
Bouquet Formation Failure in Meiosis of F1 Wheat–Rye Hybrids with Mitotic-Like Division

Abstract
:1. Introduction
2. Results
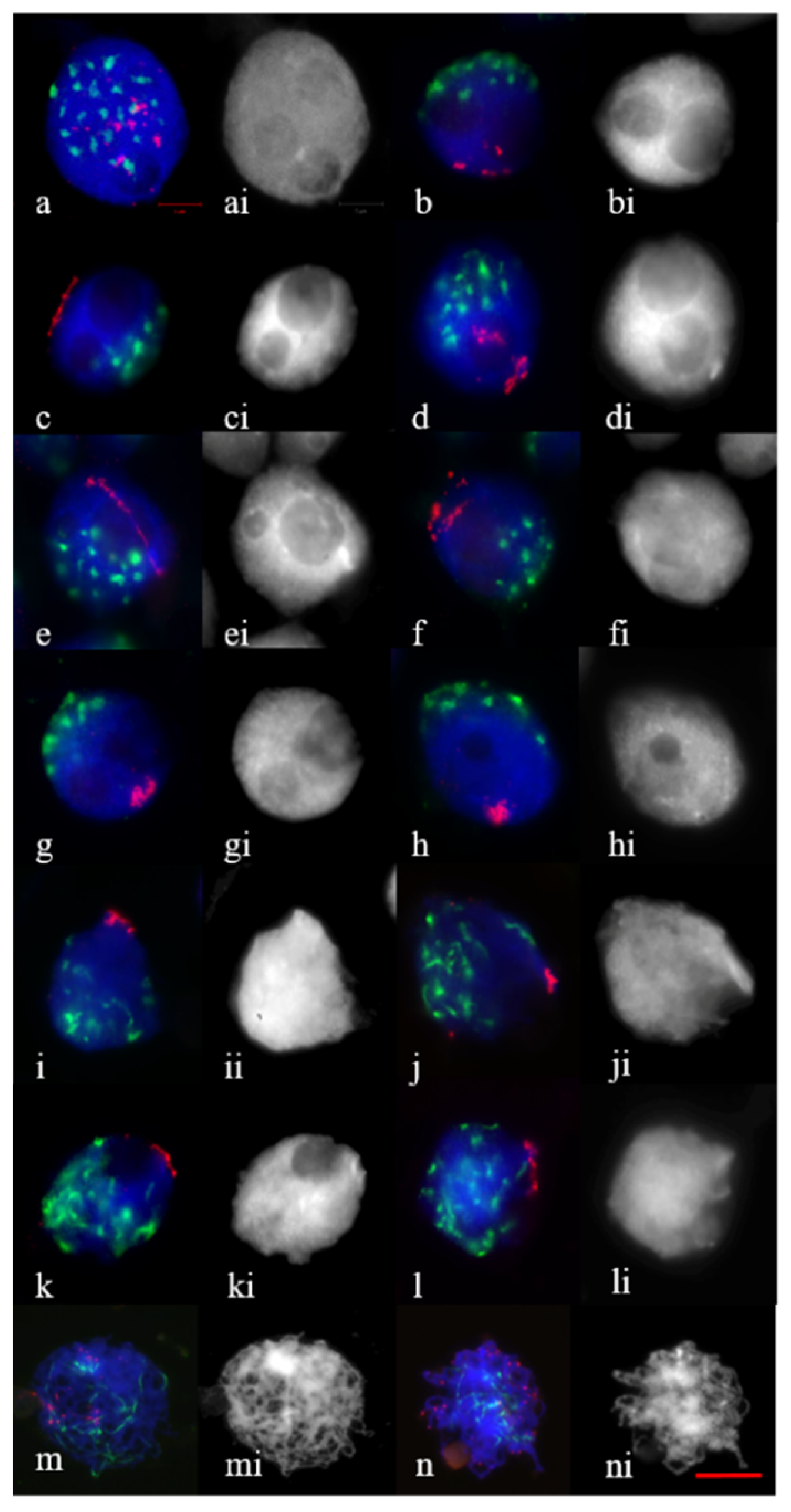
2.1. Telomere and Centromere Dynamics in Prophase in Bread Wheat Var. Saratovskaya 29, and Disomic Substitution Lines 1Rv(1A), 2R(2D)
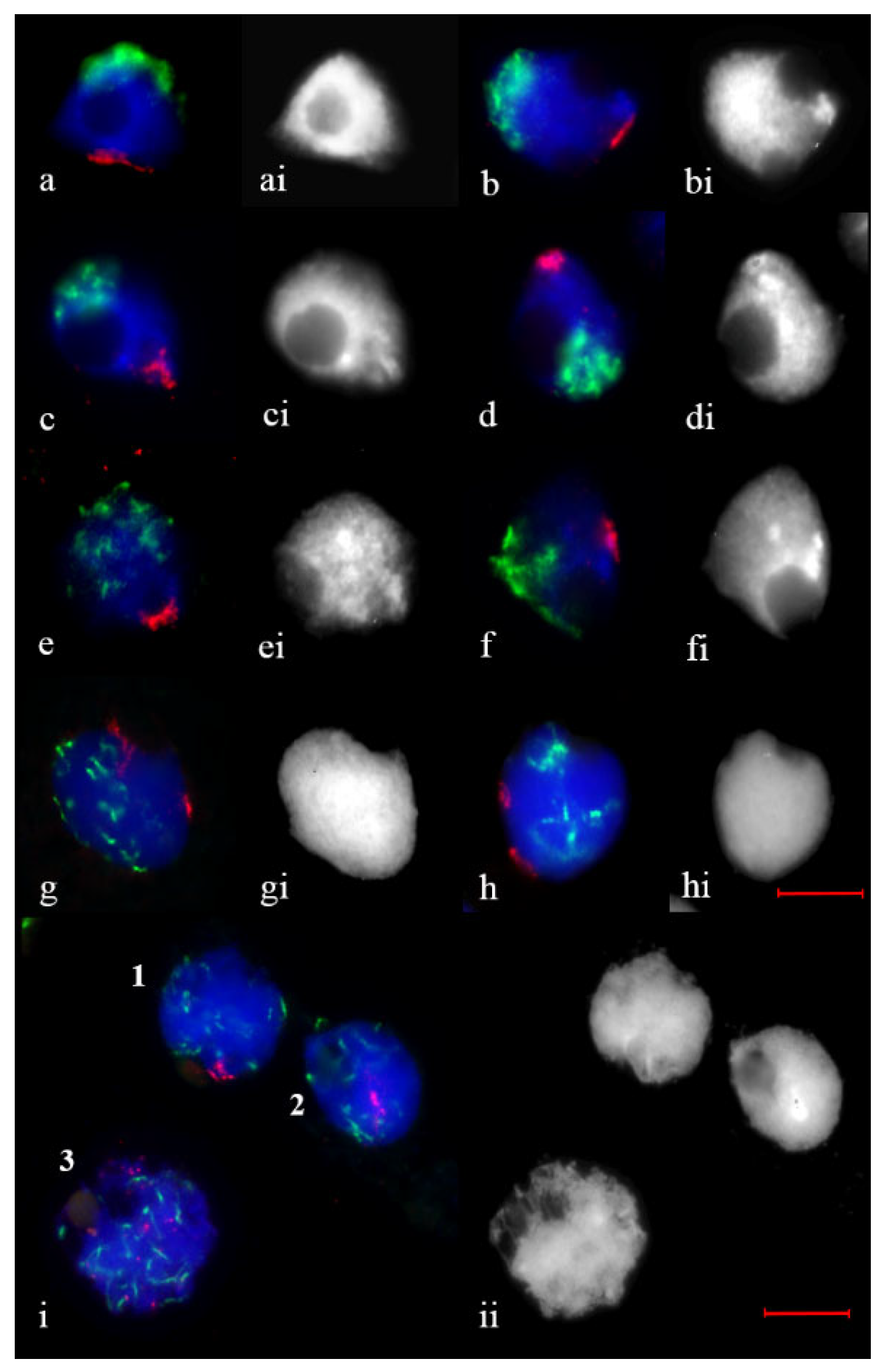
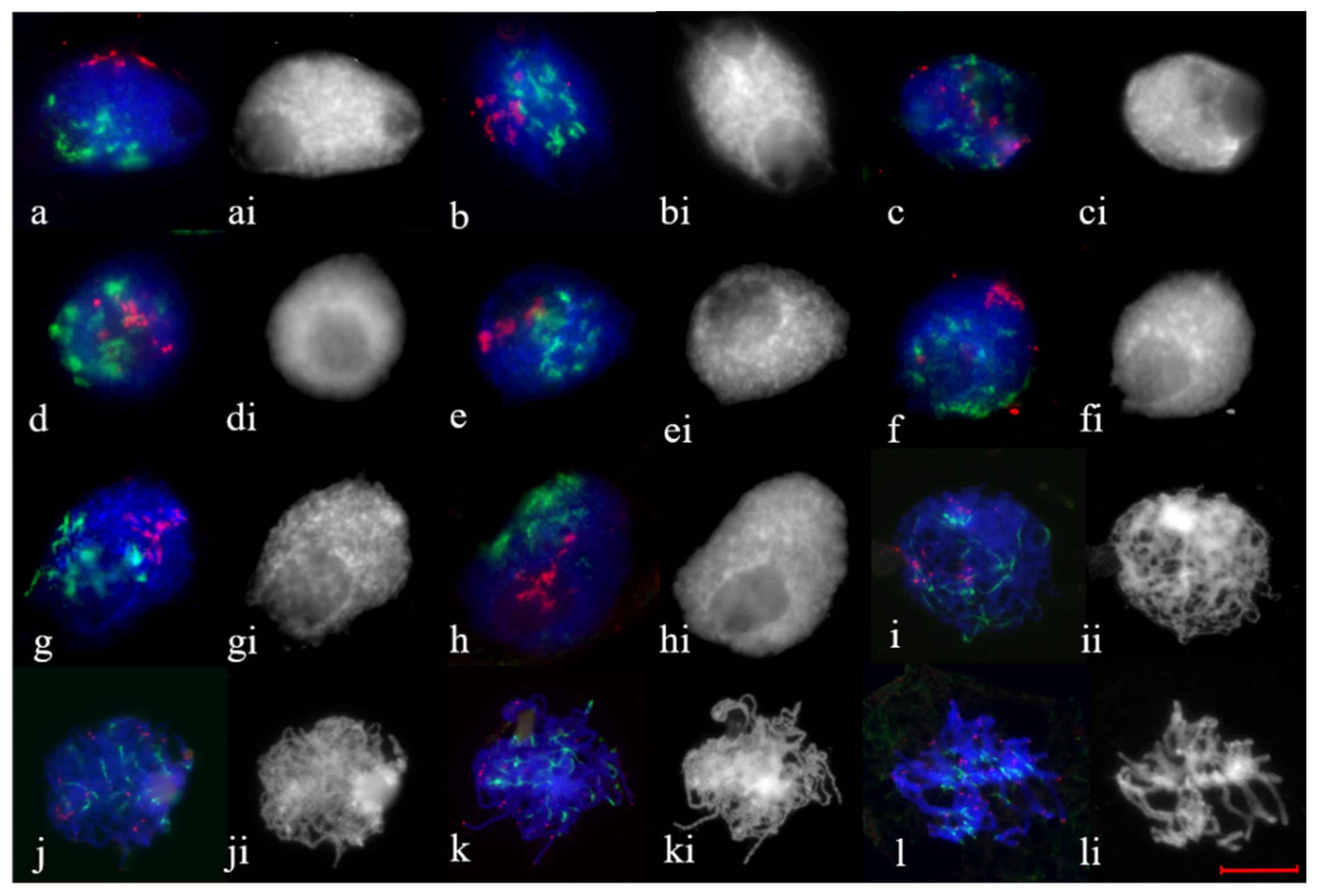
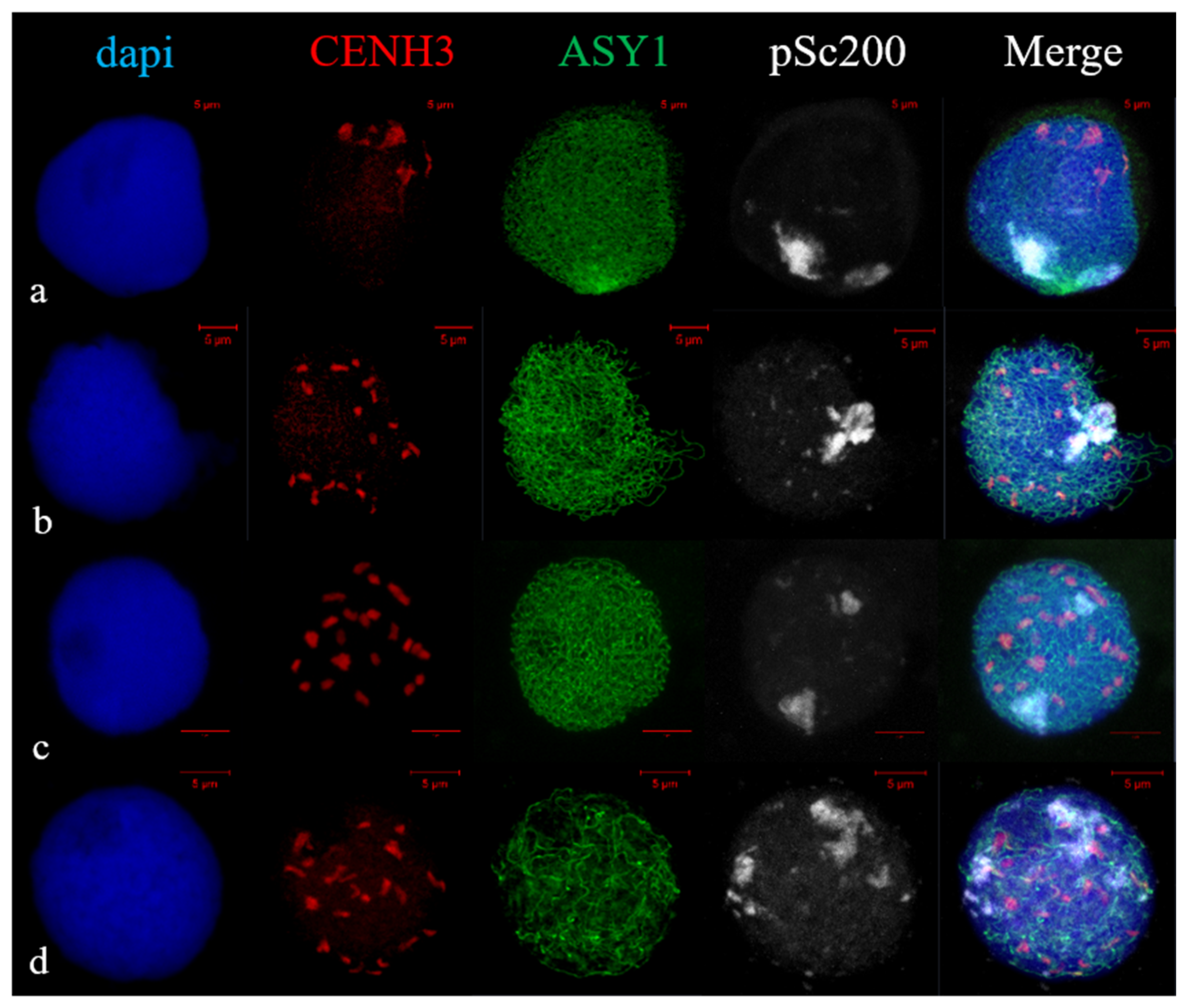
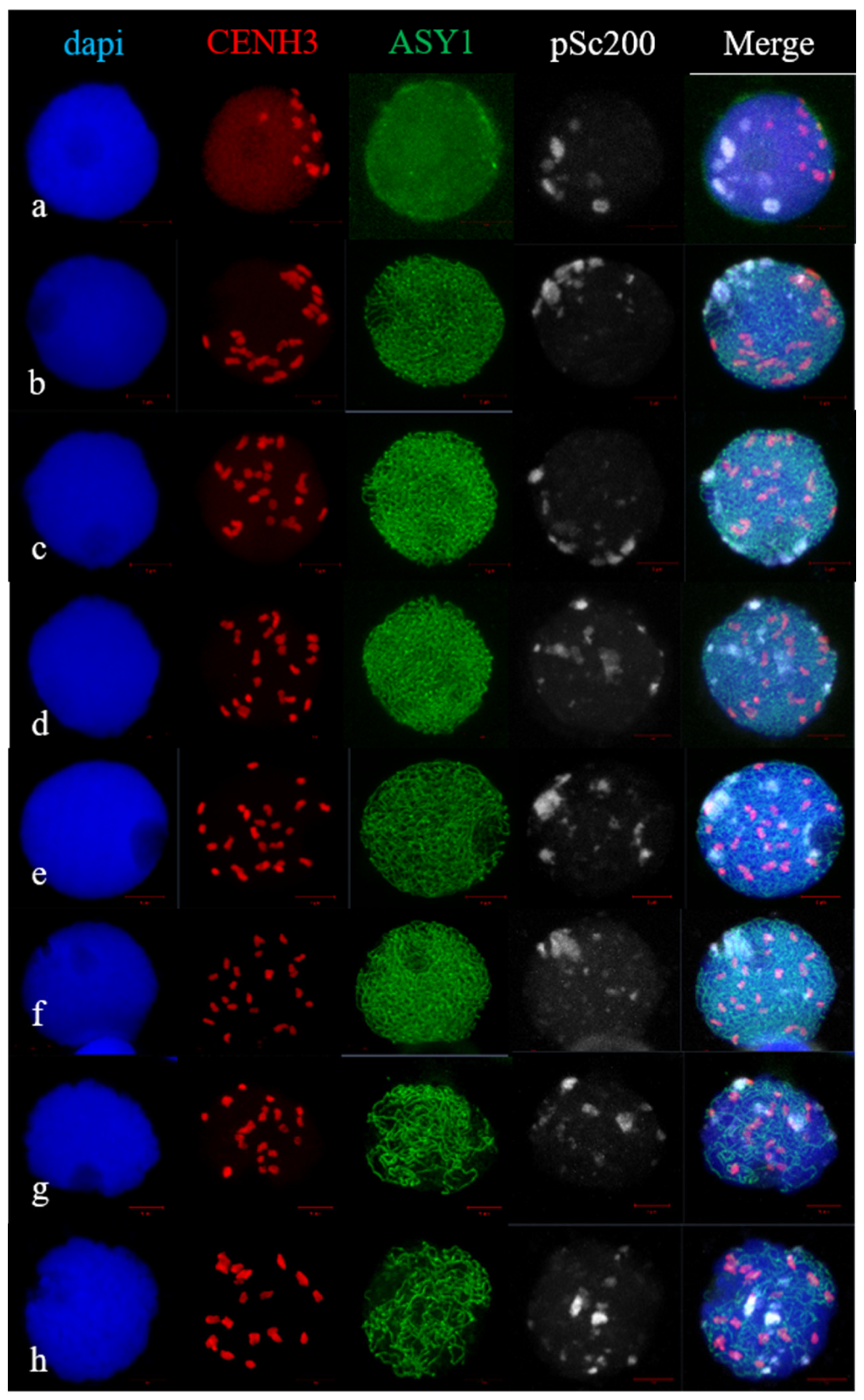
2.2. Telomere and Centromere Dynamics during Prophase in Wheat–Rye F1 Hybrids with Different Meiotic Pathway
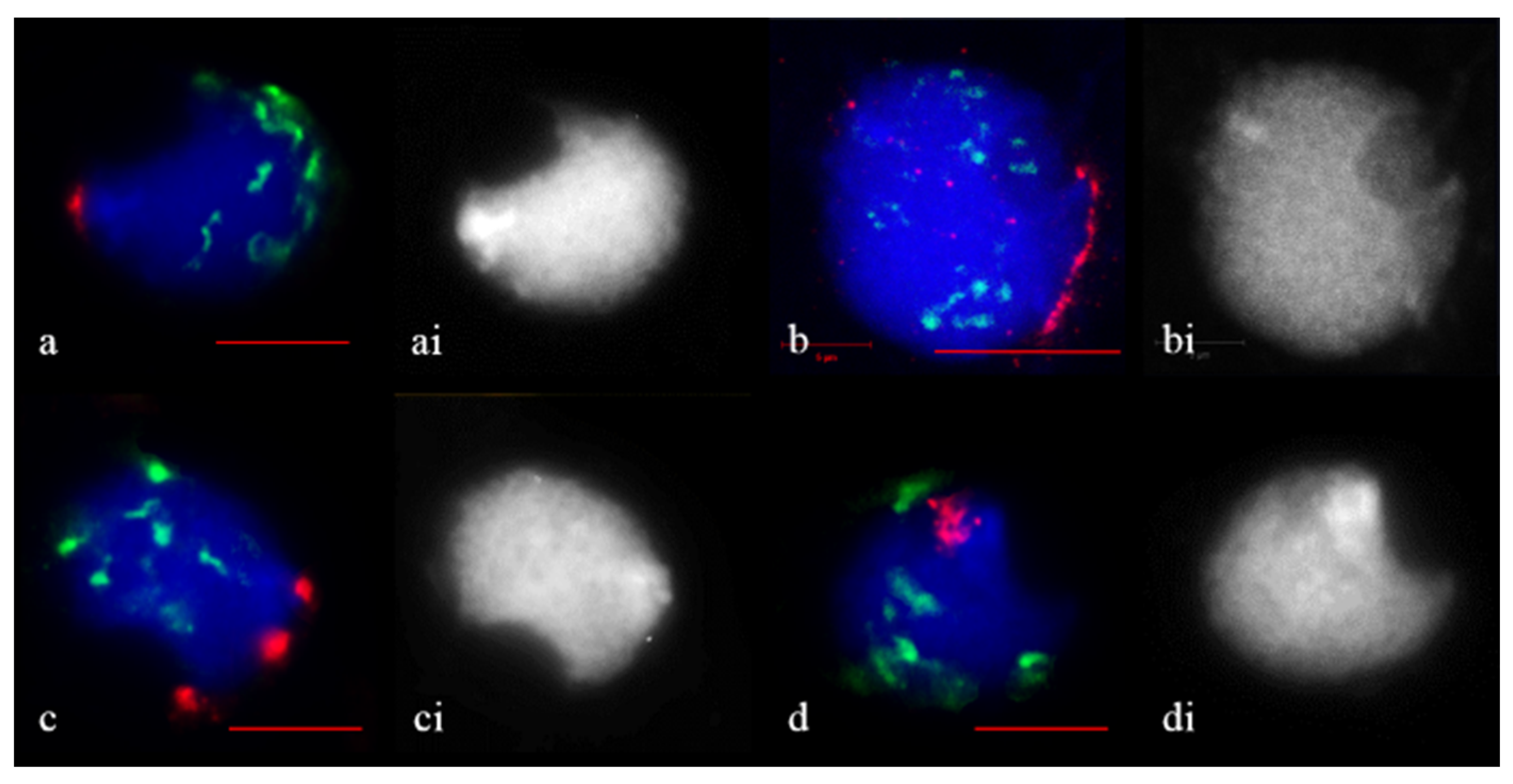
2.3. Subtelomere of Rye and Centromere Dynamics during Prophase in Wheat–Rye F1 Hybrids with Different Meiotic Pathway
3. Discussion
3.1. Defining the Time-Course of Early Prophase Progress in Wheat and Wheat–Rye Hybrids
3.2. Bouquet Formation in Wheat and Wheat–Rye F1 Hybrids with Different Meiotic Pathway
4. Materials and Methods
4.1. Plant Material
4.2. FISH and Immunostaining on Meiotic Chromosomes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mercier, R.; Mézard, C.; Jenczewski, E.; Macaisne, N.; Grelon, M. The molecular biology of meiosis in plants. Annu. Rev. Plant Biol. 2015, 66, 297–327. [Google Scholar] [CrossRef] [PubMed]
- Zickler, D.; Kleckner, N. The leptotene-zygotene transition of meiosis. Annu. Rev. Genet. 1998, 32, 619–697. Available online: https://www.annualreviews.org/doi/pdf/10.1146/annurev.genet.32.1.619 (accessed on 20 January 2022). [CrossRef] [PubMed] [Green Version]
- Zickler, D.; Kleckner, N. Recombination, pairing, and synapsis of homologs during meiosis. Cold Spring Harb. Perspect. Biol. 2015, 18, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamant, O.; Ma, H.; Cande, W.Z. Genetics of meiotic prophase I in plants. Annu. Rev. Plant. Biol. 2006, 57, 267–302. Available online: https://www.annualreviews.org/doi/pdf/10.1146/annurev.arplant.57.032905.105255 (accessed on 20 January 2022). [CrossRef] [PubMed]
- Stack, S.M.; Anderson, L.K. Crossing over as assessed by late recombination nodules is related to the pattern of synapsis and the distribution of early recombination nodules in maize. Chromosom. Res. 2002, 10, 329–345. [Google Scholar] [CrossRef] [PubMed]
