Krah, F.S.; Kollmann, J.; Ganade, G. Linking plant traits to multiple soil functions in semi-arid ecosystems. J. Arid Environ. 2020, 172, 104040. [Google Scholar] [CrossRef]
- Salgado-Negret, B.; Pulido Rodríguez, E.N.; Cabrera, M.; Ruíz Osorio, C.; Paz, H. Protocolo para la medición de rasgos fucnionales en plantas. In La Ecología Funcional como Aproximación al Estudio, Manejo y Conservación de la Biodiversidad: Protocolos y Aplicaciones; Salgado-Negret, B., Ed.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2015; pp. 37–79. [Google Scholar]
- Salgado-Negret, B.; Canessa, R.; Valladares, F.; Armesto, J.J.; Pérez, F. Functional traits variation explains the distribution of aextoxicon punctatum(Aextoxicaceae) in pronounced moisture gradients within fog-dependent forest fragments. Front. Plant Sci. 2015, 6, 511. [Google Scholar] [CrossRef] [Green Version]
- Lima e Silva, Á.M.; de Faria Lopes, S.; Porto Vitorio, L.A.; Santiago, R.R.; de Mattos, E.A.; de Brito Melo Trovão, D.M. Plant functional groups of species in semiarid ecosystems in Brazil: Wood basic density and SLA as an ecological indicator. Rev. Bras. Bot. 2014, 37, 229–237. [Google Scholar] [CrossRef]
- Chaves, M.M.; Costa, J.M.; Zarrouk, O.; Pinheiro, C.; Lopes, C.M.; Pereira, J.S. Controlling stomatal aperture in semi-arid regions—The dilemma of saving water or being cool? Plant Sci. 2016, 251, 54–64. [Google Scholar] [CrossRef] [PubMed]
- De Souza, B.C.; Oliveira, R.S.; De Araújo, F.S.; De Lima, A.L.A.; Rodal, M.J.N. Divergências funcionais e estratégias de resistência à seca entre espécies decíduas e sempre verdes tropicais. Rodriguesia 2015, 66, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- González Moreno, S.; Perales, H.; Salcedo, M. la Fluorescencia de la Clorofila a Como Herramienta en la Investigación de Efectos Tóxicos en el Aparato Fotosintético de Plantas y Algas. Rev. Educ. Bioquímica 2008, 27, 119–129. Available online: https://www.redalyc.org/pdf/490/49011464003.pdf (accessed on 19 April 2021).
- Kumar, D.; Singh, H.; Raj, S.; Soni, V. Chlorophyll a fluorescence kinetics of mung bean (Vigna radiata L.) grown under artificial continuous light. Biochem. Biophys. Rep. 2020, 24, 100813. [Google Scholar] [CrossRef] [PubMed]
- Mihaljević, I.; Viljevac Vuletić, M.; Šimić, D.; Tomaš, V.; Horvat, D.; Josipović, M.; Zdunić, Z.; Dugalić, K.; Vuković, D. Comparative Study of Drought Stress Effects on Traditional and Modern Apple Cultivars. Plants 2021, 10, 561. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 443–480. Available online: http://www.hansatech-instruments.com/docs/thefluorescencetransient.pdf (accessed on 27 September 2020).
- Strasser, R.J.; Tsimilli- Michael, M.; Srivastava, A. Analysis of the Fluorescence Transient Merope. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 1–47. [Google Scholar] [CrossRef]
- Moreno, L.; Crespo, S.C.; Pérez, W.; Melgarejo, L.M. Pruebas bioquímicas como herramientas para estudios en fisiología. In Experimentos en Fisiología Vegetal; Melgarejo, L.M., Ed.; Universidad Nacional de Colombia: Bogotá, Colombia, 2010; pp. 187–248. Available online: http://ciencias.bogota.unal.edu.co/fileadmin/Facultad_de_Ciencias/Publicaciones/Imagenes/Portadas_Libros/Biologia/Experimentos_en_fisiologia_Vegetal/ExperimentosEnFisiologiaVegetal.pdf (accessed on 27 September 2020).
