
Seed Viability and Potential Germination Rate of Nine Endemic Boswellia Taxa (Burseraceae) from Socotra Island (Yemen)

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
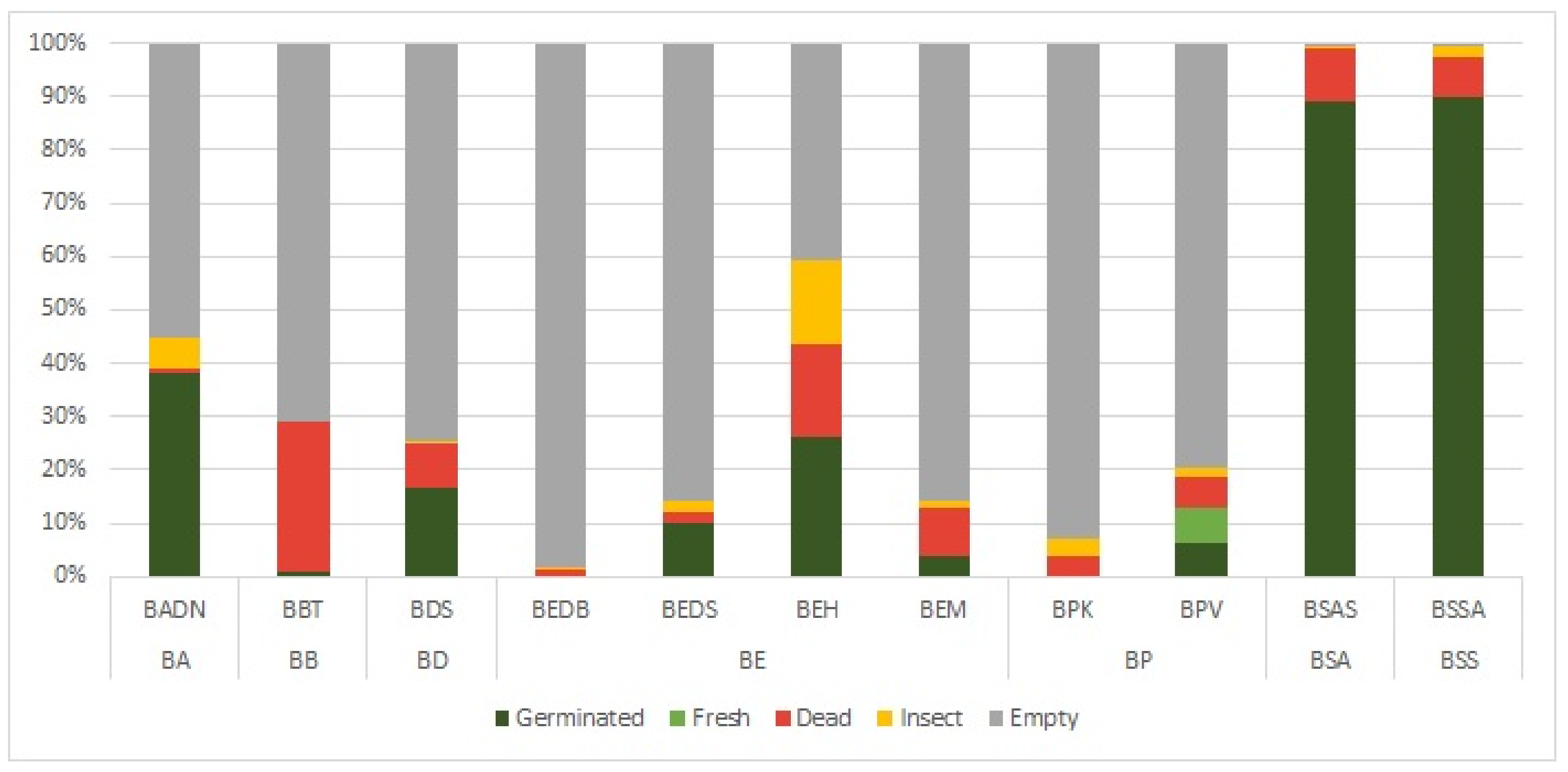
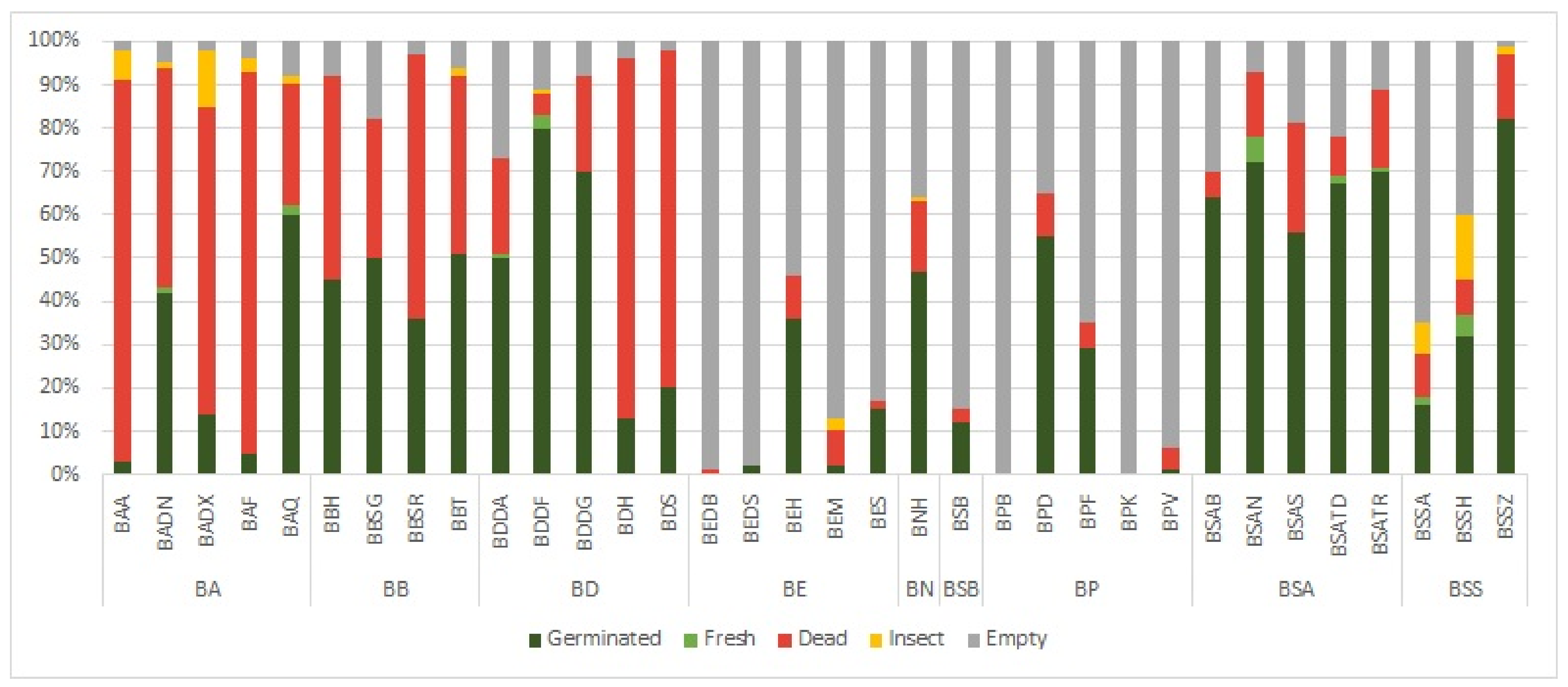
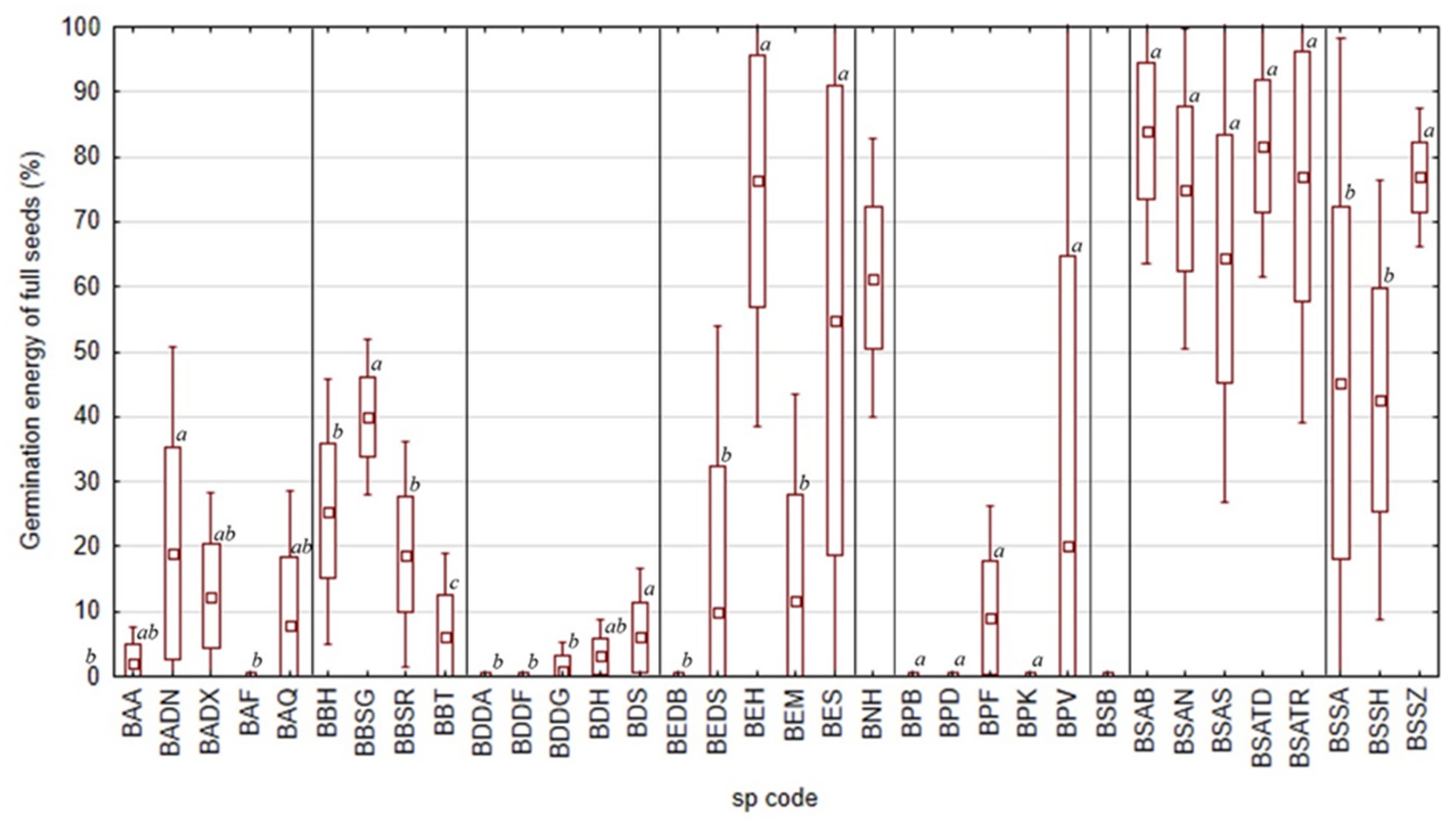
2.1. Inter-Locality Variabilities of Different Boswellia Taxa in 2020 and 2021
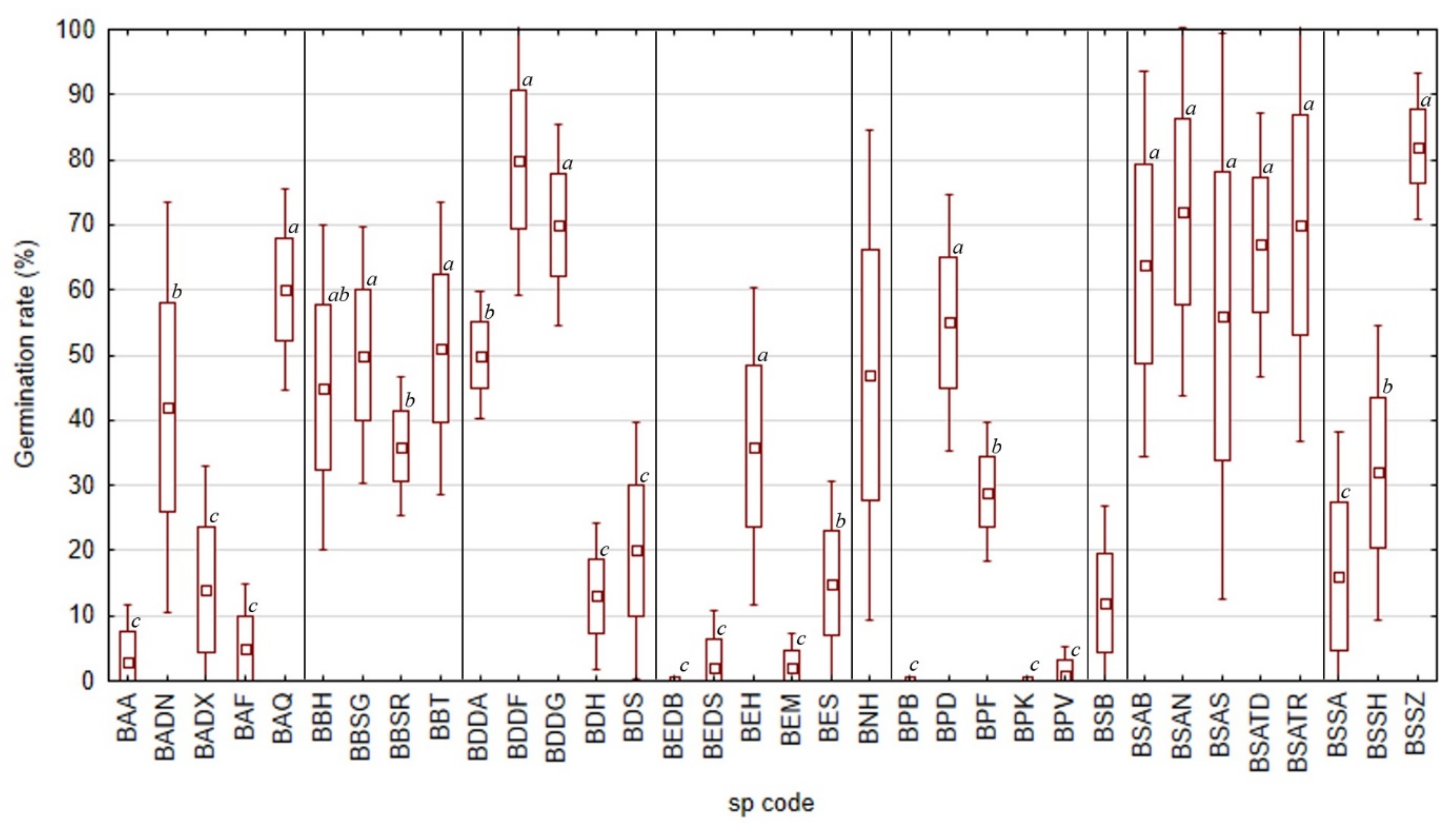
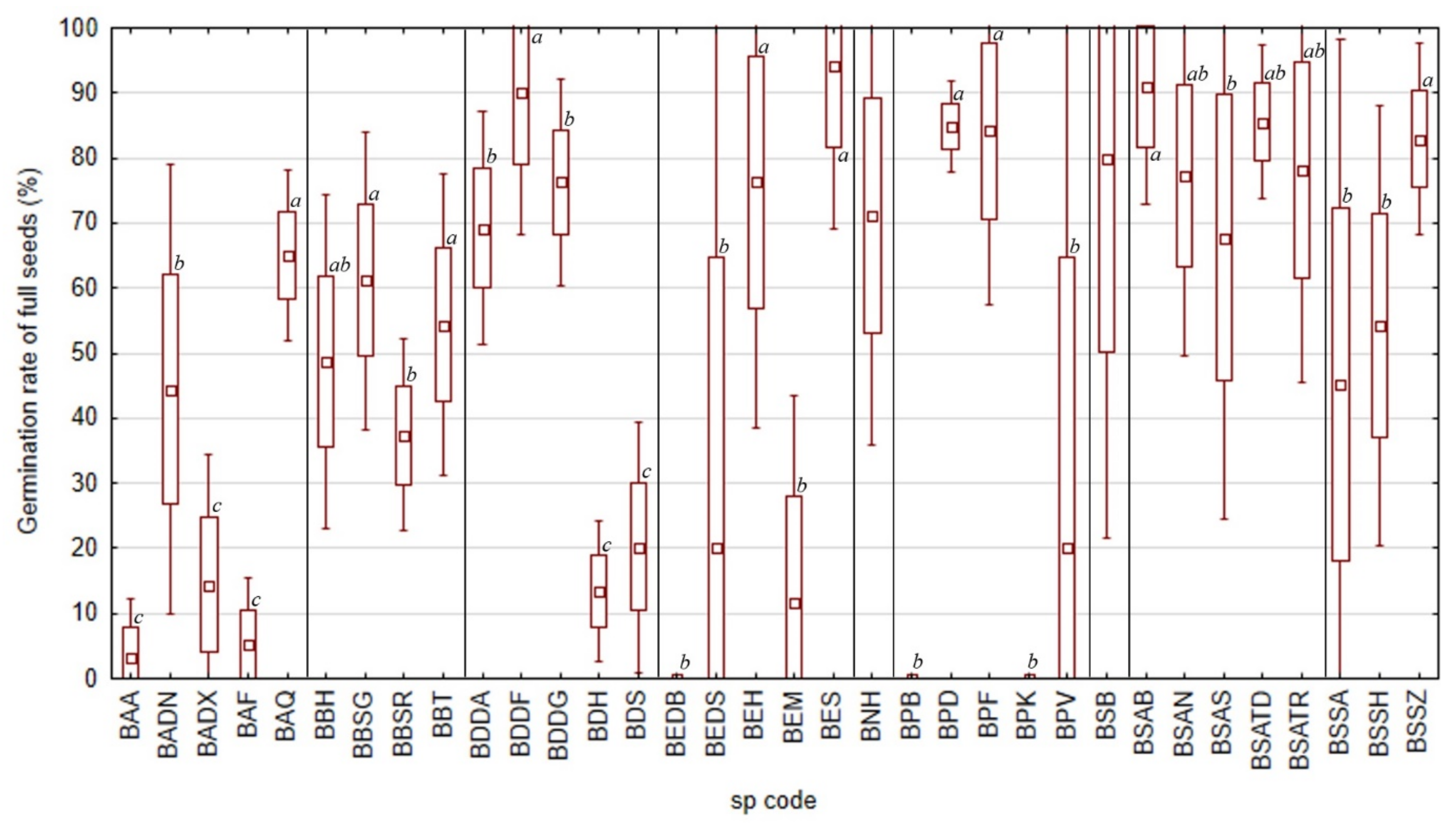
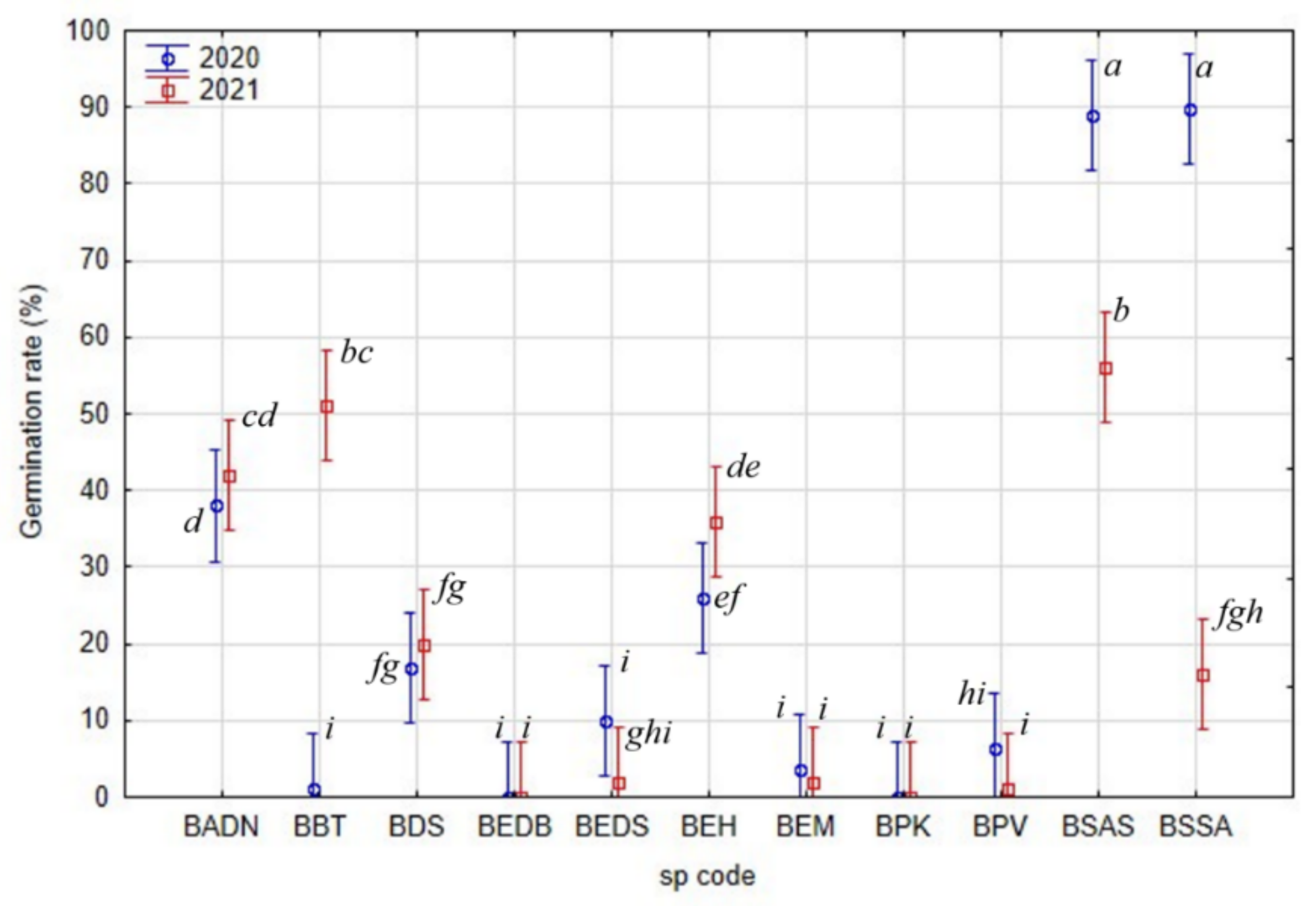
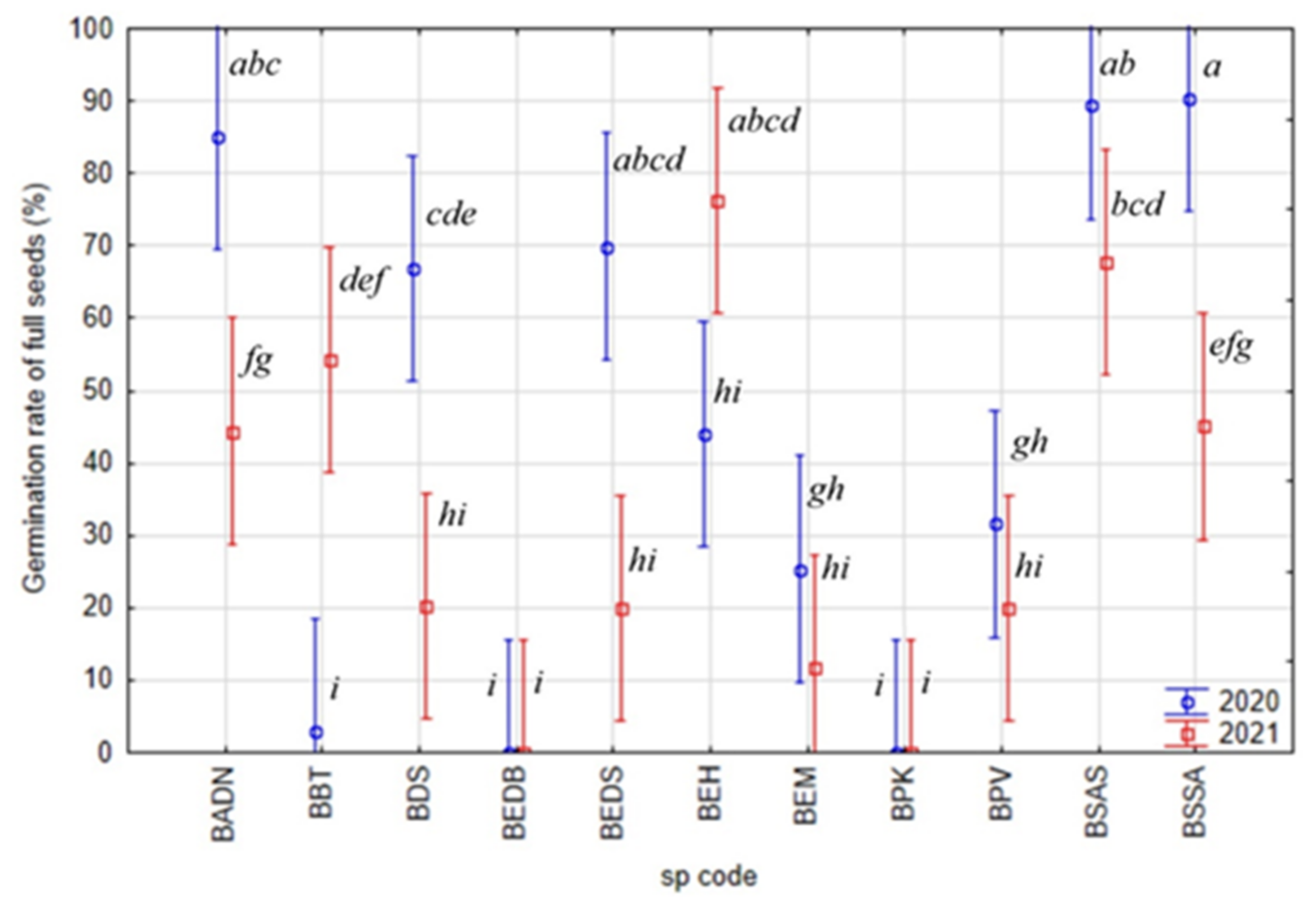
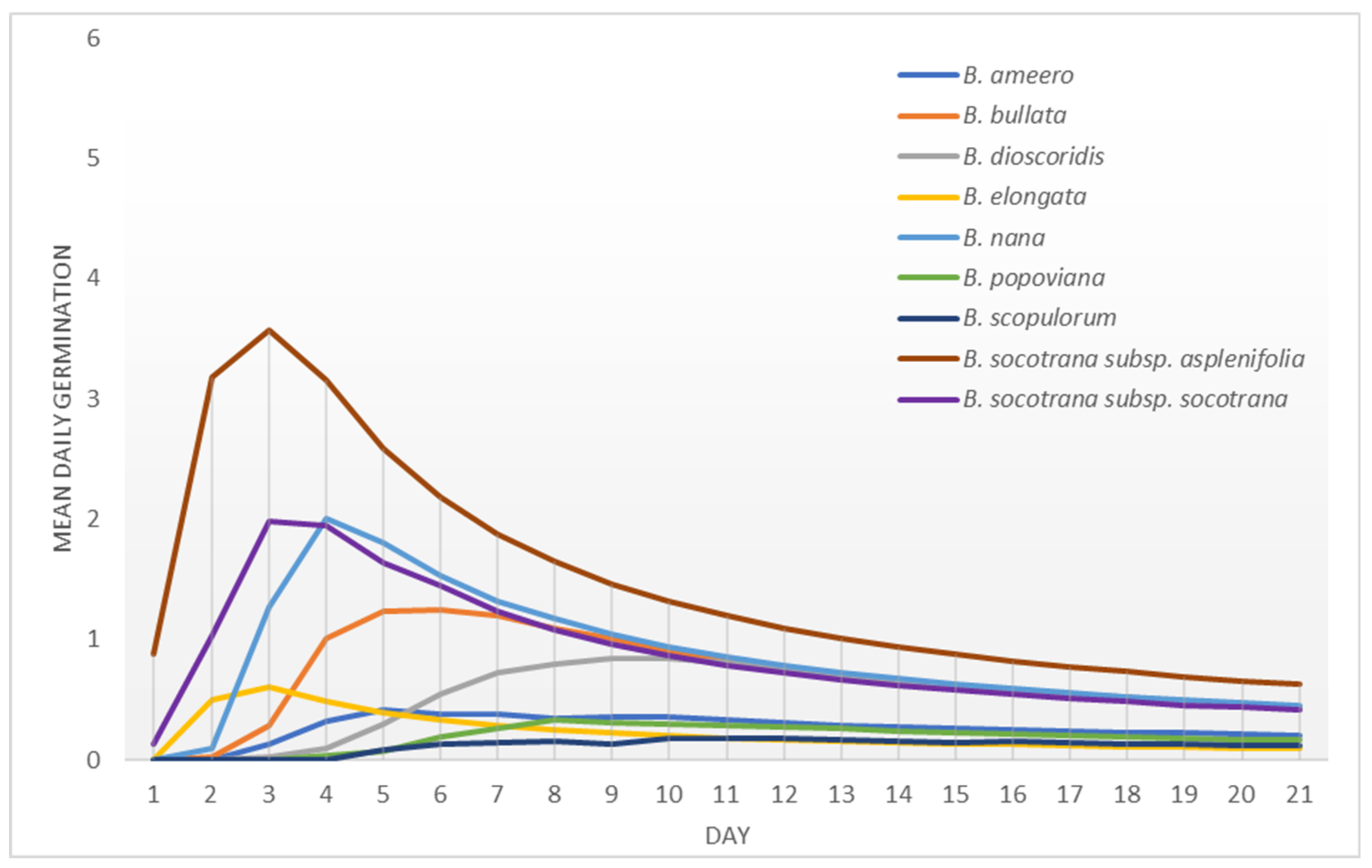
2.2. Average Inter-Annual and Inter-Species Variability of Germination Rates
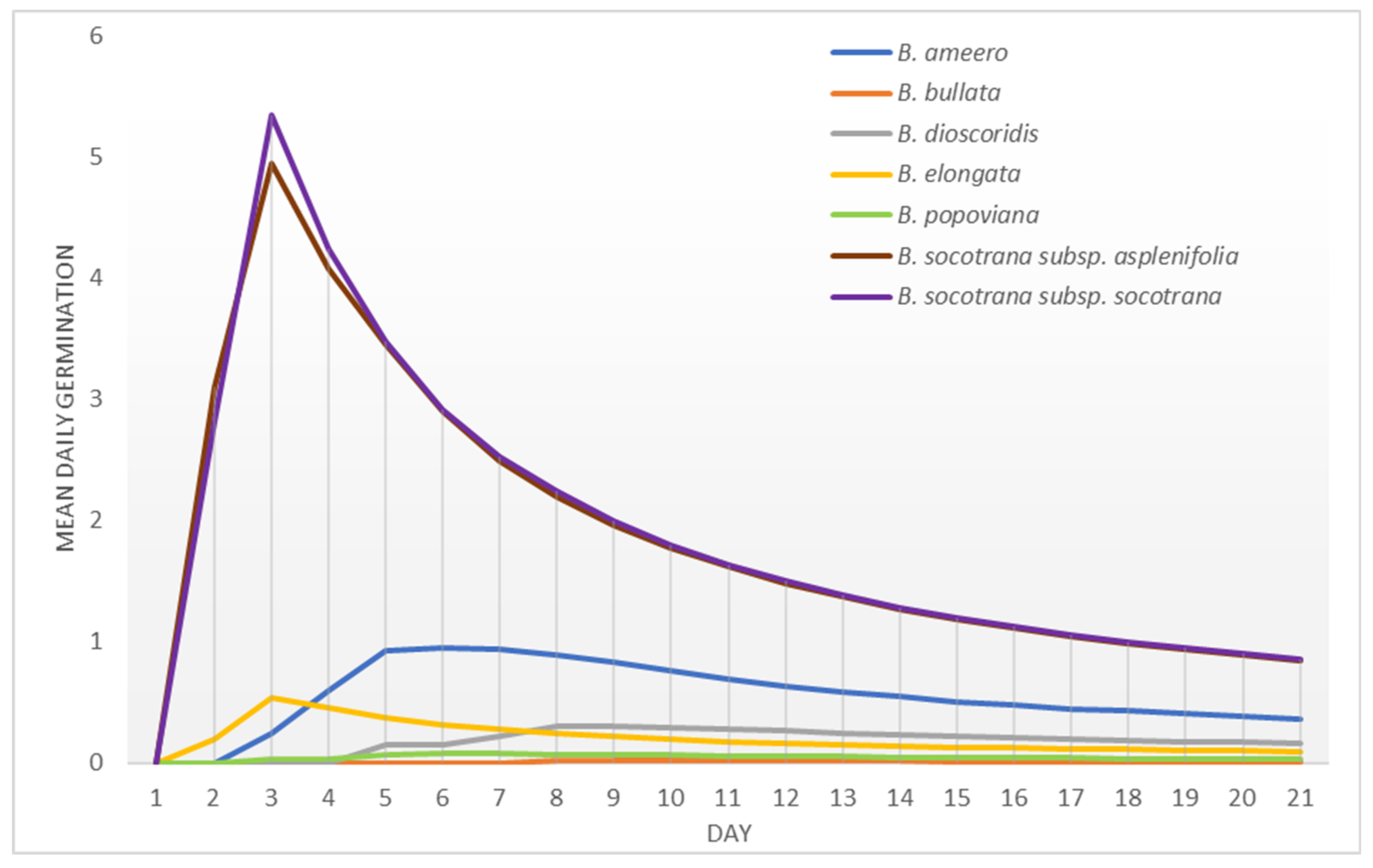
2.3. Peak Value Indices
3. Discussion
3.1. Factors Affecting Germination of Boswellia Seeds
3.2. Phenology of Fruit Ripening
3.3. Natural Regeneration
4. Materials and Methods
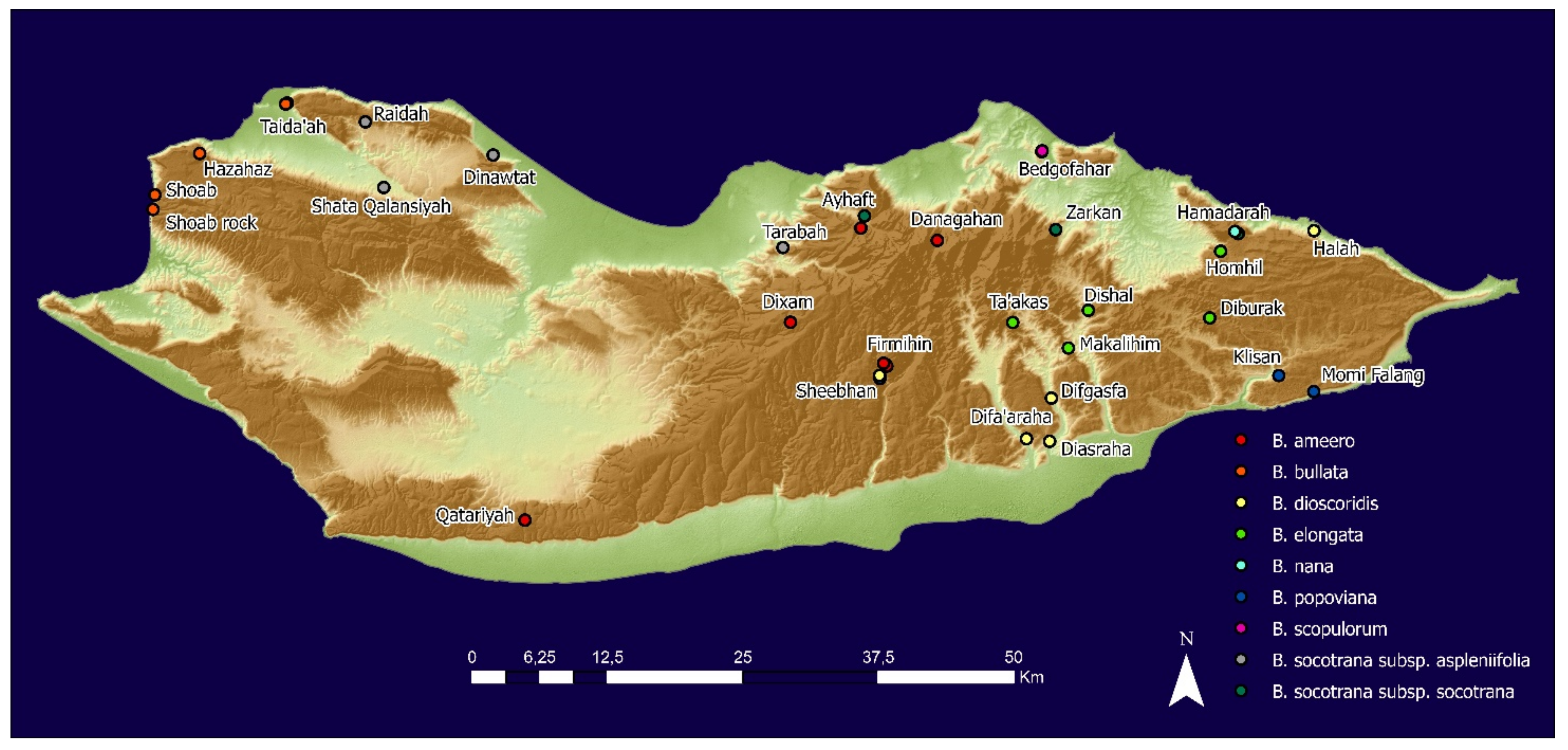
4.1. Seed Collection and Preparation
4.2. Trials
- fresh—the tissue inside was solid, light or greenish, assuming start of germination in a longer time than the length of the germination trial;
- empty—without any tissue inside or remnants of tissue that filled less than half the internal space of the seed;
- dead—the tissue inside was soft, brownish and filled more than half the internal space of the seed;
- damaged by insects—seeds containing insects at different stages of development or an insect exit hole was visible in the seed coat.
4.3. Data Evaluation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Boswellia Species | Sp. Code | Locality | Germination Rate (%) | Germination Rate of Full Seeds (%) | Germination Energy (%) | Germination Energy of Full Seeds (%) | Fresh (%) | Dead (%) | Empty (%) | Insect (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| B. ameero | BADN | Danagahan | 38 ± 7.13 | 85 ± 7.13 | 12 ± 5.34 | 27 ± 11.05 | 0 ± 0 | 1 ± 2.4 | 55 ± 9.42 | 6 ± 2.64 |
| B. bullata | BBT | Taida’ah (Ditwah mountain) | 1 ± 2 | 3 ± 5.71 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 28 ± 4 | 71 ± 4.9 | 0 ± 0 |
| B. dioscoridis | BDS | Sheebhan | 17 ± 4.12 | 67 ± 8.25 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 8 ± 2.61 | 75 ± 6.97 | 1 ± 1.2 |
| B. elongata | BEDB | Diburak | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 1 ± 1.02 | 98 ± 1.47 | 0 ± 0.49 |
| B. elongata | BEDS | Dishal | 10 ± 3.03 | 70 ± 8 | 9 ± 3.66 | 63 ± 15.22 | 0 ± 0 | 2 ± 1.17 | 86 ± 2.9 | 2 ± 0.98 |
| B. elongata | BEH | Homhil | 26 ± 3.41 | 44 ± 3.44 | 25 ± 2.93 | 43 ± 4.07 | 0 ± 0 | 17 ± 1.74 | 41 ± 8.23 | 16 ± 5.19 |
| B. elongata | BEM | Makalihim | 4 ± 1.62 | 25 ± 7.47 | 2 ± 1.02 | 19 ± 9.15 | 0 ± 0 | 9 ± 2.73 | 86 ± 3.9 | 1 ± 1.55 |
