
Genomic-Wide Identification and Characterization of the Uridine Diphosphate Glycosyltransferase Family in Eucommia ulmoides Oliver


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of Putative UGTs in E. ulmoides
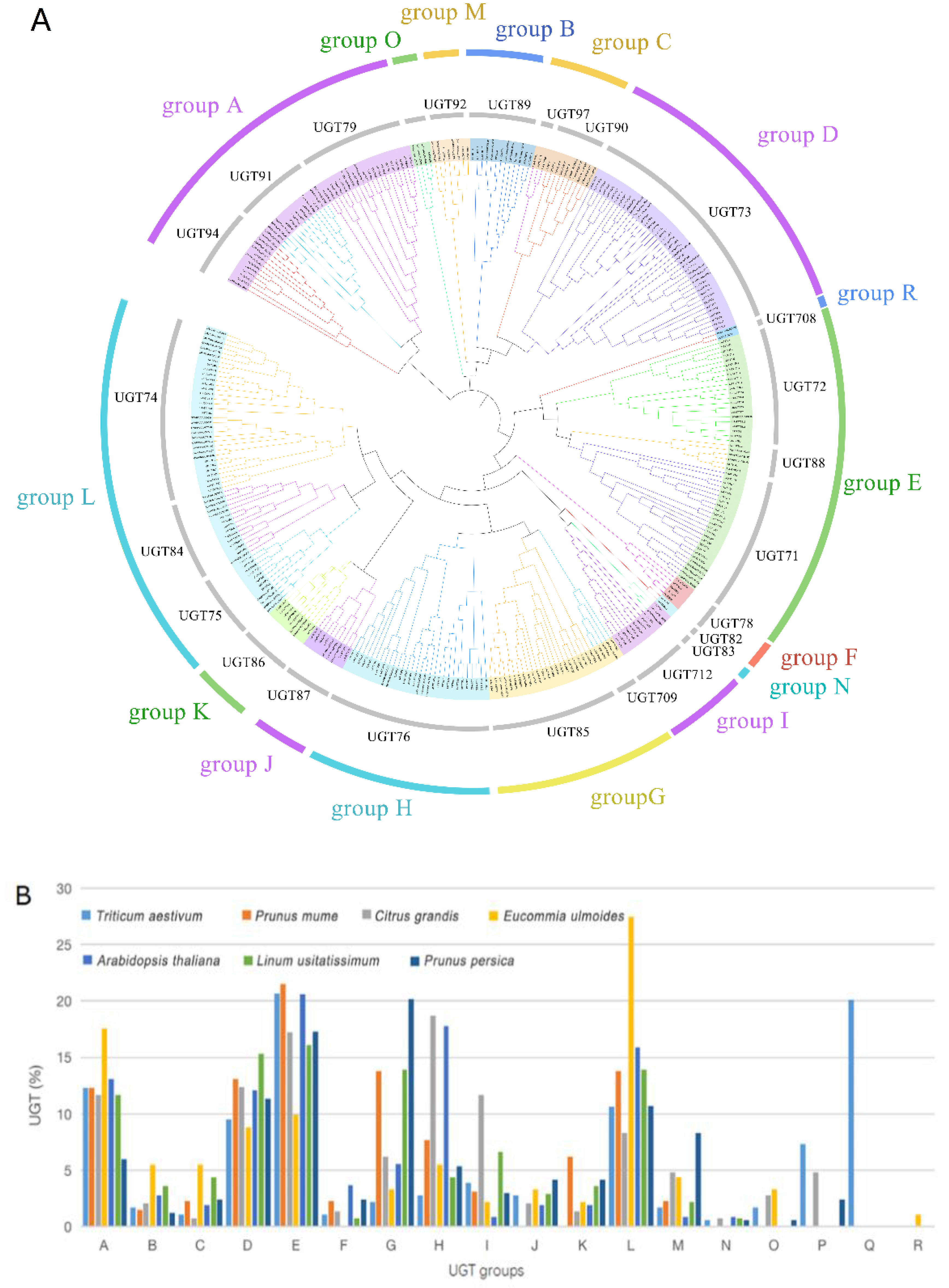
2.2. Phylogenetic Analysis of EuUGTs
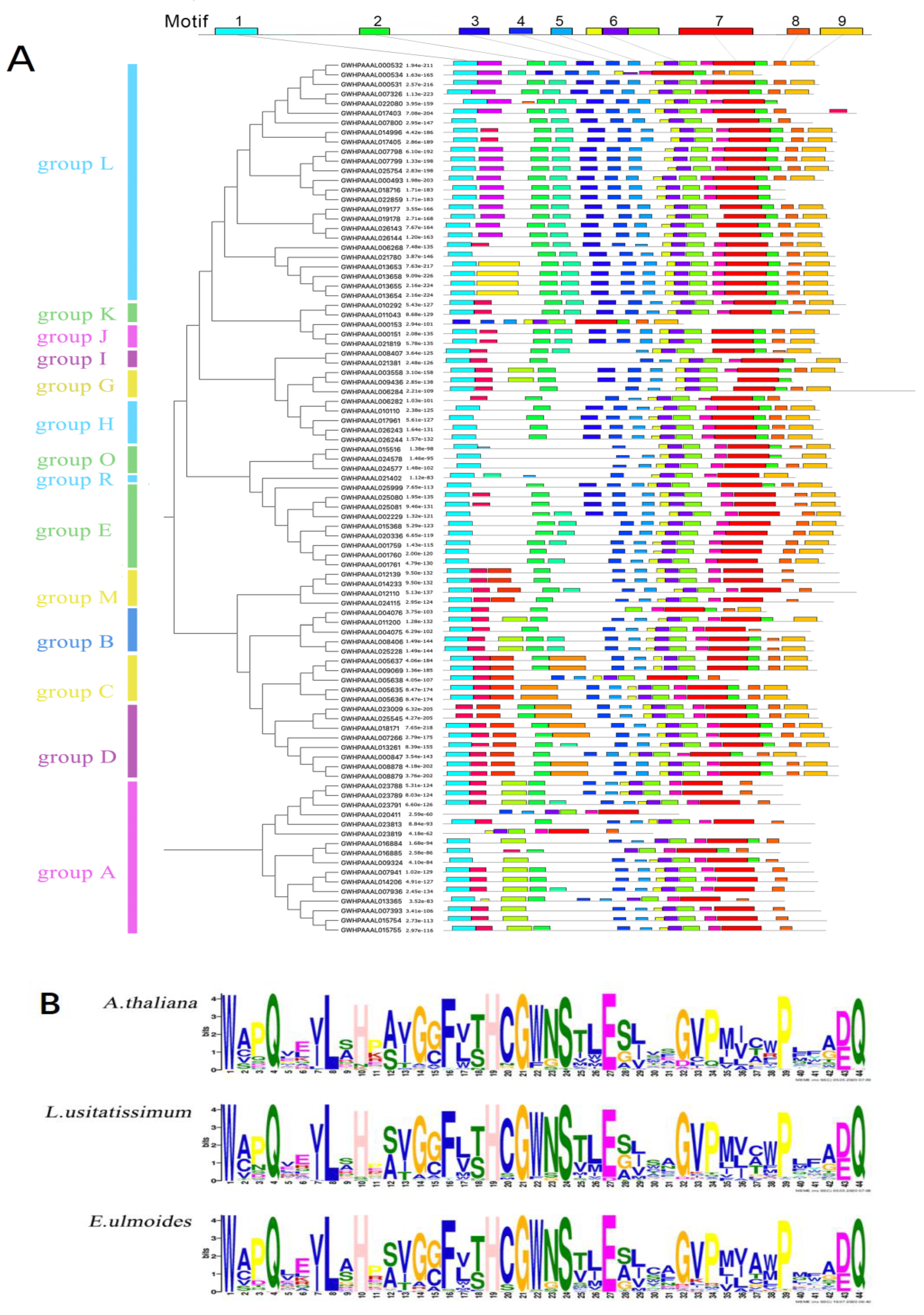
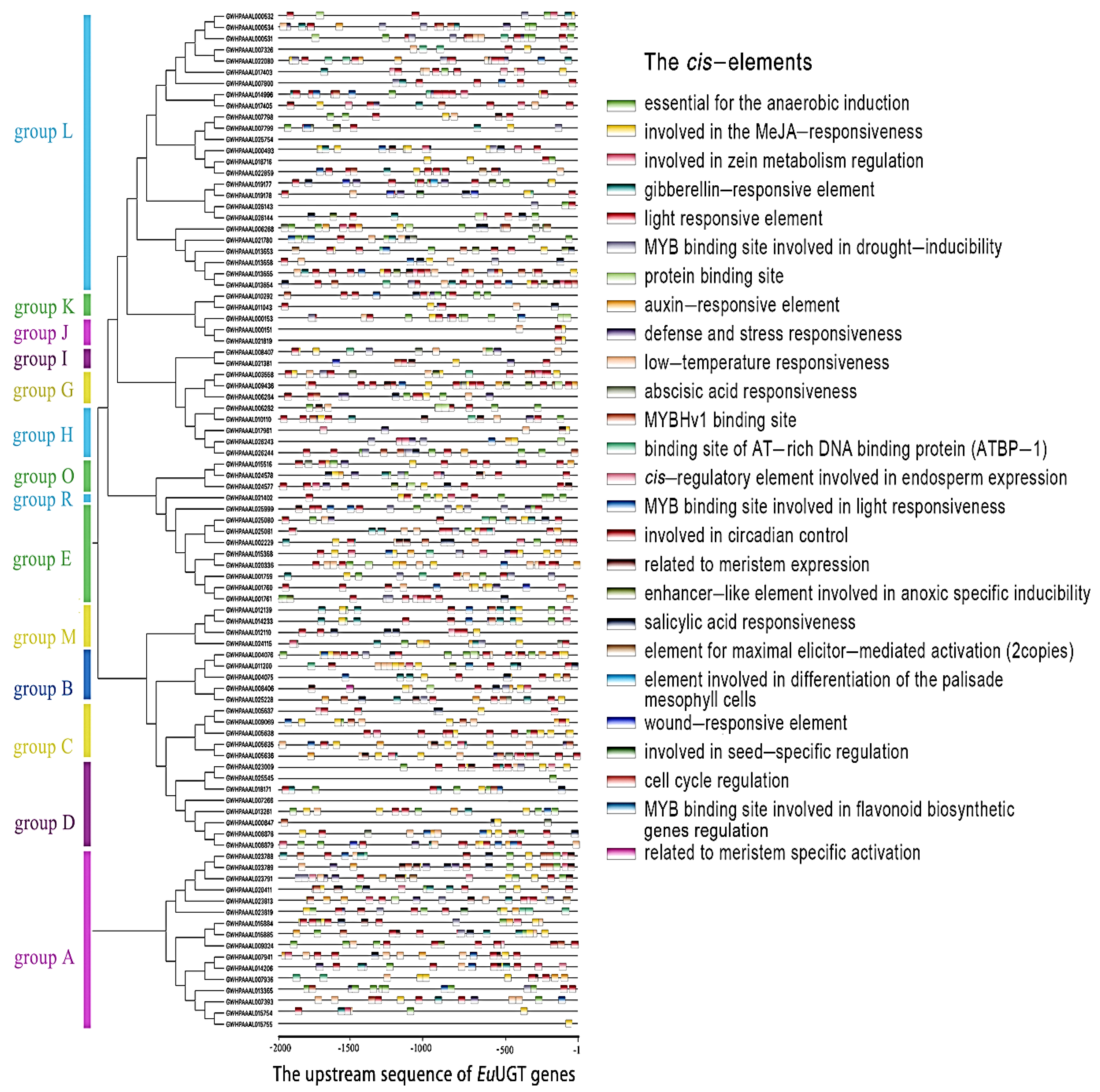
2.3. Analysis of Conserved Motifs and Prediction of Cis-Acting Elements
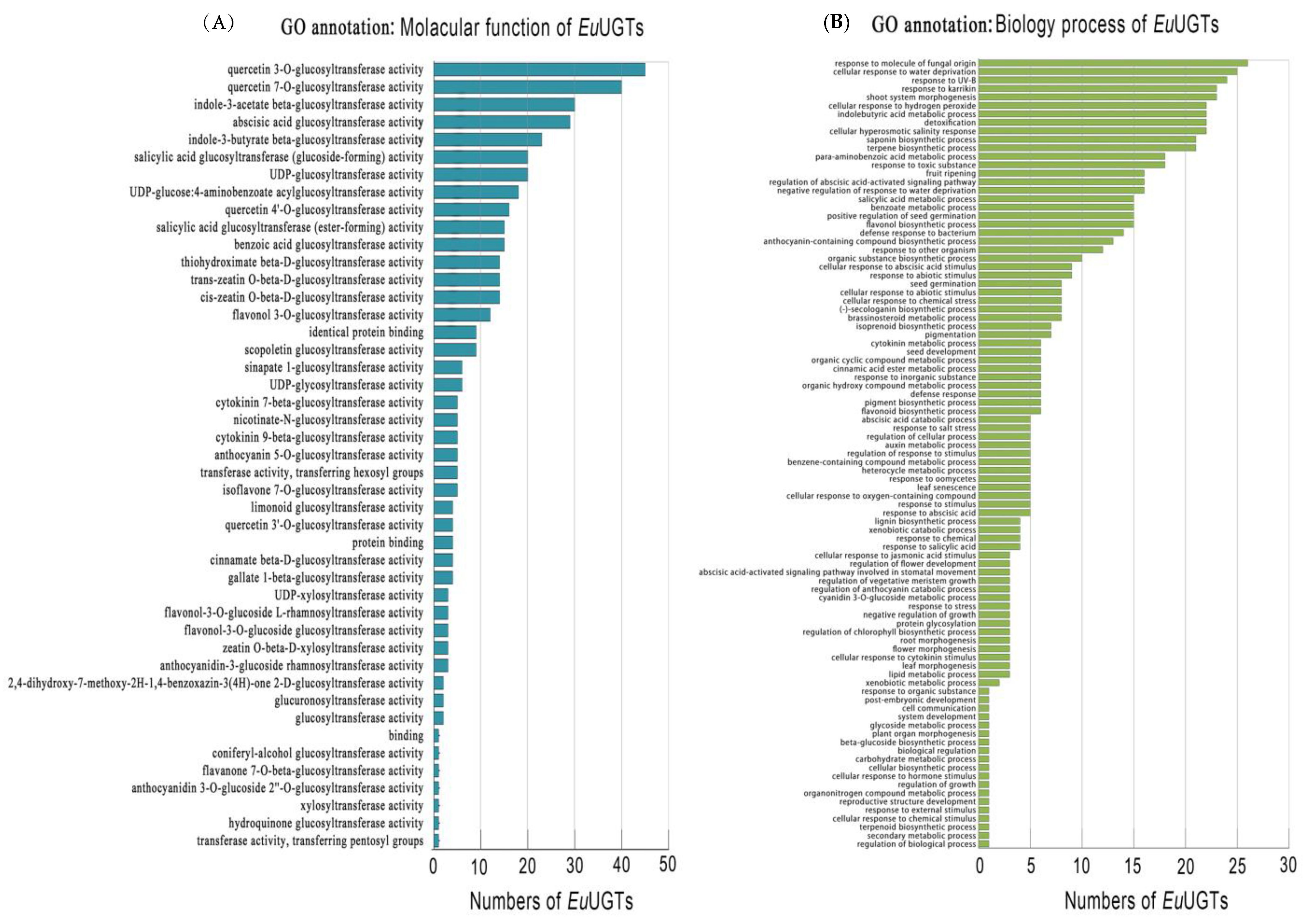
2.4. GO Annotation of EuUGTs
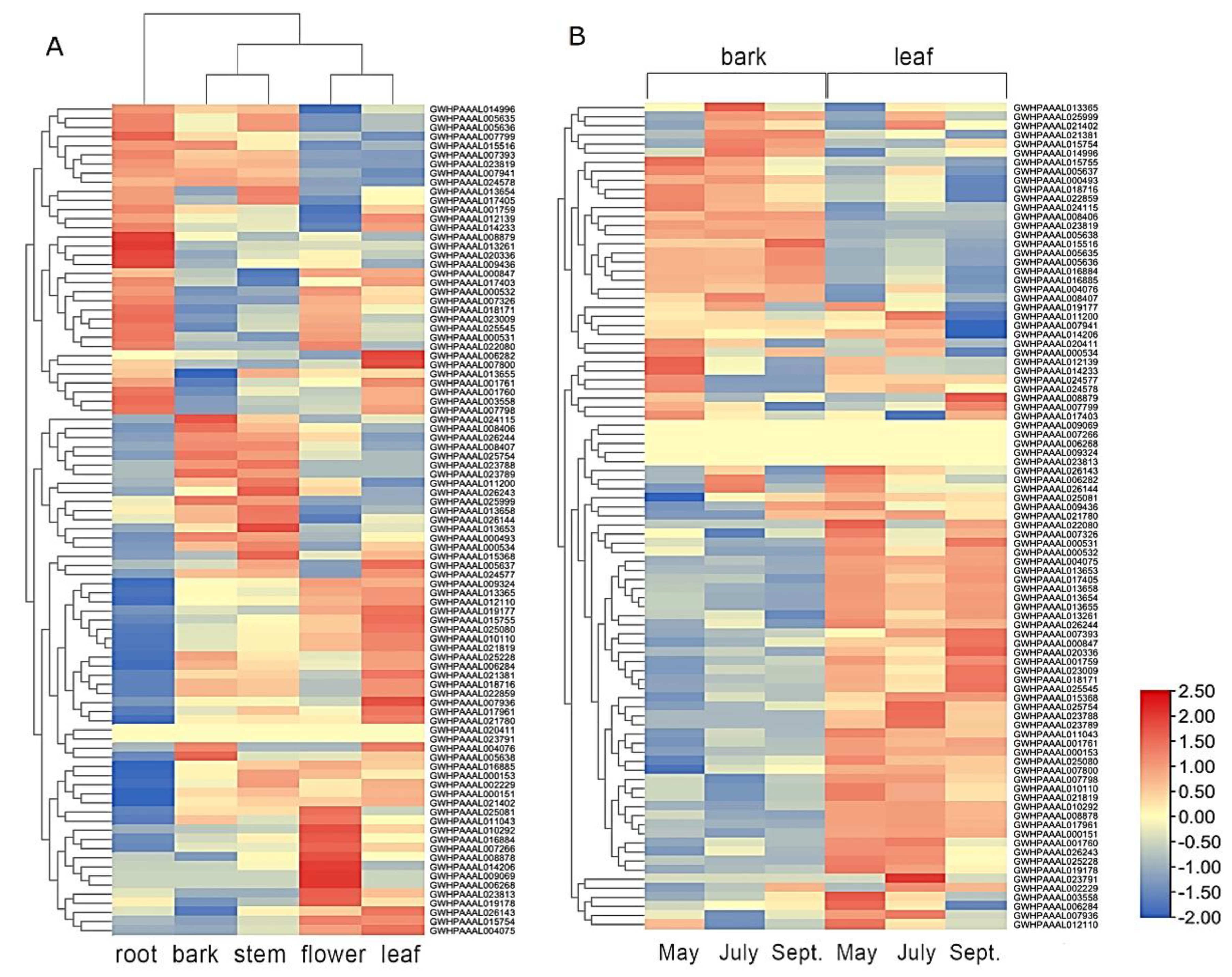
2.5. Transcriptome Analysis for Organ-Specific and Time-Specific Gene Expression
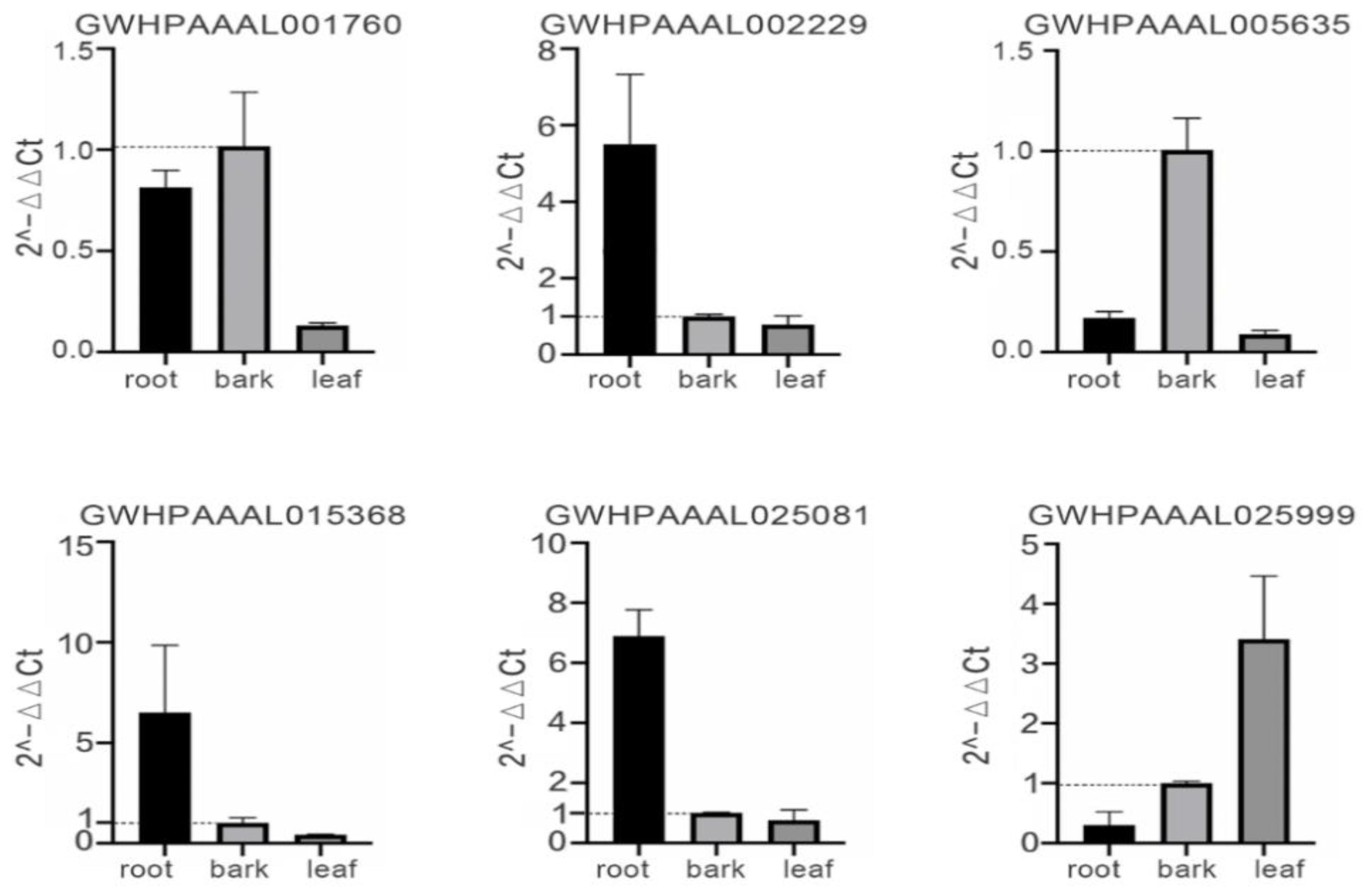
2.6. RT-qPCR Analysis of the Selected EuUGT Genes
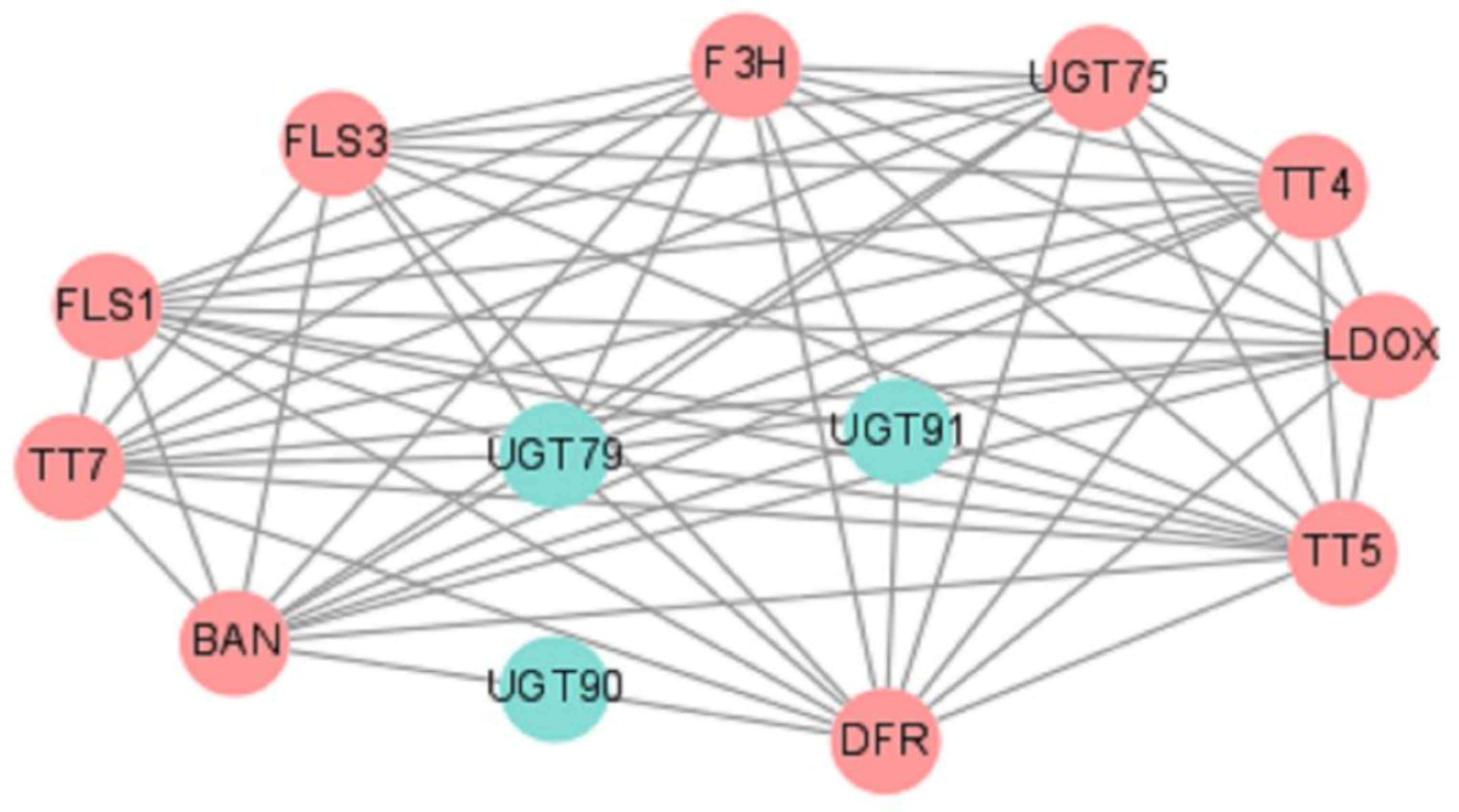
2.7. Construction of Protein–Protein Interaction (PPI) Network of EuUGTs
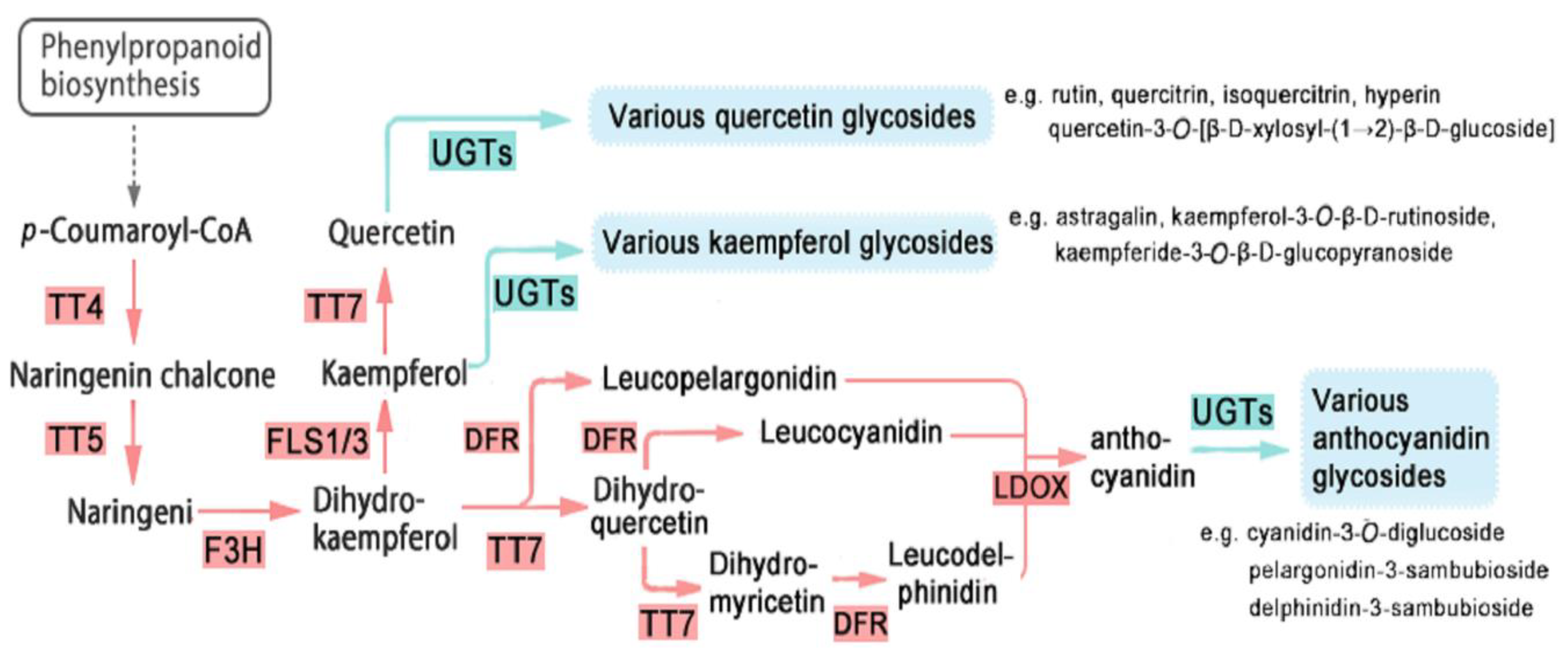
3. Discussion
4. Materials and Methods
4.1. Data Resources Used
4.2. Genome-Wide Identification of Putative UGTs in E. ulmoides
4.3. Prediction of the Cellular Location, Physical and Chemical Properties of EuUGTs
4.4. Phylogenetic Analysis of UGTs among A. thaliana, L. usitatissimum and E. ulmoides
4.5. Analysis of Conserved Motifs and Prediction of Cis-Acting Elements
4.6. GO Annotation of EuUGTs
4.7. Transcriptome Analysis for Organ-Specific and Season-Specific Gene Expression
4.8. Quantitative Real-Time PCR Analysis of the Selected EuUGT Genes
4.9. Construction of Protein–Protein Interaction (PPI) Network of EuUGTs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities. Biochem. J. 1997, 326, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, P.M.; Deleury, E.; Davies, G.J.; Henrissat, B. An evolving hierarchical family classification for glycosyltransferases. J. Mol. Biol. 2003, 328, 307–317. [Google Scholar] [CrossRef]
- Rahimi, S.; Kim, J.; Mijakovic, I.; Jung, K.H.; Choi, G.; Kim, S.C.; Kim, Y.J. Triterpenoid-biosynthetic UDP-glycosyltransferases from plants. Biotechnol. Adv. 2019, 37, 107394. [Google Scholar] [CrossRef] [PubMed]
- le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant Sci. 2016, 7, 735. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, P.I.; Owens, I.S.; Burchell, B.; Bock, K.W.; Bairoch, A.; Bélanger, A.; Fournel-Gigleux, S.; Green, M.; Hum, D.W.; Iyanagi, T.; et al. The UDP glycosyltransferase gene superfamily: Recommended nomenclature update based on evolutionary divergence. Pharmacogenet 1997, 7, 255–269. [Google Scholar] [CrossRef]