- Crespo, S.C.; Moreno-Chacón, A.L.; Rojas, A.; Melgarejo, L.M. Principal component analysis of changes due to water stress for some osmolytes, pigments and antioxidant enzymes in Gmelina arborea Robx. Leaves from trees planted in northern Colombia. J. Braz. Chem. Soc. 2011, 22, 2275–2280. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.; Medeiros, M.; Pereira, S.; Oliveira, M.; Frosi, G.; Arruda, E.; Santos, M. Ecophysiological leaf traits of native and exotic palm tree species under semi-arid conditions. Bragantia 2016, 75, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-López, A.; Yahia, E.M. Morphology and anatomy. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Carrillo-López, A., Eds.; Elsevier: Sawston, UK, 2018; pp. 113–130. [Google Scholar]
- Lopez, F.B.; Barclay, G.F. Plant Anatomy and Physiology. In Pharmacognosy: Fundamentals, Applications and Strategy; Badal, S., Delgoda, R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 45–60. [Google Scholar] [CrossRef]
- Males, J.; Griffiths, H. Stomatal biology of CAM plants. Plant Physiol. 2017, 174, 550–560. [Google Scholar] [CrossRef] [Green Version]
- Fradera-Soler, M.; Rudall, P.J.; Prychid, C.J.; Grace, O.M. Evolutionary success in arid habitats: Morpho-anatomy of succulent leaves of Crassula species from southern Africa. J. Arid Environ. 2021, 185, 104319. [Google Scholar] [CrossRef]
- Priscila, M.; Lima, D.S.; Soares, A.; Marcelo, J.; Porto, P.; Sá, F.S. Leaf Anatomy of Rubiaceae Species in a Semiarid Area of Brazil. Rodriguésia 2020, 71, 1096. [Google Scholar] [CrossRef]
- Rosado Vega, J.; Díaz Chávez, L.A.; Rosado Vega, J.; Fernández Moreno, M.I. Palos, Yerbas y Bejucos de la Guajira: Bosque Seco; Universidad de La Guajira: Riohacha, Colombia, 2018; Available online: http://174.142.98.156/shop/catalogs/show_material_details/73833 (accessed on 19 October 2019).
- Martinez, S. Monitoreo Participativo de Flora Presente en Bosque Seco de los Departamentos de Bolivar, Huila y La Guajira; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2019; Available online: http://i2d.humboldt.org.co/ceiba/resource.do?r=rrbb_plantas_cienciaparticipatva_bs_2019 (accessed on 19 October 2019).
- Rosado, J.; Moreno, M. Farmacopea guajira: El uso de las plantas medicinales xerofíticas por la etnia wayuu. Rev. CENIC Cienc. Biológicas 2010, 41, 1–10. Available online: https://www.redalyc.org/articulo.oa?id=181220509064 (accessed on 19 October 2019).
- Rangel-Ch, J.O. La vegetación de la región caribe de Colombia: Composición florística y aspectos de la estructura. In Colombia Diversidad Biótica XII: La Región Caribe de Colombia; Rangel-Ch, J.O., Ed.; Instituto de Ciencias Naturales, Universidad Nacional de Colombia: Bogotá, Colombia, 2012; pp. 365–476. Available online: http://ciencias.bogota.unal.edu.co/menu-principal/publicaciones/biblioteca-digital/colombia-diversidad-biotica/ (accessed on 19 October 2019).