| B. popoviana | BPK | Klisan | 0 ± 0 | 0 ± 0 | 0 ± 0.4 | 2 ± 4.44 | 0 ± 0 | 4 ± 2.5 | 93 ± 2.61 | 3 ± 1.85 |
| B. popoviana | BPV | Momi Falang | 6 ± 1.02 | 32 ± 4.59 | 1 ± 1.17 | 6 ± 5.57 | 6 ± 1.02 | 6 ± 1.94 | 80 ± 2.15 | 2 ± 0.75 |
| B. socotrana subsp. aspleniifolia | BSAS | Shata Qalansiyah | 89 ± 3.85 | 89 ± 4.01 | 81 ± 3.61 | 82 ± 3.87 | 0 ± 0 | 10 ± 3.6 | 0 ± 0.49 | 0 ± 0.49 |
| B. socotrana subsp. socotrana | BSSA | Ayhaft | 90 ± 2.32 | 90 ± 2.17 | 85 ± 3.06 | 85 ± 3.18 | 0 ± 0 | 7 ± 2.73 | 1 ± 0.8 | 2 ± 0.98 |
| Boswellia Species | Sp. Code | Locality | Germination Rate (%) | Germination Rate of Full Seeds (%) | Germination Energy (%) | Germination Energy of Full Seeds (%) | Fresh (%) | Dead (%) | Empty (%) | Insect (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| B. ameero | BAA | Ayhaft | 3 ± 4 | 3 ± 4.19 | 2 ± 2.45 | 2 ± 2.52 | 0 ± 0 | 88 ± 8.72 | 2 ± 2.45 | 7 ± 5.1 |
| B. ameero | BADN | Danagahan | 42 ± 14.35 | 44 ± 15.76 | 18 ± 13.64 | 19 ± 14.61 | 1 ± 2 | 51 ± 15.62 | 5 ± 4.47 | 1 ± 2 |
| B. ameero | BADX | Dixam | 14 ± 8.6 | 14 ± 9.15 | 12 ± 6.78 | 12 ± 7.26 | 0 ± 0 | 71 ± 13.19 | 2 ± 2.45 | 13 ± 5.1 |
| B. ameero | BAF | Firmihin | 5 ± 4.47 | 5 ± 4.73 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 88 ± 7.48 | 4 ± 4.9 | 3 ± 4 |
| B. ameero | BAQ | Qatariyah | 60 ± 7.07 | 65 ± 6 | 7 ± 8.72 | 8 ± 9.54 | 2 ± 2.45 | 28 ± 4 | 8 ± 4 | 2 ± 2.45 |
| B. bullata | BBH | Hazahaz | 45 ± 11.4 | 49 ± 11.73 | 24 ± 10.2 | 26 ± 9.31 | 0 ± 0 | 47 ± 9.8 | 8 ± 6.78 | 0 ± 0 |
| B. bullata | BBSG | Shoab rock (green flowers) | 50 ± 8.94 | 61 ± 10.47 | 33 ± 6.78 | 40 ± 5.44 | 0 ± 0 | 32 ± 9.27 | 18 ± 6.78 | 0 ± 0 |
| B. bullata | BBSR | Shoab (red flower) | 36 ± 4.9 | 37 ± 6.71 | 18 ± 6.78 | 19 ± 7.87 | 0 ± 0 | 61 ± 8.6 | 3 ± 4 | 0 ± 0 |
| B. bullata | BBT | Taida’ah (Ditwah mountain) | 51 ± 10.2 | 54 ± 10.58 | 6 ± 5.83 | 6 ± 5.81 | 0 ± 0 | 41 ± 10.68 | 6 ± 5.83 | 2 ± 2.45 |
| B. dioscoridis | BDDA | Wadi Diasraha | 50 ± 4.47 | 69 ± 8.17 | 0 ± 0 | 0 ± 0 | 1 ± 2 | 22 ± 8.12 | 27 ± 9.27 | 0 ± 0 |
| B. dioscoridis | BDDF | Wadi Difa’araha | 80 ± 9.49 | 90 ± 10 | 0 ± 0 | 0 ± 0 | 3 ± 2.45 | 5 ± 6.32 | 11 ± 8.6 | 1 ± 2 |
| B. dioscoridis | BDDG | Wadi Digasfa | 70 ± 7.07 | 76 ± 7.21 | 1 ± 2 | 1 ± 2 | 0 ± 0 | 22 ± 7.48 | 8 ± 6.78 | 0 ± 0 |
| B. dioscoridis | BDH | Halah (Hoq cave) | 13 ± 5.1 | 13 ± 4.93 | 3 ± 2.45 | 3 ± 2.55 | 0 ± 0 | 83 ± 2.45 | 4 ± 3.74 | 0 ± 0 |
| B. dioscoridis | BDS | Sheebhan | 20 ± 8.94 | 20 ± 8.77 | 6 ± 4.9 | 6 ± 4.89 | 0 ± 0 | 78 ± 6.78 | 2 ± 2.45 | 0 ± 0 |