- Li, Y.; Baldauf, S.; Lim, E.K.; Bowles, D.J. Phylogenetic analysis of the UDP-glycosyltransferase multigene family of Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 4338–4343. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Li, P.; Wang, Y.; Dong, R.R.; Yu, H.M.; Hou, B.K. Genome-wide identification and phylogenetic analysis of family-1 UDP glycosyltransferase in maize (Zea mays). Planta 2014, 239, 1265–1279. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Ahmad, D.; Zhang, X.; Zhang, Y.; Wu, L.; Jiang, P.; Ma, H. Genome-wide analysis of family-1 UDP glycosyltransferases (UGT) and identification of UGT genes for FHB resistance in wheat (Triticum aestivum L.). BMC Plant Biol. 2018, 18, 67. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Hanada, K. An evolutionary view of functional diversity in family 1 glycosyltransferases. Plant J. 2011, 66, 182–193. [Google Scholar] [CrossRef]
- Barvkar, V.T.; Pardeshi, V.C.; Kale, S.M.; Kadoo, N.Y.; Gupta, V.S. Phylogenomic analysis of UDP glycosyltransferase 1 multigene family in Linum usitatissimum identified genes with varies expression patterns. BMC Genom. 2012, 13, 175. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.E.; Tian, L. Phylogenomic analysis of UDP-dependent glycosyltransferases provides insights into the evolutionary landscape of glycosylation in plant metabolism. Plant J. 2019, 100, 1273–1288. [Google Scholar] [CrossRef] [PubMed]
- Caputi, L.; Malnoy, M.; Goremykin, V.; Nikiforova, S.; Martens, S. A genome-wide phylogenetic reconstruction of family 1 UDP-glycosyltransferases revealed the expansion of the family during the adaptation of plants to life on land. Plant J. 2012, 69, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.B.; Jayakodi, M.; Lee, Y.S.; Nguyen, V.B.; Park, H.S.; Koo, H.J.; Choi, I.Y.; Kim, D.H.; Chung, Y.J.; Ryu, B.; et al. Identification of candidate UDP-glycosyltransferases involved in protopanaxadiol-type ginsenoside biosynthesis in Panax ginseng. Sci. Rep. 2018, 8, 11744. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Yun, X.; Zheng, X.; Xu, H.; Zhan, R.; Chen, W.; Xu, Y.; Chen, Y.; Zhang, J. Transcriptomic comparison reveals candidate genes for triterpenoid biosynthesis in two closely related Ilex Species. Front. Plant Sci. 2017, 8, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Yao, S.; Dai, X.; Yin, Q.; Liu, Y.; Jiang, X.; Wu, Y.; Qian, Y.; Pang, Y.; Gao, L.; et al. Identification of UDP-glycosyltransferases involved in the biosynthesis of astringent taste compounds in tea (Camellia sinensis). J. Exp. Bot. 2016, 8, 2285–2297. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Tang, L.; He, J.W.; Li, J.; Wang, Y.Z. Ethnobotany, phytochemistry and pharmacological properties of Eucommia ulmoides: A review. Am. J. Chin. Med. 2019, 47, 259–300. [Google Scholar] [CrossRef]