- Chaturvedi, R.K.; Raghubanshi, A.S.; Singh, J.S. Plant functional traits with particular reference to tropical deciduous forests: A review. J. Biosci. 2011, 36, 963–981. [Google Scholar] [CrossRef]
- Cousins, A.B.; Mullendore, D.L.; Sonawane, B.V. Recent developments in mesophyll conductance in C3, C4, and crassulacean acid metabolism plants. Plant J. 2020, 101, 816–830. [Google Scholar] [CrossRef] [Green Version]
- Edwards, E.J.; Diaz, M. Ecological physiology of Pereskia guamacho, a cactus with leaves. Plant Cell Environ. 2006, 29, 247–256. [Google Scholar] [CrossRef]
- Edwards, E.J.; Donoghue, M.J. Pereskia and the origin of the cactus life-form. Am. Nat. 2006, 167, 777–793. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, R.K.; Tripathi, A.; Raghubanshi, A.S.; Singh, J.S. Functional traits indicate a continuum of tree drought strategies across a soil water availability gradient in a tropical dry forest. For. Ecol. Manag. 2021, 482, 118740. [Google Scholar] [CrossRef]
- Vendramini, F.; Díaz, S.; Gurvich, D.E.; Wilson, P.J.; Thompson, K.; Hodgson, J.G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytol. 2002, 154, 147–157. [Google Scholar] [CrossRef]
- Liu, S.; Yan, Z.; Chen, Y.; Zhang, M.; Chen, J.; Han, W. Foliar pH, an emerging plant functional trait: Biogeography and variability across northern China. Glob. Ecol. Biogeogr. 2019, 28, 386–397. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Quested, H.M.; Van Logtestijn, R.S.P.; Pérez-Harguindeguy, N.; Gwynn-Jones, D.; Díaz, S.; Callaghan, T.V.; Press, M.C.; Aerts, R. Foliar pH as a new plant trait: Can it explain variation in foliar chemistry and carbon cycling processes among subarctic plant species and types? Oecologia 2006, 147, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Mendes, K.R.; Granja, J.A.A.; Ometto, J.P.; Antonino, A.C.D.; Menezes, R.S.C.; Pereira, E.C.; Pompelli, M.F. Croton blanchetianus modulates its morphophysiological responses to tolerate drought in a tropical dry forest. Funct. Plant Biol. 2017, 44, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- de Sousa Mendes, M.M.; de Lacerda, C.F.; Fernandes, F.É.P.; Cavalcante, A.C.R.; de Oliveira, T.S. Ecophysiology of deciduous plants grown at different densities in the semiarid region of Brazil. Theor. Exp. Plant Physiol. 2013, 25, 94–105. Available online: https://www.semanticscholar.org/paper/Ecophysiology-of-deciduous-plants-grown-at-in-the-Mendes-Lacerda/2860468556880781337dc93ea7f98b61e241a206 (accessed on 27 August 2021). [CrossRef] [Green Version]
- Chapin, F.S.; Matson, P.A.; Vitousek, P.M. Principles of Terrestrial Ecosystem Ecology; Chapin, F.S., Matson, P.A., Vitousek, P.M., Eds.; Springer International Publishing: New York, NY, USA, 2011; 536p, p. 536. [Google Scholar] [CrossRef]
- Garcia-Forner, N.; Adams, H.D.; Sevanto, S.; Collins, A.D.; Dickman, L.T.; Hudson, P.J.; Zeppel, M.J.B.; Jenkins, M.W.; Powers, H.; Martínez-Vilalta, J.; et al. Responses of two semiarid conifer tree species to reduced precipitation and warming reveal new perspectives for stomatal regulation. Plant Cell Environ. 2016, 39, 38–49. [Google Scholar] [CrossRef] [Green Version]
- De Micco, V.; Aronne, G. Morpho-anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 37–61. [Google Scholar] [CrossRef]
- Yücedağ, C.; Sanders, J.; Musah, M.; Gailing, O. Stomatal density in Quercus petraea and Q. Robur natural populations in northern Turkey. Dendrobiology 2019, 81, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Sundberg, M.D. A comparison of stomatal distribution and length in succulent and non- succulent desert plants. Phytomorphology 1986, 36, 53–66. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201301401900 (accessed on 19 April 2021).