| B. elongata | BEDB | Diburak | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 1 ± 2 | 99 ± 2 | 0 ± 0 |
| B. elongata | BEDS | Dishal | 2 ± 4 | 20 ± 40 | 1 ± 2 | 10 ± 20 | 0 ± 0 | 0 ± 0 | 98 ± 4 | 0 ± 0 |
| B. elongata | BEH | Homhil | 36 ± 11.14 | 76 ± 17.3 | 36 ± 11.14 | 76 ± 17.3 | 0 ± 0 | 10 ± 5.48 | 54 ± 6.63 | 0 ± 0 |
| B. elongata | BEM | Makalihim | 2 ± 2.45 | 12 ± 14.53 | 2 ± 2.45 | 12 ± 14.53 | 0 ± 0 | 8 ± 2.45 | 87 ± 5.1 | 3 ± 2.45 |
| B. elongata | BES | Shabarah | 15 ± 7.07 | 94 ± 11.43 | 11 ± 8 | 55 ± 32.3 | 0 ± 0 | 2 ± 4 | 83 ± 10.3 | 0 ± 0 |
| B. nana | BNH | Hamadarah (Homhil) | 47 ± 17.2 | 71 ± 16.19 | 40 ± 11.4 | 61 ± 9.83 | 0 ± 0 | 16 ± 7.35 | 36 ± 9.7 | 1 ± 2 |
| B. popoviana | BPB | Bedgofahar | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 100 ± 0 | 0 ± 0 |
| B. popoviana | BPD | Sheebhan (Didrafarantan) | 55 ± 8.94 | 85 ± 3.23 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 10 ± 3.16 | 35 ± 11.4 | 0 ± 0 |
| B. popoviana | BPF | Firmihin | 29 ± 4.9 | 84 ± 12.19 | 3 ± 2.45 | 9 ± 7.84 | 0 ± 0 | 6 ± 4.9 | 65 ± 6.32 | 0 ± 0 |
| B. popoviana | BPK | Klisan | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 100 ± 0 | 0 ± 0 |
| B. popoviana | BPV | Momi Falang | 1 ± 2 | 20 ± 40 | 1 ± 2 | 20 ± 40 | 0 ± 0 | 5 ± 4.47 | 94 ± 3.74 | 0 ± 0 |
| B. scopulorum | BSB | Bedgofahar | 12 ± 6.78 | 80 ± 26.67 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 3 ± 4 | 85 ± 5.48 | 0 ± 0 |
| B. socotrana subsp. aspleniifolia | BSAB | Raidah (Taida’ah) | 64 ± 13.56 | 91 ± 8.27 | 59 ± 12.41 | 84 ± 9.39 | 0 ± 0 | 6 ± 5.83 | 30 ± 11.4 | 0 ± 0 |
| B. socotrana subsp. aspleniifolia | BSAN | Dinawtat | 72 ± 12.88 | 77 ± 12.6 | 70 ± 12.25 | 75 ± 11.27 | 6 ± 3.74 | 15 ± 8.37 | 7 ± 4 | 0 ± 0 |
| B. socotrana subsp. aspleniifolia | BSAS | Shata Qalansiyah | 56 ± 19.85 | 68 ± 19.77 | 53 ± 17.49 | 64 ± 17.17 | 0 ± 0 | 25 ± 13.04 | 19 ± 11.58 | 0 ± 0 |
| B. socotrana subsp. aspleniifolia | BSATD | Taida’ah (Ditwah mountain) | 67 ± 9.27 | 86 ± 5.35 | 64 ± 11.58 | 82 ± 9.15 | 2 ± 2.45 | 9 ± 3.74 | 22 ± 6 | 0 ± 0 |
| B. socotrana subsp. aspleniifolia | BSATR | Tarabah | 70 ± 15.17 | 78 ± 14.9 | 69 ± 17.15 | 77 ± 17.29 | 1 ± 2 | 18 ± 11.66 | 11 ± 7.35 | 0 ± 0 |
| B. socotrana subsp. socotrana | BSSA | Ayhaft | 16 ± 10.2 | 45 ± 24.19 | 16 ± 10.2 | 45 ± 24.19 | 2 ± 2.45 | 10 ± 11.4 | 65 ± 10.49 | 7 ± 6.78 |
| B. socotrana subsp. socotrana | BSSH | Hamadarah (Homhil) | 32 ± 10.3 | 54 ± 15.43 | 25 ± 10 | 43 ± 15.47 | 5 ± 6.32 | 8 ± 5.1 | 40 ± 11.83 | 15 ± 7.07 |
| B. socotrana subsp. socotrana | BSSZ | Zarkan (Asmin) | 82 ± 5.1 | 83 ± 6.69 | 76 ± 3.74 | 77 ± 4.86 | 0 ± 0 | 15 ± 6.32 | 1 ± 2 | 2 ± 2.45 |
References