- López, E.; Pradillo, M.; Romero, C.; Santos, J.L.; Cuñado, N. Pairing and synapsis in wild type Arabidopsis thaliana. Chromosom. Res. 2008, 16, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Sepsi, A.; Higgins, J.D.; Heslop-Harrison, J.S.; Schwarzacher, T. CENH3 morphogenesis reveals dynamic centromere associations during synaptonemal complex formation and the progression through male meiosis in hexaploid wheat. Plant J. 2017, 89, 235–249. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1111/tpj.13379 (accessed on 20 January 2022). [CrossRef] [Green Version]
- Bass, H.W.; Marshall, W.F.; Sedat, J.W.; Agard, D.A.; Cande, W.Z. Telomeres cluster de novo before the initiation of synapsis: A three-dimensional spatial analysis of telomere positions before and during meiotic prophase. J. Cell Biol. 1997, 137, 5–18. Available online: http://jcb.rupress.org/content/jcb/137/1/5.full.pdf (accessed on 20 January 2022). [CrossRef]
- Petrovská, B.; Šebela, M.; Doležel, J. Inside a plant nucleus: Discovering the proteins. J. Exp. Bot. 2015, 66, 1627–1640. [Google Scholar] [CrossRef]
- Poulet, A.; Probst, A.V.; Graumann, K.; Tatout, C.; Evans, D. Exploring the evolution of the proteins of the plant nuclear envelope. Nucleus 2017, 8, 46–59. [Google Scholar] [CrossRef] [Green Version]
- Alleva, B.; Smolikove, S. Moving and stopping: Regulation of chromosome movement to promote meiotic chromosome pairing and synapsis. Nucleus 2017, 8, 613–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koszul, R.; Kleckner, N. Dynamic chromosome movements during meiosis: A way to eliminate unwanted connections? Trends Cell Biol. 2009, 12, 716–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, B.; Su, H.; Birchler, J.A.; Han, F. Molecular mechanisms of homologous chromosome pairing and segregation in plants. J. Genet. Genom. 2014, 41, 117–123. Available online: https://www.sciencedirect.com/science/article/pii/S1673852713002129?via%3Dihub (accessed on 20 January 2022). [CrossRef] [PubMed]
- Hurel, A.; Phillips, D.; Vrielynck, N.; Mézard, C.; Grelon, M.; Christophorou, N. A cytological approach to studying meiotic recombination and chromosome dynamics in Arabidopsis thaliana male meiocytes in three dimensions. Plant J. 2018, 95, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Golubovskaya, I.N.; Harper, L.C.; Pawlowski, W.P.; Schichnes, D.; Cande, W.Z. The pam1 gene is required for meiotic bouquet formation and efficient homologous synapsis in maize (Zea mays L.). Genetics 2002, 162, 1979–1993. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1462385/pdf/12524364.pdf (accessed on 20 January 2022). [CrossRef]