- Wang, J.Y.; Chen, X.J.; Zhang, L.; Pan, Y.Y.; Gu, Z.X.; Yuan, Y. Anti-inflammatory effects of Eucommia ulmoides Oliv.: Male flower extract on lipopolysaccharide-induced inflammation. Chin. Med. J. 2019, 132, 319–328. [Google Scholar] [CrossRef]
- Lang, C.; Liu, Z.; Taylor, H.W.; Baker, D.G. Effect of Eucommia ulmoides on systolic blood pressure in the spontaneous hypertensive rat. Am. J. Chin. Med. 2005, 33, 215–230. [Google Scholar] [CrossRef]
- Wu, D.; Yu, D.; Zhang, Y.; Dong, J.; Li, D.; Wang, D. Metabolite profiles, bioactivity, and HPLC fingerprint of different varieties of Eucommia ulmoides Oliv.: Towards the utilization of medicinal and commercial Chinese endemic tree. Molecules 2018, 23, 1898. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Pan, H.; Zhang, X.; Wang, H.; Liu, S.; Zhang, H.; Qian, H.; Wang, L.; Ying, H. Geniposide improves glucose homeostasis via regulating foxo1/pdk4 in skeletal muscle. J. Agric. Food. Chem. 2019, 67, 4483–4492. [Google Scholar] [CrossRef]
- Ma, Z.G.; Kong, C.Y.; Song, P.; Zhang, X.; Yuan, Y.P.; Tang, Q.Z. Geniposide protects against obesity-related cardiac injury through Ampkalpha- and Sirt1-dependent mechanisms. Oxid. Med. Cell Longev. 2018, 2018, 6053727. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Li, P.; Duan, J.X.; Liu, T.; Guan, X.X.; Mei, W.X.; Liu, Y.P.; Sun, G.Y.; Wan, L.; Zhong, W.J.; et al. Aucubin alleviates bleomycin-induced pulmonary fibrosis in a mouse model. Inflammation 2017, 40, 2062–2073. [Google Scholar] [CrossRef]
- Chang, I.M. Liver-protective activities of aucubin derived from traditional oriental medicine. Res. Comm. Mol. Pathol. Pharmacol. 1998, 102, 189–204. [Google Scholar] [CrossRef]
- Yao, J.; Zou, Z.; Wang, X.; Ji, X.; Yang, J. Pinoresinol diglucoside alleviates oxLDL-induced dysfunction in human umbilical vein endothelial cells. Evid. Based Complement. Alternat. Med. 2016, 2016, 3124519. [Google Scholar] [CrossRef]
- Li, Y.J.; Dong, X.; Zhao, D.G. Cloning and sequence analysis of glucosyltransferase gene EuCGT1 in Eucommia ulmoides Olive. J. Mount. Agri. Biol. 2014, 33, 27–35. [Google Scholar] [CrossRef]
- Leng, Y.; Liu, S.H.; Zhao, Y.C.; Zhao, D.G. Prokaryotic expression and protein purification of EUGT2 gene from Eucommia ulmoides. Mol. Plant Breed. 2020, 18, 3240–3245. [Google Scholar] [CrossRef]