- Bongers, F.; Groenendijk, P.; Bekele, T.; Birhane, E.; Damtew, A.; Decuyper, M.; Eshete, A.; Gezahgne, A.; Girma, A.; Khamis, M.A.; et al. Frankincense in peril. Nat. Sustain. 2019, 2, 602–610. [Google Scholar] [CrossRef]
- Maděra, P.; Paschová, Z.; Ansorgová, A.; Vrškový, B.; Lvončík, S.; Habrová, H. Volatile compounds in oleo-gum resin of Socotran species of Burseraceae. Acta Univ. Agric. Silvic. Mendel. Brun. 2017, 65, 73–90. [Google Scholar] [CrossRef] [Green Version]
- Ogbazghi, W. The Distribution and Regeneration of Boswellia papyrifera (Del.) Hochst. in Eritrea. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2001; p. 131. [Google Scholar]
- Ogbazghi, W.; Bongers, F.; Rijkers, T.; Wessel, M. Population structure and morphology of the frankincense tree Boswellia papyrifera along an altitude gradient in Eritrea. J. Drylands 2006, 1, 85–94. [Google Scholar]
- Gebrehiwot, K.; Muys, F.B.; Haile, M.; Mitloehner, R. Introducing Boswellia papyrifera (Del.) Hochst. and its non-timber forest product, frankincense. Int. For. Rev. 2003, 5, 348–353. [Google Scholar] [CrossRef]
- Negussie, A.; Aerts, R.; Gebrehiwot, K.; Muys, B. Seedling mortality causes recruitment limitation of Boswellia papyrifera in northern Ethiopia. J. Arid Environ. 2008, 72, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Sabo, P.; Ouédraogo, A.; Salako, K.V.; Glèlè Kakaï, R. Land use impacts on Boswellia dalzielii Hutch., an African frankincense tree in Burkina Faso. Bois Forêts Trop. 2021, 349, 51–63. [Google Scholar] [CrossRef]
- Attorre, F.; Taleb, N.; De Sanctis, M.; Farcomeni, A.; Guillet, A.; Vitale, M. Developing conservation strategies for endemic tree species when faced with time and data constraints: Boswellia spp. on Socotra (Yemen). Biodivers. Conserv. 2011, 20, 1483–1499. [Google Scholar] [CrossRef]
- Lvončík, S.; Vahalík, P.; Bongers, F.; Peijenburg, J.; Hušková, K.; Van Rensburg, J.J.; Hamdiah, S.; Maděra, P. Development of Boswellia elongata Balf. F., population in Homhil protected area, Soqotra. Rend. Lincei Sci. Fis. Nat. 2020, 31, 747–759. [Google Scholar] [CrossRef]
- Thulin, M. The Genus Boswellia (Burseraceae): The Frankincense Trees; Acta Universitatis Upsaliensis: Uppsala, Sweden, 2020. [Google Scholar]
- Miller, A.G.; Morris, M. Ethnoflora of the Soqotra Archipelago; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2004. [Google Scholar]
- Lvončík, S.; Řepka, R. Boswellia socotrana: One or two taxa? Novon 2020, 28, 17–23. [Google Scholar] [CrossRef]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable land use management needed to conserve the dragon´s blood tree of Socotra Island, a vulnerable endemic umbrella species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef] [Green Version]