- Aragón-Gastélum, J.L.; Ramírez-Benítez, J.E.; González-Durán, E.; González-Salvatierra, C.; Ramírez-Tobías, H.M.; Flores, J.; Gutiérrez-Alcántara, E.J.; Méndez-Guzmán, E.; Jarquín-Gálvez, R. Photochemical activity in early-developmental phases of Agave angustifolia subsp. tequilana under induced global warming: Implications to temperature stress and tolerance. Flora Morphol. Distrib. Funct. Ecol. Plants 2020, 263, 151535. [Google Scholar] [CrossRef]
- da Jardim, A.M.R.F.; Santos, H.R.B.; Alves, H.K.M.N.; Ferreira-Silva, S.L.; de Souza, L.S.B.; do Araújo Júnior, G.N.; de Souza, M.S.; de Araújo, G.G.L.; de Souza, C.A.A.; da Silva, T.G.F. Genotypic differences relative photochemical activity, inorganic and organic solutes and yield performance in clones of the forage cactus under semi-arid environment. Plant Physiol. Biochem. 2021, 162, 421–430. [Google Scholar] [CrossRef]
- Liu, Y.; Lei, S.G.; Chen, X.Y.; Chen, M.; Zhang, X.Y.; Long, L.L. Disturbance mechanism of coal mining subsidence to typical plants in a semiarid area using O–J–I–P chlorophyll a fluorescence analysis. Photosynthetica 2020, 58, 1178–1187. [Google Scholar] [CrossRef]
- Ayyaz, A.; Amir, M.; Umer, S.; Iqbal, M.; Bano, H.; Gul, H.S.; Noor, Y.; Kanwal, A.; Khalid, A.; Javed, M.; et al. Melatonin induced changes in photosynthetic efficiency as probed by OJIP associated with improved chromium stress tolerance in canola (Brassica napus L.). Heliyon 2020, 6, e04364. [Google Scholar] [CrossRef]
- Li, L.; Li, X.Y.; Xu, X.W.; Lin, L.S.; Zeng, F.J. Effects of high temperature on the chlorophyll a fluorescence of Alhagi sparsifolia at the southern Taklamakan Desert. Acta Physiol. Plant. 2014, 36, 243–249. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Gonçalves, J.F.; Dos Santos, U.M. Utilization of the chlorophyll a fluorescence technique as a tool for selecting tolerant species to environments of high irradiance. Braz. J. Plant Physiol. 2005, 5, 307–313. [Google Scholar] [CrossRef]
- Sitko, K.; Rusinowski, S.; Kalaji, H.M.; Szopiński, M.; Małkowski, E. Photosynthetic efficiency as bioindicator of environmental pressure in A. halleri. Plant Physiol. 2017, 175, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, H.; Chen, J.M.; Wang, R.; Mo, G.; Luo, S.; Luo, X.; He, L.; Gonsamo, A.; Arabian, J.; Zhang, Y.; et al. The global distribution of leaf chlorophyll content. Remote Sens. Environ. 2020, 236, 111479. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.; Chen, J.M.; Croft, H.; Luo, X.; Zheng, T.; Rogers, C.; Liu, J. Fine-scale leaf chlorophyll distribution across a deciduous forest through two-step model inversion from Sentinel-2 data. Remote Sens. Environ. 2021, 264, 112618. [Google Scholar] [CrossRef]
- Rivas, R.; Barros, V.; Falcão, H.; Frosi, G.; Arruda, E.; Santos, M. Ecophysiological Traits of Invasive C3 Species Calotropis procera to Maintain High Photosynthetic Performance Under High VPD and Low Soil Water Balance in Semi-Arid and Seacoast Zones. Front. Plant Sci. 2020, 11, 717. [Google Scholar] [CrossRef]
- Rong-Hua, L.; Pei-guo, G.; Baum, M.; Grando, S.; Ceccarelli, S. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Baunthiyal, M.; Sharma, V. Response of Three Semi-Arid Plant Species to Fluoride; Consequences for Chlorophyll Florescence. Int. J. Phytoremediation 2014, 16, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Koźmińska, A.; Al Hassan, M.; Wiszniewska, A.; Hanus-Fajerska, E.; Boscaiu, M.; Vicente, O. Responses of succulents to drought: Comparative analysis of four Sedum (Crassulaceae) species. Sci. Hortic. 2019, 243, 235–242. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Roldán, A.; Azcón, R. The effectiveness of arbuscular-mycorrhizal fungi and Aspergillus niger or Phanerochaete chrysosporium treated organic amendments from olive residues upon plant growth in a semi-arid degraded soil. J. Environ. Manag. 2010, 91, 2547–2553. [Google Scholar] [CrossRef]
- Lersten, N.R.; Curtis, J.D. Survey of leaf anatomy, especially secretory structures, of tribe Caesalpinieae (Leguminosae, Caesalpinioideae). Plant Syst. Evol. 1996, 200, 21–39. [Google Scholar] [CrossRef]