- Harper, L.C.; Golubovskaya, I.N.; Cande, W.Z. A bouquet of chromosomes. J. Cell Sci. 2004, 117, 4025–4032. Available online: http://jcs.biologists.org/content/joces/117/18/4025.full.pdf (accessed on 20 January 2022). [CrossRef] [Green Version]
- Phillips, D.; Nibau, C.; Wnetrzak, J.; Jenkins, G. High resolution analysis of meiotic chromosome structure and behavior in barley (Hordeum vulgare L.). PLoS ONE 2012, 7, e39539. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.D.; Perry, R.M.; Barakate, A.; Ramsay, L.; Waugh, R.; Halpin, C.; Armstrong, S.J.; Franklin, F.C. Spatio-temporal asymmetry of the meiotic program underlies the predominantly distal distribution of meiotic crossovers in barley. Plant Cell 2012, 24, 4096–4109. [Google Scholar] [CrossRef] [Green Version]
- Sepsi, A.; Fábián, A.; Jäger, K.; Heslop-Harrison, J.S.; Schwarzacher, T. ImmunoFISH: Simultaneous visualisation of proteins and DNA sequences gives insight into meiotic processes in nuclei of grasses. Front. Plant Sci. 2018, 9, 1193. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6102387/pdf/fpls-09-01193.pdf (accessed on 20 January 2022). [CrossRef]
- Carlton, P.M.; Cowan, C.R.; Cande, W.Z. Directed motion of telomeres in the formation of the meiotic bouquet revealed by time course and simulation analysis. Mol. Biol. Cell 2003, 14, 2832–2843. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC165680/pdf/0142832.pdf (accessed on 20 January 2022). [CrossRef] [Green Version]
- Cowan, C.R.; Carlton, P.M.; Cande, W.Z. Reorganization and polarization of the meiotic bouquet-stage cell can be uncoupled from telomere clustering. J. Cell Sci. 2002, 115, 3757–3766. Available online: http://jcs.biologists.org/content/joces/115/19/3757.full.pdf (accessed on 20 January 2022). [CrossRef] [PubMed] [Green Version]
- Mikhailova, E.I.; Sosnikhina, S.P.; Kirillova, G.A.; Tikholiz, O.A.; Smirnov, V.G.; Jones, N.R.; Jenkins, G. Nuclear dispositions of subtelomeric and pericentromeric chromosomal domains during meiosis in asynaptic mutants of rye (Secale cereale L.). J. Cell Sci. 2001, 114, 1875–1882. Available online: http://jcs.biologists.org/content/joces/114/10/1875.full.pdf (accessed on 20 January 2022). [CrossRef] [PubMed]