- Swartout, B.T.; Solowey, E. Increasing Boswellia sacra seeds’ germination viability and genetic variability utilizing various methods. Net J. Agric. Sci. 2018, 6, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Eslamieh, J. Creating “Perfect” Boswellia. Cactus Succul. J. 2010, 82, 126–131. [Google Scholar] [CrossRef]
- Eshete, A.; Teketay, D.; Lemenih, M.; Bongers, F. Effects of resin tapping and tree size on the purity, germination and storage behavior of Boswellia papyrifera (Del.) Hochst. seeds from Metema District, northwestern Ethiopia. For. Ecol. Manag. 2012, 269, 31–36. [Google Scholar] [CrossRef]
- Ouédraogo, A.; Thiombiano, A. Regeneration pattern of four threatened tree species in Sudanian savannas of Burkina Faso. Agrofor. Syst. 2012, 86, 35–48. [Google Scholar] [CrossRef]
- Rijkers, T.; Ogbazghi, W.; Wessel, M.; Bongers, F. The effect of tapping for frankincense on sexual reproduction in Boswellia papyrifera. J. Appl. Ecol. 2006, 43, 1188–1195. [Google Scholar] [CrossRef]
- Gebre Medhin, T.G.; Negash, L. The Effect of Different Pre-Sowing Seed Treatments on the Germination of Boswellia papyrifera, a Key Dryland Tree; Final Report; Ethiopian Society of Soil Science: Addis Ababa, Ethiopia, 1999. [Google Scholar]
- Adam, A.A.; El Tayeb, A.M. A comparative study of natural regeneration of B. papyrifera and other tree species in Jebel Marra Darfur; Sudan. Res. J. Agric. Biol. Sci. 2008, 4, 94–102. [Google Scholar]
- Savithramma, N.; Ankanna, S.; Bhumi, G. Effect of nanoparticles on seed germination and seedling growth of Boswellia ovalifoliolata an endemic and endangered medicinal tree taxon. Nano Vis. 2020, 2, 2. [Google Scholar]
- De Souza, A.C.; de Oliveira, L.M.; Souza, G.F.; Schmidt, S.S.; Liesch, P.P. Causes of low seed quality in Ilex paraguariensis A. St. Hil. samples (Aquifoliaceae). Floresta Ambient. 2020, 27, e20170960. [Google Scholar] [CrossRef]
- Wang, K.S. Relationship between empty seed and genetic factors in European beech (Fagus sylvatica L.). Silva Fenn. 2003, 37, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, M.; Shupp, E.W. Empty seeds reduce seed predation by birds in Juniperus osteosperma. Evol. Ecol. 1998, 12, 823–827. [Google Scholar] [CrossRef]
- Raju, A.J.S.; Lakshmi, P.V.; Ramana, K.V. Entomophily, ornithophily and anemochory in the highly self-incompatible Boswellia ovalifoliolata Bal. & Henry (Burseraceae), an endemic and endangered medicinal tree species. J. Threat. Taxa 2012, 4, 2673–2684. [Google Scholar] [CrossRef] [Green Version]