- Martel, C.; Rojas, N.; Marín, M.; Avilés, R.; Neira, E.; Santiago, J. Caesalpinia spinosa (Caesalpiniaceae) leaves: Anatomy, histochemistry, and secondary metabolites. Rev. Bras. Bot. 2014, 37, 167–174. [Google Scholar] [CrossRef]
- García, M.; Torrecilla, P. Anatomía foliar de algunos taxones representativos de las secciones del género Pithecellobium Martius s. str. (Leguminosae: Mimosoideae: Ingeae en Venezuela). Ernstia 1999, 9, 1–18. [Google Scholar]
- Alvarado, G.; García, M.; Jáuregui, D.; Alvarado, H.; Zambrano, F. Leaf anatomy of six arboreal eudicotyledons species growing along an altitudinal gradient on the high basin of the Tocuyo river, Venezuela. Biologia 2020, 75, 523–533. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Correia, C.M.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Lopes, J.I.; Torres-Pereira, J.M.G. Sclerophylly and leaf anatomical traits of five field-grown olive cultivars growing under drought conditions. Tree Physiol. 2004, 24, 233–239. [Google Scholar] [CrossRef]
- Medina, E.; Garcia, V.; Cuevas, E. Sclerophylly and Oligotrophic Environments: Relationships Between Leaf Structure, Mineral Nutrient Content, and Drought Resistance in Tropical Rain Forests of the Upper Rio Negro Region. Biotropica 1990, 22, 51. [Google Scholar] [CrossRef]
- Da Penha Xavier, L.; Pereira de Arruda, E.C. Leaf anatomy of Senna cana (Fabaceae) in a seasonally dry tropical forest. Rev. Caatinga 2021, 34, 155–165. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, I.W. Comparative anatomy of the leaf–bearing Cactaceae, I. Foliar vasculature of Pereskia, Pereskiopsis and Quiabentia. J. Arnold Arbor. 1960, 41, 341–356. Available online: https://www.jstor.org/stable/43782467?read-now=1#page_scan_tab_contents (accessed on 14 May 2021). [CrossRef]
- Mauseth, J.D.; Landrum, J.V. Relictual vegetative anatomical characters in Cactaceae: The Genus Pereskia. J. Plant Res. 1997, 110, 55–64. [Google Scholar] [CrossRef]
- Duarte, M.R.; Hayashi, S.S. Estudo anatômico de folha e caule de Pereskia aculeata Mill. (Cactaceae). Rev. Bras. Farmacogn. 2005, 15, 103–109. [Google Scholar] [CrossRef]
- Maciel, V.B.V.; Yoshida, C.M.P.; Goycoolea, F.M. Agronomic Cultivation, Chemical Composition, Functional Activities and Applications of Pereskia Species—A Mini Review. Curr. Med. Chem. 2018, 26, 4573–4584. [Google Scholar] [CrossRef]
- Karwowska, K.; Brzezicka, E.; Kozieradzka-Kiszkurno, M.; Chernetskyy, M. Anatomical structure of the leaves of Crassula cordata (Crassulaceae). Mod. Phytomorphol. 2015, 8, 53–54. [Google Scholar] [CrossRef]
- Ogburn, R.M.; Edwards, E.J. Anatomical variation in Cactaceae and relatives: Trait lability and evolutionary innovation. Am. J. Bot. 2009, 96, 391–408. [Google Scholar] [CrossRef] [Green Version]
- Bailey, W. Comparative anatomy of the leaf-bearing Cactaceae, III: Form and distribution of crustals in Pereskia, Pereskiopsis and Quiabentia. J. Arnold Arbor. 1961, 42, 334–346. Available online: https://www.jstor.org/stable/43781392 (accessed on 27 August 2021). [CrossRef]
- Gómez-Espinoza, O.; González-Ramírez, D.; Méndez-Gómez, J.; Guillén-Watson, R.; Medaglia-Mata, A.; Bravo, L.A. Calcium oxalate crystals in leaves of the extremophile plant Colobanthus quitensis (Kunth) bartl. (caryophyllaceae). Plants 2021, 10, 1787. [Google Scholar] [CrossRef]
- Warwick, N.W.M.; Hailey, L.; Clarke, K.L.; Gasson, P.E. Climate trends in the wood anatomy of Acacia sensu stricto (Leguminosae: Mimosoideae). Ann. Bot. 2017, 119, 1249–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, S.; Dang, Q.L.; Man, R.; Tedla, B. Northward migration of trembling aspen will increase growth but reduce resistance to drought-induced xylem cavitation. Botany 2019, 97, 627–638. [Google Scholar] [CrossRef]
- Pérez, L.V.; Rojas, Y.A.; Melgarejo, L.M. Agua. In Experimentos en Fisiología Vegetal; Melgarejo, L.M., Ed.; Universidad Nacional de Colombia: Bogotá, Colombia, 2010; pp. 63–78. Available online: http://ciencias.bogota.unal.edu.co/fileadmin/Facultad_de_Ciencias/Publicaciones/Imagenes/Portadas_Libros/Biologia/Experimentos_en_fisiologia_Vegetal/ExperimentosEnFisiologiaVegetal.pdf (accessed on 27 September 2020).