- Mikhailova, E.I.; Phillips, D.; Sosnikhina, S.P.; Lovtsyus, A.V.; Jones, R.N.; Jenkins, G. Molecular assembly of meiotic proteins Asy1 and Zyp1 and pairing promiscuity in rye (Secale cereale L.) and its synaptic mutant sy10. Genetics 2006, 174, 1247–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Perez, E.; Shaw, P.J.; Moore, G. Polyploidy induces centromere association. J. Cell Biol. 2000, 148, 233–238. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2174288/pdf/9910061.pdf (accessed on 20 January 2022). [CrossRef] [PubMed] [Green Version]
- Martinez-Perez, E.; Shaw, P.; Aragon-Alcaide, L.; Moore, G. Chromosomes form into seven groups in hexaploid and tetraploid wheat as a prelude to meiosis. Plant J. 2003, 36, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobolth, P. Chromosome pairing in allohexaploid wheat var. Chinese Spring transformation of multivalent into bivalents, a mechanism for exclusive bivalent formation. Carlsberg Res. Commun. 1981, 46, 129–173. [Google Scholar] [CrossRef] [Green Version]
- Sears, E.R.; Okamoto, M. Intergenomic chromosome relationship in hexaploid wheat. Proc. Tenth Int. Congr. Genet. 1958, 2, 258–259. [Google Scholar]
- Riley, R.; Chapman, V. Genetic control of the cytologically diploid behaviour of hexaploid wheat. Nature 1958, 182, 713–715. [Google Scholar] [CrossRef]
- Griffiths, S.; Shar, R.; Foote, T.N.; Bertin, I.; Wanous, M.; Reader, S.; Colas, I.; Moore, G. Molecular characterization of Ph1 as a major chromosome pairing locus in polyploid wheat. Nature 2006, 439, 749–752. [Google Scholar] [CrossRef]
- Dhaliwal, H.S.; Gill, B.S.; Waines, J.G. Analysis of induced homoeologous pairing in a ph mutant wheat x rye hybrid. J. Hered. 1977, 68, 207–209. [Google Scholar] [CrossRef]
- Naranjo, T.; Fernandez-Rueda, P. Pairing and recombination between individual chromosomes of wheat and rye hybrids carrying the ph1b mutation. Theor. Appl. Genet. 1996, 93, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.C.; Shaw, P.; Phillips, D.; Reader, S.; Moore, G. Licensing MLH1 sites for crossover during meiosis. Nat. Commun. 2014, 5, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, E.; Martın, A.C.; Pendle, A.; Colas, I.; Jones, A.M.; Moore, G.; Shaw, P. The Ph1 locus suppresses Cdk2-type activity during premeiosis and meiosis in wheat. Plant Cell 2012, 24, 152–162. Available online: http://www.plantcell.org/content/plantcell/24/1/152.full.pdf (accessed on 20 January 2022). [CrossRef] [PubMed] [Green Version]