- Baruah, P.M.; Krishnatreya, D.B.; Bordoloi, K.S.; Gill, S.S.; Agarwala, N. Genome wide identification and characterization of abiotic stress response lncRNA in Capsicum annuum. Plant Physiol. Biochem. 2021, 162, 221–236. [Google Scholar] [CrossRef]
- Kumar, M.; Kherawat, B.S.; Dey, P.; Saha, D.; Singh, A.; Bhatia, S.K.; Ghodake, G.S.; Kadam, A.A.; Kim, H.-U.; Manorama; et al. Genome-wide identification and characterization of PIN-FORMED (PIN) gene family reveals role in developmental and various stress conditions in Triticum aestivum L. Int. J. Mol. Sci. 2021, 22, 7396. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kherawat, B.S.; Singh, A.; Dey, P.; Kabi, M.; Debnath, D.; Saha, D.; Khandual, A.; Manorama, S.R.; Ali, A.; et al. Genome-wide identification and characterization of the brassinazole-resistant (BZR) gene family and its expression in the various developmental stage and stress conditions in wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2021, 22, 8743. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, B.; Zhou, W.; Xie, L.; Wang, L.; Zhang, Y.Z.Q. Genome-wide identification and expression analysis of the AT-hook Motif Nuclear Localized gene family in soybean. BMC Genom. 2021, 22, 361. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, H.; Liu, J.; Du, Q.; Lu, S.; Liu, C. Genome-wide identification and functional characterization of natural antisense transcripts in Salvia miltiorrhiza. Sci. Rep. 2021, 11, 4769. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Fang, Y.-X.; Zhang, Z.; Zheng, J.; Zhang, X.; Li, J.; Niu, C.; Xue, D.; Zhang, X. Genome-wide identification and expression pattern analysis of the KCS gene family in barley. Plant Growth Reg. 2021, 93, 89–103. [Google Scholar] [CrossRef]
- Wuyun, T.; Wang, L.; Liu, H.; Wang, X.; Zhang, L.; Bennetzen, J.L.; Li, T.; Yang, L.; Liu, P.; Du, L.; et al. The hardy rubber tree genome provides insights into the evolution of polyisoprene biosynthesis. Mol. Plant. 2018, 11, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus x domestica borkh). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, M.L.F.; Rodriguez, E.; Casas, M.I.; Labadie, G.; Grotewold, E.; Casati, P. Identification of a bifunctional maize C- and O-glucosyltransferase. J. Biol. Chem. 2013, 288, 31678–31688. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, Z.; Yang, J.; Mutanda, I.; Li, S.; Zhang, Q.; Zhang, Y.; Zhang, Y.; Wang, Y. Pathway-specific enzymes from bamboo and crop leaves biosynthesize anti-nociceptive c-glycosylated flavones. Comm. Biol. 2020, 3, 110. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.; Li, Y.; Lim, E.K. Higher plant glycosyltransferases. Genome Biol. 2001, 2, reviews3004.1–3004.6. [Google Scholar] [CrossRef]