- Suárez Salazar, J.C.; Melgarejo, L.M.; Casanoves, F.; Di Rienzo, J.A.; DaMatta, F.M.; Armas, C. Photosynthesis limitations in cacao leaves under different agroforestry systems in the Colombian Amazon. PLoS ONE 2018, 13, e0206149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Oren, R.; Hinckley, T.M. Actual and potential transpiration and carbon assimilation in an irrigated poplar plantation. Tree Physiol. 2008, 28, 559–577. [Google Scholar] [CrossRef] [Green Version]
- Brestic, M.; Zivcak, M. PSII fluorescence techniques for measurements of drought and high temperature stress signal of crop plants: Protocols and applications. Mol. Stress Physiol. Plants 2013, 87–131. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Ganesan, M.; Strasser, R.J.; Han, Y.; Kim, J.I.; Lee, H.Y.; Song, P.S. In Vivo assessment of cold tolerance through chlorophyll-a fluorescence in transgenic zoysiagrass expressing mutant phytochrome A. PLoS ONE 2015, 10, e0127200. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. Available online: https://www.sciencedirect.com/science/article/abs/pii/0076687987480361 (accessed on 27 September 2020).
- Solarte, M.E.; Moreno, L.; Melgarejo, L.M. Fotosíntesis y pigmentos vegetales. In Experimentos en Fisiología Vegetal; Melgarejo, L.M., Ed.; Universidad Nacional de Colombia: Bogotá, Colombia, 2010; Volume 1, pp. 107–122. Available online: http://ciencias.bogota.unal.edu.co/fileadmin/Facultad_de_Ciencias/Publicaciones/Imagenes/Portadas_Libros/Biologia/Experimentos_en_fisiologia_Vegetal/ExperimentosEnFisiologiaVegetal.pdf (accessed on 27 September 2020).
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef]
- Johansen, D.A. Plan Microtechnique; McGraw-Hill: New York, NY, USA, 1940; Available online: https://www.worldcat.org/title/plant-microtechnique/oclc/399073 (accessed on 20 July 2021).
- Robles, A.; Raz, L.; Marquínez, X. Anatomía floral de Peristethium leptostachyum (Loranthaceae). Rev. Biol. Trop. 2016, 64, 341–352. Available online: https://www.scielo.sa.cr/scielo.php?pid=S0034-77442016000100341&script=sci_abstract&tlng=es (accessed on 20 July 2021). [CrossRef] [Green Version]
- Barclay, R.; McElwain, J.; Dilcher, D.; Sageman, B. The cuticle database: Developing an interactive tool for taxonomic and paleoenvironmental study of the fossil cuticle record. CFS Cour. Forsch. Senckenb. 2007, 258, 39–55. Available online: https://www.scholars.northwestern.edu/en/publications/the-cuticle-database-developing-an-interactive-tool-for-taxonomic (accessed on 20 July 2021).