- Wang, X.; Holm, P.B. Chromosome pairing and synaptonemal complex formation in wheat-rye hybrids. Carlsberg Res. Commun. 1988, 53, 167–190. Available online: https://link.springer.com/content/pdf/10.1007%2FBF02907178.pdf (accessed on 20 January 2022). [CrossRef] [Green Version]
- Martinez-Perez, E.; Shaw, P.; Moore, G. The Ph1 locus is needed to ensure specific somatic and meiotic centromere association. Nature 2001, 411, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Joppa, L.R. Mechanisms and inheritance of first division restitution in hybrids of wheat, rye, and Aegilops squarrosa. Genome 1995, 38, 607–615. [Google Scholar] [CrossRef]
- Olesczuk, S.; Lukaszewski, A.J. The origin of unusual chromosome constitutions among newly formed allopolyploids. Am. J. Botany 2014, 101, 318–326. Available online: https://bsapubs.onlinelibrary.wiley.com/doi/epdf/10.3732/ajb.1300286 (accessed on 20 January 2022). [CrossRef] [Green Version]
- Silkova, O.G.; Loginova, D.B. Sister chromatid separation and monopolar spindle organization in the first meiosis as two mechanisms of unreduced gametes formation in wheat-rye hybrids. Plant Reprod. 2016, 29, 199–213. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4909807/pdf/497_2016_Article_279.pdf (accessed on 20 January 2022). [CrossRef] [Green Version]
- Loginova, D.B.; Silkova, O.G. Mitotic behavior of centromeres in meiosis as the fertility restoration mechanism in wheat-rye amphihaploids. Russ. J. Genet. 2014, 50, 818–827. Available online: https://link.springer.com/content/pdf/10.1134/S1022795414070114.pdf (accessed on 20 January 2022). [CrossRef]
- Aase, H.C. Cytology of Triticum, Secale, and Aegilops hybrids with reference to phylogeny. Res. Stud. State Coll. Wash. 1930, 2, 5–60. [Google Scholar]
- Maan, S.S.; Sasakuma, T. Fertility of amphihaploids in Triticinae. J. Hered. 1977, 57, 76–83. [Google Scholar] [CrossRef]
- Silkova, O.G.; Adonina, I.G.; Krivosheina, E.A.; Shchapova, A.I.; Shumny, V.K. Chromosome pairing in meiosis of partially fertile wheat–rye (ABDR) hybrids. Plant Reprod. 2013, 26, 33–41. Available online: https://link.springer.com/content/pdf/10.1007%2Fs00497-012-0207-2.pdf (accessed on 20 January 2022). [CrossRef]