- Shao, H.; He, X.; Achnine, L.; Blount, J.W.; Dixon, R.A.; Wang, X. Crystal structures of a multifunctional triterpene/flavonoid glycosyltransferase from Medicago truncatula. Plant Cell 2005, 17, 3141–3154. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Hou, H.; Hu, J.; Liu, B. Optimized microwave extraction, characterization and antioxidant capacity of biological polysaccharides from Eucommia ulmoides Oliver leaf. Sci. Rep. 2018, 8, 6561. [Google Scholar] [CrossRef] [Green Version]
- Lan, J.Y.; Liu, Z.; Zhang, Q.; Pei, Z.C.; Li, B.H. A study on growth characteristics of Eucommia ulmoides Oliver in different areas and the dynamic changes of main effective components in leaves. J. Agric. Univ. Hebei 2019, 42, 51–56. [Google Scholar] [CrossRef]
- Wang, L.N.; Li, W.; Qin, J.P.; Yang, M.H. Study on dynamic variation of main contents in different parts of Eucommia ulmoides. Chin. Pharm. 2009, 18, 29–31. [Google Scholar] [CrossRef]
- Guerriero, G.; Berni, R.; Munoz-Sanchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.A.; Cantini, C.; Cai, G.; Hausman, J.F.; et al. Production of plant secondary metabolites: Examples, tips and suggestions for biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satake, H.; Koyama, T.; Bahabadi, S.E.; Matsumoto, E.; Ono, E.; Murata, J. Essences in metabolic engineering of lignan biosynthesis. Metabolites 2015, 5, 270–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.-M.; Cheng, W.-W.; Song, X.; Long, Y.-H.; Xing, Z.-B. Genome-wide identification and expression analysis of glycosyltransferase gene family 1 in Quercus robur L. J. Appl. Genet. 2021. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Tsukamoto, C.; Ishimoto, M. Reconstruction of the evolutionary histories of UGT gene superfamily in legumes clarifies the functional divergence of duplicates in specialized metabolism. Int. J. Mol. Sci. 2020, 21, 1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masada, S.; Terasaka, K.; Mizukami, H. A single amino acid in the PSPG-box plays an important role in the catalytic function of CaUGT2 (Curcumin glucosyltransferase), a group D family 1 glucosyltransferase from Catharanthus roseus. FEBS Lett. 2007, 581, 2605–2610. [Google Scholar] [CrossRef] [Green Version]