- Trofimov, D.; Rohwer, J.G. Epidermal features allowing identification of evolutionary lineages in the Ocotea complex (Lauraceae). Perspect. Plant Ecol. Evol. Syst. 2018, 31, 17–35. [Google Scholar] [CrossRef]
- Dave, Y.S.; Patel, N.D. Structure of Stomatal Complexes in Pedilanthus tithymaloidesPOIT. III. Flora 1976, 165, 235–241. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0367253017318522 (accessed on 20 July 2021). [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | H. brasiletto | P. dulce | P. guamacho | Group |
|---|---|---|---|---|
| WC (%) | 151.79 ± 10.1 | 121.43 ± 34.87 | 744.59 ± 87.76 | (a, b, c) |
| RWC (%) | 83.46 ± 10.48 | 90.85 ± 4.21 | 97.78 ± 0.61 | (a, b, c) |
| WSD (%) | 16.54 ± 10.49 | 9.15 ± 4.21 | 2.22 ± 0.6 | (a, b, c) |
| LA (cm2) | 3.67 ± 1.56 | 6.91 ± 3.7 | 7.39 ± 1.96 | (a, b, b) |
| SLA (cm2/g) | 134.19 ± 10.88 | 108.88 ± 23.82 | 113.58 ± 38.79 | (a, b, b) |
| LDMC (mg/g) | 364.8 ± 19.97 | 437.47 ± 21.63 | 115.38 ± 13.28 | (a, b, c) |
| Thickness (mm) | 0.2 ± 0.03 | 0.23 ± 0.05 | 0.81 ± 0.34 | (a, b, c) |
| pH | 5.17 ± 0.27 | 5.72 ± 0.08 | 5.24 ± 0.32 | (a, b, a) |
| SD (stomata/mm2) | 223.53 ± 55.88 | 647.06 ± 164.7 | 38.06 ± 6.36 | (a, b, c) |
| gs 3:30–5:30 h (mmol·m−2·s−1) | 64.8 ± 20 | 60.6 ± 20.7 | 50.8 ± 24.3 | (a, a, a) |
| gs 12:30–15:00 h (mmol·m−2·s−1) | 45.3 ± 16.55 | 87.6 ± 14.9 | 22 ± 3.3 | (a, b, c) |
| gs 17:30–19:30 h (mmol·m−2·s−1) | 40.6 ± 5.5 | 39.8 ± 6.6 | 35.9 ± 6.9 | (a, ab, c) |
| Variable | H. brasiletto | P. dulce | P. guamacho | Group |
|---|---|---|---|---|
| PIABS | 5.84 ± 2.19 | 4.42 ± 3.37 | 1.16 ± 0.55 | (a, a, b) |
| ΦPo (Fv/Fm) | 0.84 ± 0.01 | 0.82 ± 0.02 | 0.73 ± 0.09 | (a, a, b) |
| ΦEo | 0.48 ± 0.05 | 0.44 ± 0.09 | 0.3 ± 0.05 | (a, b, c) |
| ΨEo | 0.57 ± 0.07 | 0.53 ± 0.09 | 0.43 ± 0.03 | (a, b, c) |
| ABS/RC | 1.19 ± 0.22 | 1.24 ± 0.34 | 1.86 ± 0.16 | (a, a, b) |
| DIo/RC | 0.19 ± 0.03 | 0.22 ± 0.09 | 0.5 ± 0.22 | (a, a, b) |
| TRo/RC | 1 ± 0.19 | 1.02 ± 0.26 | 1.35 ± 0.12 | (a, a, b) |
| ETo/RC | 0.56 ± 0.05 | 0.53 ± 0.06 | 0.56 ± 0.11 | (ab, b, a) |
| REo/RC | 0.14 ± 0.03 | 0.15 ± 0.03 | 0.17 ± 0.02 | (a, b, b) |
| ABS/CSo | 309 ± 58 | 360 ± 101 | 290 ± 108 | (a, b, a) |
| DIo/CSo | 51.45 ± 9.96 | 64.24 ± 22.97 | 84.74 ± 25.94 | (a, b, c) |
| TRo/CSo | 259.78 ± 48.07 | 296.93 ± 72.13 | 224.88 ± 106.8 | (a, a, b) |
| ETo/CSo | 147.75 ± 31.07 | 159.42 ± 48.64 | 98.85 ± 36.06 | (a, a, b) |
| REo/CSo | 37.27 ± 14.76 | 45.53 ± 13.11 | 27.25 ± 6.87 | (a, b, c) |
| Variable | H. brasiletto | P. dulce | P. guamacho | p-Value | Groups |
|---|---|---|---|---|---|
| Mesophyll (mm) | 0.103 ± 0.003 | 0.092 ± 0.01 | 0.415 ± 0.149 | 6 × 10−8 | (a, b, c) |
| Upper epidermis(mm) | 0.016 ± 0.003 | 0.01 ± 0.002 | 0.022 ± 0.003 | 2 × 10−7 | (a, b, c) |
| Lower epidermis (mm) | 0.012 ± 0.003 | 0.01 ± 0.001 | 0.021 ± 0.005 | 1 × 10−7 | (a, a, b) |
| Palisade parenchyma (mm) | 0.033 ± 0.006 | 0.031 ± 0.013 | - | NS | (a, a, -) |
| Spongy parenchyma (mm) | 0.04 ± 0.006 | 0.035 ± 0.006 | - | 3 × 10−3 | (a, b, -) |
| Undifferentiated parenchyma (mm) | - | - | 0.368 ± 0.139 | - | - |
| Upper cuticle (mm) | - | 0.002 ± 0 | - | - | - |
| Lower cuticle (mm) | - | 0.001 ± 0.001 | - | - | - |