- Sunnichan, V.G.; Mohan Ran, H.Y.; Shivanna, K.R. Reproductive biology of Boswellia serrata, the source of salai guggul, an important gum-resin. Bot. J. Linn. Soc. 2005, 147, 73–82. [Google Scholar] [CrossRef] [Green Version]
- García, C.; Vasconcelos, R. The beauty and the beast: Endemic mutualistic interactions promote community-based conservation on Socotra Island (Yemen). J. Nat. Conserv. 2017, 35, 20–23. [Google Scholar] [CrossRef]
- Scholte, P.; De Geest, P. The climate of Socotra Island (Yemen). A first-time assessment of the timing of the monsoon wind reversal and its influence on precipitation and vegetation patterns. J. Arid Environ. 2010, 74, 1507–1515. [Google Scholar] [CrossRef]
- Kalivodová, H.; Culek, M.; Čermák, M.; Maděra, P.; Habrová, H. Potential importance of Socotra dragon’s blood tree cloud forests and woodlands for capturing horizontal precipitation. Rend. Lincei Sci. Fis. Nat. 2020, 31, 607–621. [Google Scholar] [CrossRef]
- Abiyu, A.; Bongers, F.; Eshete, A.; Gebrehiwot, K.; Kindu, M.; Lemenih, M.; Moges, Y.; Ogbazghi, W.; Sterck, F.J. Incense woodlands in Ethiopia and Eritrea: Regeneration problems and restoration possibilities. In Degraded Forests in Eastern Africa; Routledge: Abingdon, UK, 2010; pp. 144–163. [Google Scholar]
- Bantihun, A.; Tesema, T. Regeneration status of Acacia polyacantha and Boswellia papyrifera species in Shimelegir Forest, Jawi District, Ethiopia. J. Nat. Sci. Res. 2018, 8, 27–36. [Google Scholar] [CrossRef]
- Groenendijk, P.; Eshete, A.; Sterck, F.J.; Zuidema, P.A.; Bongers, F. Limitations to sustainable frankincense production: Blocked regeneration, high adult mortality and declining populations. J. Appl. Ecol. 2012, 49, 164–173. [Google Scholar] [CrossRef]
- Teshome, M.; Eshete, A.; Bongers, F. Uniquely regenerating frankincense tree populations in western Ethiopia. For. Ecol. Manag. 2017, 389, 127–135. [Google Scholar] [CrossRef]
- Tolera, M.; Sass-Klaassen, U.; Eshete, A.; Bongers, F.; Sterck, F.J. Frankincense tree recruitment failed over the past half century. For. Ecol. Manag. 2013, 304, 65–72. [Google Scholar] [CrossRef]
- Adam, A.A.; Osman, A.A. Sprouting capacity of Boswellia papyrifera (Del.) Hochst in Jebel Mrarra Area, Darfur; Sudan: Effect of stump diameter and height. Res. J. Agric. Biol. Sci. 2008, 4, 51–57. [Google Scholar]
- Abiyu, A.; Dejene, T.; Eshete, A.; Sisay, K. Vegetative propagation of Boswellia papyrifera: Time of collection and propagule size affect survival and establishment. J. Arid Environ. 2016, 133, 122–124. [Google Scholar] [CrossRef]