- Hansen, E.H.; Osmani, S.A.; Kristensen, C.; Møller, B.L.; Hansen, J. Substrate specificities of family 1 UGTs gained by domain swapping. Phytochemistry 2009, 70, 473–482. [Google Scholar] [CrossRef]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Ludwig-Muller, J.; Chung, G.; Ahmad, M.O.; Yang, S.H.; Lee, S.I. Comparative genomic and transcriptomic analysis of family-1 UDP glycosyltransferase in three Brassica species and Arabidopsis indicates stress-responsive. Sci. Rep. 2018, 8, 1875. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Rawat, V.; Suresh, C.G. Genome-wide identification and tissue-specific expression analysis of UDP-glycosyltransferases genes confirm their abundance in Cicer arietinum (Chickpea) genome. PLoS ONE 2014, 9, e109715. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Muhammad, A.; Li, G.; Zhang, J.; Cheng, J.; Qiu, J.; Jiang, T.; Jin, Q.; Cai, Y.; Lin, Y. Family-1 UDP glycosyltransferase in pear (Pyrus bretschneideri): Molecular identification, phylogenomic characterization and expression profiling during stone cell formation. Mol. Biol. Rep. 2019, 46, 2153–2175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-C.; Peng, X.-X.; Lu, Y.-B.; Wu, X.-X.; Chen, L.-W.; Feng, H. Genome-wide association study reveals the genes associated with the leaf inclusion contents in Chinese medical tree Eucommia ulmoides. Biosci. Biotech. Biochem. 2021, 85, 233–241. [Google Scholar] [CrossRef]
- Beejmohun, V.; Fliniaux, O.; Hano, C.; Pilard, S.; Grand, E.; Lesur, D.; Cailleu, D.; Lamblin, F.; Laine, E.; Kovensky, J.; et al. Coniferin dimerisation in lignan biosynthesis in flax cells. Phytochemistry 2007, 68, 2744–2752. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.; Kim, G.D.; Kang, W.; Park, J.E.; Kim, S.H.; Choe, E.; Kim, J.I.; Auh, J.H. Pinoresinol diglucoside is screened as a putative alpha-glucosidase inhibiting compound in Actinidia arguta leaves. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 473–479. [Google Scholar] [CrossRef]
- Do, K.H.; Choi, Y.W.; Kim, E.K.; Yun, S.J.; Kim, M.S.; Lee, S.Y.; Ha, J.M.; Kim, J.H.; Kim, C.D.; Son, B.G.; et al. Pinoresinol-4,4’-di-o-beta-d-glucoside from Valeriana officinalis root stimulates calcium mobilization and chemotactic migration of mouse embryo fibroblasts. Phytomedicine 2009, 16, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, X.; Li, Y.; Yan, Y.; Yuan, Q. Metabolic engineering of Escherichia coli for microbial synthesis of monolignols. Metab. Eng. 2017, 39, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Davin, L.B.; Lewis, N.G. An historical perspective on lignan biosynthesis: Monolignol, allylpenol and hydroxycinnamic acid coupling and downstream metabolism. Phytochemistry 2003, 2, 257–288. [Google Scholar] [CrossRef]