- Bennett, M.D.; Rao, M.K.; Smith, J.B.; Bayliss, M.W. Cell development in the anther, the ovule, and the young seed of Triticum aestivum L. var. Chinese Spring. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1973, 266, 39–81. [Google Scholar] [CrossRef]
- Thomas, J.B.; Kaltsikes, P.J. A bouquet-like attachment plate for telomeres in leptotene of rye revealed by heterochromatin staining. Heredity 1976, 36, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, S.J.; Jones, G.H. Meiotic cytology and chromosome behaviour in wild-type Arabidopsis thaliana. J. Exp. Bot. 2003, 54, 1–10. [Google Scholar] [CrossRef]
- Hesse, S.; Zelkowski, M.; Mikhailova, E.I.; Keijzer, C.J.; Houben, A.; Schubert, V. Ultrastructure and dynamics of synaptonemal complex components during meiotic pairing and synapsis of standard (A) and accessory (B) rye chromosomes. Front. Plant Sci. 2019, 10, 773. [Google Scholar] [CrossRef] [Green Version]
- Zickler, D. From early homologue recognition to synaptonemal complex formation. Chromosoma 2006, 115, 158–174. [Google Scholar] [CrossRef]
- Prieto, P.; Shaw, P.; Moore, G. Homologue recognition during meiosis is associated with a change in chromatin conformation. Nat. Cell Biol. 2004, 6, 906–908. [Google Scholar] [CrossRef] [Green Version]
- Prieto, P.; Moore, G.; Reader, S. Control of conformation changes associated with homologue recognition during meiosis. Theor. Appl. Genet. 2005, 111, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Bass, H.W.; Bordoli, S.J.; Foss, E.M. The desynaptic (dy) and desynaptic1 (dsy1) mutations in maize (Zea mays L.) cause distinct telomere-misplacement phenotypes during meiotic prophase. J. Exp. Bot. 2003, 54, 39–46. [Google Scholar] [CrossRef]
- Murphy, S.P.; Bass, H.W. The maize (Zea mays) desynaptic (dy) mutation defines a pathway for meiotic chromosome segregation linking nuclear morphology, telomere distribution, and synapsis. J. Cell Sci. 2012, 125, 3681–3690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.P.; Gumber, H.K.; Mao, Y.; Bass, H.W. A dynamic meiotic SUN belt includes the zygotene-stage telomere bouquet and is disrupted in chromosome segregation mutants of maize (Zea mays L.). Front. Plant Sci. 2014, 5, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Ma, L.; Zhang, C.; Du, G.; Shen, Y.; Tang, D.; Li, Y.; Yu, H.; Ma, B.; Cheng, Z. The SUN Domain Proteins OsSUN1 and OsSUN2 Play Critical but Partially Redundant Roles in Meiosis. Plant Physiol. 