| IDV (mm) | 0.012 ± 0.002 | 0.011 ± 0.001 | 0.011 ± 0.004 | NS | (a, a, a) |
| WWV (mm) | 0.002 ± 0 | 0.002 ± 0 | 0.003 ± 0.001 | 4 × 10−5 | (a, ab, c) |
| LX (mm) | 0.072 ± 0.004 | 0.053 ± 0.007 | 0.049 ± 0.016 | 4 × 10−5 | (a, b, b) |
| Percentage (%) | 27.613 ± 3.517 | 19.203 ± 7.631 | 43.023 ± 12.075 | 4 × 10−5 | (a, b, c) |
| Parameter | Basic Physiological Interpretation | Equation |
|---|---|---|
| Basic parameters derived from OJIP transient | ||
| Fo | Minimum fluorescence, when all RC PS II are open | - |
| Fm | Maximum fluorescence, when all RC PS II are closed | - |
| Fv | Variable fluorescence | Fv = Fm − Fo |
| Vt | Relative variable fluorescence at time t | |
| Vj | Relative variable fluorescence at time 2 ms (J-step) | Fm − Fo) |
| Vi | Relative variable fluorescence at time 30 ms (I-step) | Fm − Fo) |
| TFm | Time of reaching maximum fluorescence | |
| Mo | Initial slope of relative variable chlorophyll fluorescence, express the rate of electron trapping | |
| Quantum yields | ||
| Quantum efficiency of photosystem II | ||
| Probability that an absorbed exciton moves an electron after QA− | ||
| Efficiency with which a trapped exciton can move an electron after QA− | ||
| Efficiency with which an electron from the intersystem electron carriers moves to reduce end electron acceptors at the PSI acceptor side (RE) | ||
| Specific energy fluxes (per reaction center—RC) | ||
| ABS/RC | Absorption (ABS) per RC | |
| DIo/RC | Dissipation (DI) at time 0 per RC | |
| TRo/RC | Trapped energy flux (TR) at time 0 per RC | |
| ETo/RC | Electron transport flux (ET) at time 0 per RC | |
| REo/RC | Reduction in final electron acceptors on the electron acceptor side of PSI (RE) at time 0 per RC | |
| Phenomenological fluxes (per excited cross-section—CS) | ||
| ABS/CSo | Absorption (ABS) per CS using the Fodark value the samples exhibited while in a dark-adapted state | |
| DIo/CSo | DIo/CSo = (ABS/CSo) − (TRo/CSo) | |
| TRo/CSo | Trapping (TR) at time 0 per CS | (ABS/CSo) |
| ETo/CSo | Electron transport (ET) at time 0 per CS | (ABS/CSo) |
| REo/CSo | Reduction in final electron acceptors on the electron acceptor side of PS I (RE) at time 0 per CS | (ABS/CSo) |
| Performance index | ||
| PIABS | Performance index | PIABS = [RC/ABS][φPO/(1 − φ)] ∗ [ψ/1 − ψo] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toro-Tobón, G.; Alvarez-Flórez, F.; Mariño-Blanco, H.D.; Melgarejo, L.M. Foliar Functional Traits of Resource Island-Forming Nurse Tree Species from a Semi-Arid Ecosystem of La Guajira, Colombia. Plants 2022, 11, 1723. https://doi.org/10.3390/plants11131723
Toro-Tobón G, Alvarez-Flórez F, Mariño-Blanco HD, Melgarejo LM. Foliar Functional Traits of Resource Island-Forming Nurse Tree Species from a Semi-Arid Ecosystem of La Guajira, Colombia. Plants. 2022; 11(13):1723. https://doi.org/10.3390/plants11131723
Chicago/Turabian StyleToro-Tobón, Gabriela, Fagua Alvarez-Flórez, Hernán D. Mariño-Blanco, and Luz M. Melgarejo. 2022. "Foliar Functional Traits of Resource Island-Forming Nurse Tree Species from a Semi-Arid Ecosystem of La Guajira, Colombia" Plants 11, no. 13: 1723. https://doi.org/10.3390/plants11131723