- Satake, H.; Ono, E.; Murata, J. Recent advances in the metabolic engineering of lignan biosynthesis pathways for the production of transgenic plant-based foods and supplements. J. Agric. Food Chem. 2013, 61, 11721–11729. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, J.; Ni, Y.; Liu, Y.; Gao, Z.; Zhao, X.; Gao, Z. Tracing the mass flow from glucose and phenylalanine to pinoresinol and its glycosides in Phomopsis sp. XP-8 using stable isotope assisted TOF-MS. Sci. Rep. 2019, 9, 18495. [Google Scholar] [CrossRef] [PubMed]
- Schmid, G.; Grisebach, H. Enzymic synthesis of lignin precursors. Purification and properties of UDPglucose: Coniferyl-alcohol glucosyltransferase from cambial sap of spruce (Picea abies L.). Eur. J. Biochem. 1982, 123, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Ono, E.; Kim, H.J.; Murata, J. Molecular and functional characterization of novel furofuran class lignan glucosyltransferases from Forsythia. Plant Biotech. 2010, 27, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Okazawa, A.; Kusunose, T.; Ono, E.; Kim, H.J.; Muranaka, T. Glucosyltransferase activity of Arabidopsis UGT71c1 towards pinoresinol and lariciresinol. Plant Biotechnol. 2015, 5, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.W.; Wang, W.C.; Jin, S.H.; Hou, B.K. Over-expression of a putative poplar glycosyltransferase gene, PtGT1, in tobacco increases lignin content and causes early flowering. J. Exp. Bot. 2012, 63, 2799–2808. [Google Scholar] [CrossRef]
- Lin, J.S.; Huang, X.X.; Li, Q.; Cao, Y.; Bao, Y.; Li, X.F.M.Y.J.; Fu, C.; Hou, B.K. UDP-glycosyltransferase 72B1 catalyzes the glucose conjugation of monolignols and is essential for the normal cell wall lignification in Arabidopsis thaliana. Plant J. 2016, 88, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Guo, F.F.; Xiao, J.P.; Wei, J.Y.; Tang, L.Y.; Yang, H.J. Research advances in chemical constituents and pharmacological activities of different parts of Eucommia ulmoides. Chin. J. Chin. Mater. Med. 2020, 3, 497–512. [Google Scholar] [CrossRef]
- Yan, Y.; Zhao, H.; Zou, L.S.; Liu, X.H.; Chai, C.; Wang, S.N.; Jiao, Y.J. Difference of chemical constituents in different medicinal parts of Eucommia ulmoides by LC-Triple TOF MS/MS. J. Chin. Mass Spectr. Soc. 2018, 39, 101–111. [Google Scholar] [CrossRef]
- Wu, B.; Liu, X.; Zhang, B. Genome-wide characterization, evolution and expression profiling of UDP-glycosyltransferase family in pomelo (Citrus grandis) fruit. BMC Plant Biol. 2020, 20, 459. [Google Scholar] [CrossRef]