2020, 183, 1517–1530. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, M.J.; Pawlowski, W.P. Live imaging of rapid chromosome movements in meiotic prophase I in maize. Proc. Natl. Acad. Sci. USA 2009, 106, 20989–20994. [Google Scholar] [CrossRef] [Green Version]
- Scherthan, H. Telomere attachment and clustering during meiosis. Cell. Mol. Life Sci. 2007, 64, 117–124. [Google Scholar] [CrossRef]
- Silkova, O.G.; Dobrovolskaya, O.B.; Dubovets, N.I.; Adonina, I.G.; Kravtsova, L.A.; Roder, M.S.; Salina, E.A.; Shchapova, A.I.; Shumny, V.K. Production of wheat–rye substitution lines and identification of chromosome composition of karyotypes using C-banding, GISH, and SSR markers. Rus. J. Genet. 2006, 42, 645–653. Available online: https://link.springer.com/content/pdf/10.1134%2FS1022795406060093.pdf (accessed on 20 January 2022). [CrossRef]
- Silkova, O.G.; Dobrovolskaya, O.B.; Dubovets, N.I.; Adonina, I.G.; Kravtsova, L.A.; Shchapova, A.I.; Shumny, V.K. Production of wheat–rye substitution lines based on winter rye cultivars with karyotype identification by means of C-banding, GISH, and SSR markers. Russ. J. Genet. 2007, 43, 957–960. Available online: https://link.springer.com/content/pdf/10.1134%2FS1022795407080200.pdf (accessed on 20 January 2022). [CrossRef]
- Cheung, W.Y.; Money, T.A.; Abbo, S.; Devos, K.M.; Gale, M.D.; Moore, G. A family of related sequences associated with (TTTAGGG)n repeats are located in the interstitial regions of wheat chromosomes. Mol. Gen. Genet. 1994, 245, 349–354. [Google Scholar] [CrossRef]
- Zhang, P.; Wanlong, L.; Fellers, J.; Friebe, B.; Gill, B.S. BAC-FISH in wheat identifies chromosome landmarks consisting of different types of transposable elements. Chromosoma 2004, 112, 288–299. [Google Scholar] [CrossRef]