- Haile, G.; Gebrehiwot, K.; Lemenih, M.; Bongers, F. Time of collection and cutting sizes affect vegetative propagation of Boswellia papyrifera (Del.) Hochst through leafless branch cuttings. J. Arid Environ. 2011, 75, 873–877. [Google Scholar] [CrossRef]
- Lvončík, S.; Maděra, P.; Volařík, D.; Vrškový, B.; Habrová, H. First proposal of seed regions for frankincense trees (Boswellia spp.) on Socotra Island. J. Landsc. Ecol. 2013, 6, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Niebler, J.; Eslamieh, J.; Buettner, A. Frankincense revisited, part II: Volatiles in rare Boswellia species and hybrids. Chem. Biodivers. 2016, 13, 630–643. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). International Rules for Seed Testing; International Seed Testing Association: Bassersdorf, Switzerland, 2020; Chapter 5; pp. 21–52. [Google Scholar]
- Czabator, F.J. Germination value: An index combining speed and completeness of pine seed germination. For. Sci. 1962, 8, 386–396. [Google Scholar]
- Attorre, F.; Van Damme, K. Twenty years of biodiversity research and nature conservation in the Socotra Archipelago (Yemen). Rend. Lincei Sci. Fis. Nat. 2020, 31, 563–569. [Google Scholar] [CrossRef]
- Maděra, P.; Van Damme, K. Socotra Archipelago (Yemen). In The Encyclopedia of Conservation; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 9780124095489. [Google Scholar] [CrossRef]











| Species | Locality | Sp. Code | Coordinates | Altitude | Date of Collection |
|---|---|---|---|---|---|
| Boswellia ameero (BA) | Ayhaft | BAA | 12.594013 N 53.988233 E | 424 | 18 May 2021 |
| Danagahan | BADN | 12.585567 N 54.052267 E | 629 | 2 June 2020, 27 May 2021 | |
| Dixam | BADX | 12.517885 N 53.930242 E | 928 | 17 May 2021 | |
| Firmihin | BAF | 12.481588 N 54.010128 E | 617 | 17 May 2021 | |
| Qatariyah | BAQ | 12.351153 N 53.708817 E | 860 | 1 June 2021 | |
| Boswellia bullata (BB) | Hazahaz | BBH | 12.657705 N 53.440213 E | 476 | 24 May 2021 |
| Shoab rock (green flowers) | BBSG | 12.62341 N 53.403162 E | 1 | 7 May 2021 | |
| Shoab (red flower) | BBSR | 12.61131 N 53.401565 E | 1 | 7 May 2021 | |
| Taida’ah (Ditwah mountain) | BBT | 12.697685 N 53.514572 E | 215 | 20 June 2020, 15 May 2021 | |
| Boswellia dioscoridis (BD) | Wadi Diasraha | BDDA | 12.419418 N 54.144430 E | 67 | 26 May 2021 |
| Wadi Difa’araha | BDDF | 12.423367 N 54.123345 E | 78 | 26 May 2021 | |
| Wadi Digasfa | BDDG | 12.455117 N 54.146743 E | 167 | 26 May 2021 | |
| Halah (Hoq cave) | BDH | 12.591258 N 54.360668 E | 70 | 27 May 2021 | |
| Sheebhan | BDS | 12.473968 N 54.004037 E | 572 | 20 May 2020, 17 May 2021 | |
| Boswellia elongata (BE) | Diburak | BEDB | 12.523304 N 54.277032 E | 360 | 7 June 2020, 3 June 2021 |