- Lewis, D.R.; Ramirez, M.V.; Miller, N.D.; Vallabhaneni, P.; Ray, W.K.; Winkel, R.F.H.B.S.J.; Muday, G.K. Auxin and ethylene induce flavonol accumulation through distinct transcriptional networks. Plant Physiol. 2011, 156, 144–164. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Gao, L.; Gao, J.; Xu, Y.; Liu, H.; Cao, X.; Zhang, B.; Chen, K. Genome-wide identification, expression patterns, and functional analysis of UDP glycosyltransferase family in peach (Prunus persica L. Batsch). Front. Plant Sci. 2017, 8, 389. [Google Scholar] [CrossRef]
- Wang, P.; Wei, Y.; Fan, Y.; Liu, Q.; Wei, W.; Yang, C.; Zhang, L.; Zhao, G.; Yue, J.; Yan, X.; et al. Production of bioactive ginsenosides RH2 and RG3 by metabolically engineered yeasts. Metab. Eng. 2015, 29, 97–105. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. Hmmer web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, V.; Kosarev, P.; Seledsov, I.; Vorobyev, D. Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 2006, 7 (Suppl. 1), S10.1–S10.12. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-seq data. Nat. Biotech. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totawa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. Weblogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the blast2go suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with hisat2 and hisat-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome assembly from long-read RNA-seq alignments with stringtie2. Genom. Biol. 2019, 20, 278. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. Tbtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chen, J. Comparison of methods of RNA extraction with Eucommia ulmoides. J. For. Eng. 2007, 21, 19–21. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, D.; Wang, L.-C.; Tang, T.; Feng, H. Genomic-Wide Identification and Characterization of the Uridine Diphosphate Glycosyltransferase Family in Eucommia ulmoides Oliver. Plants 2021, 10, 1934. https://doi.org/10.3390/plants10091934
Ouyang D, Wang L-C, Tang T, Feng H. Genomic-Wide Identification and Characterization of the Uridine Diphosphate Glycosyltransferase Family in Eucommia ulmoides Oliver. Plants. 2021; 10(9):1934. https://doi.org/10.3390/plants10091934
Chicago/Turabian StyleOuyang, Dan, Lan-Chun Wang, Ting Tang, and Hong Feng. 2021. "Genomic-Wide Identification and Characterization of the Uridine Diphosphate Glycosyltransferase Family in Eucommia ulmoides Oliver" Plants 10, no. 9: 1934. https://doi.org/10.3390/plants10091934