- Qi, L.L.; Wu, J.J.; Friebe, B.; Qian, C.; Gu, Y.Q.; Fu, D.L.; Gill, B.S. Sequence organization and evolutionary dynamics of Brachypodium-specific centromere retrotransposons. Chromosom. Res. 2013, 21, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Schubert, V.; Ruban, A.; Houben, A. Chromatin ring formation at plant centromeres. Front. Plant Sci. 2016, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vershinin, A.V.; Schwarzacher, T.; Heslop-Harrison, J.S. The large-scale genomic organization of repetitive DNA families at the telomeres of rye chromosomes. Plant Cell 1995, 7, 1823–1833. Available online: http://www.plantcell.org/content/plantcell/7/11/1823.full.pdf (accessed on 20 January 2022). [CrossRef] [PubMed] [Green Version]
- Manzanero, S.; Arana, P.; Puertas, M.J.; Houben, A. The chromosomal distribution of phosphorylated histone H3 differs between plants and animals at meiosis. Chromosoma 2000, 109, 308–317. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrids | Tight Cluster | One Diffuse Cluster | Two Diffuse Clusters | Clumps | Location along the Nucleus Circumference |
|---|---|---|---|---|---|
| 2R(2D) × R | 26.5 | 52.4 | 21.1 | 0 | 0 |
| 1R(1A) × R | 6.3 | 5.1 | 0 | 60.4 | 28.2 |
| 5R(5D) × R | 8.6 | 36.2 | 0 | 32.2 | 23 |
| 6R(6A) × R | 17.4 | 10.2 | 0 | 64.1 | 8.3 |
| Hybrids | Cluster | Two Clusters | Clumps | Location along the Nucleus Circumference |
|---|---|---|---|---|
| 2R(2D) × R | 83.2 | 16.8 | 0 | 0 |
| 1R(1A) × R | 12.19 | 9.75 | 65.87 | 12.19 |
| FISH | Immunostaining + FISH | ||||
|---|---|---|---|---|---|
| Plants | Anthers/Slides | Meiocytes | Plants | Meiocytes | |
| 2R(2D) × R | 5 | 14 | 863 | 3 | 97 |
| 1R(1A) × R | 5 | 16 | 1637 | 5 | 41 |
| 5R(5D) × R | 3 | 8 | 476 | - | - |
| 6R(6A) × R | 3 | 9 | 661 | - | - |
| S29 | 3 | 9 | 353 | - | - |
| 2R(2D) | 5 | 10 | 756 | - | - |
| 1R(1A) | 5 | 16 | 1036 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silkova, O.G.; Loginova, D.B.; Zhuravleva, A.A.; Shumny, V.K. Bouquet Formation Failure in Meiosis of F1 Wheat–Rye Hybrids with Mitotic-Like Division. Plants 2022, 11, 1582. https://doi.org/10.3390/plants11121582
Silkova OG, Loginova DB, Zhuravleva AA, Shumny VK. Bouquet Formation Failure in Meiosis of F1 Wheat–Rye Hybrids with Mitotic-Like Division. Plants. 2022; 11(12):1582. https://doi.org/10.3390/plants11121582
Chicago/Turabian StyleSilkova, Olga G., Dina B. Loginova, Anastasia A. Zhuravleva, and Vladimir K. Shumny. 2022. "Bouquet Formation Failure in Meiosis of F1 Wheat–Rye Hybrids with Mitotic-Like Division" Plants 11, no. 12: 1582. https://doi.org/10.3390/plants11121582