| Dishal | BEDS | 12.527823 N 51.177206 E | 282 | 26 May 2020, 2 June 2021 | |
| Homhil | BEH | 12.576047 N 54.286267 E | 335 | 1 June 2020, 10 June 2021 | |
| Makalihim | BEM | 12.505041 N 54.168744 E | 169 | 28 May 2020, 2 June 2021 | |
| Shabarah | BES | 12.524795 N 53.885643 E | 441 | 4 July 2021 | |
| Boswellia nana (BN) | Hamadarah (Homhil) | BNH | 12.593068 N 54.303272 E | 616 | 19 May 2021 |
| Boswellia popoviana (BP) | Bedgofahar | BPB | 12.657987 N 54.166502 E | 178 | 30 May 2021 |
| Sheebhan (Didrafarantan) | BPD | 12.471406 N 54.004312 E | 570 | 5 June 2021 | |
| Firmihin | BPF | 12.482612 N 54.009487 E | 608 | 17 May 2021 | |
| Klisan | BPK | 12.473802 N 54.335207 E | 270 | 19 June 2020, 10 June 2021 | |
| Momi Falang | BPV | 12.460495 N 54.364178 E | 304 | 19 June 2020, 10 June 2021 | |
| Boswellia scopulorum (BS) | Bedgofahar | BSB | 12.656790 N 54.139097 E | 149 | 20 May 2021 |
| Boswellia socotrana subsp. aspleniifolia (BSA) | Raidah (Taida’ah) | BSAB | 12.6832650 N 53.5772433 E | 346 | 6 June 2021 |
| Dinawtat | BSAN | 12.655353 N 53.683472 E | 34 | 16 May 2021 | |
| Shata Qalansiyah | BSAS | 12.629508 N 53.592948 E | 128 | 28 May 2020, 17 May 2021 | |
| Taida’ah (Ditwah mountain) | BSATD | 12.699898 N 53.515875 E | 288 | 15 May 2021 | |
| Tarabah | BSATR | 12.579373 N 53.924450 E | 128 | 9 June 2021 | |
| Boswellia socotrana subsp. socotrana (BSS) | Ayhaft | BSSA | 12.596872 N 53.989797 E | 242 | 20 May 2020, 23 June 2021 |
| Hamadarah (Homhil) | BSSH | 12.591187 N 54.301247 E | 466 | 12 June 2021 | |
| Zarkan (Asmin) | BSSZ | 12.595920 N 54.151198 E | 408 | 15 June 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdiah, S.; Karas, L.; Houšková, K.; Van Damme, K.; Attorre, F.; Vahalík, P.; Habrová, H.; Lvončík, S.; Eler, K.; Maděra, P. Seed Viability and Potential Germination Rate of Nine Endemic Boswellia Taxa (Burseraceae) from Socotra Island (Yemen). Plants 2022, 11, 1418. https://doi.org/10.3390/plants11111418
Hamdiah S, Karas L, Houšková K, Van Damme K, Attorre F, Vahalík P, Habrová H, Lvončík S, Eler K, Maděra P. Seed Viability and Potential Germination Rate of Nine Endemic Boswellia Taxa (Burseraceae) from Socotra Island (Yemen). Plants. 2022; 11(11):1418. https://doi.org/10.3390/plants11111418
Chicago/Turabian StyleHamdiah, Salem, Lukáš Karas, Kateřina Houšková, Kay Van Damme, Fabio Attorre, Petr Vahalík, Hana Habrová, Samuel Lvončík, Klemen Eler, and Petr Maděra. 2022. "Seed Viability and Potential Germination Rate of Nine Endemic Boswellia Taxa (Burseraceae) from Socotra Island (Yemen)" Plants 11, no. 11: 1418. https://doi.org/10.3390/plants11111418