
Antifungal Activity of the Extract of a Macroalgae, Gracilariopsis persica, against Four Plant Pathogenic Fungi



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling from G. persica
2.2. Extraction to Assess Antifungal Activity
2.3. Preparation of Methanolic Extract for HPLC and GC-MS Analysis
2.4. Analysis of Methanol Extract by Gas Chromatograph-Mass Spectrometry (GC-MS)
2.5. Measurement of Phenol Compounds by the HPLC Method
2.6. Preparation of Potato Dextrose Agar (PDA)
2.7. In Vitro Antifungal Activity of the Extract
2.8. Fungistatic or Fungicide Effect of the Extract
2.9. Data Analysis
3. Results
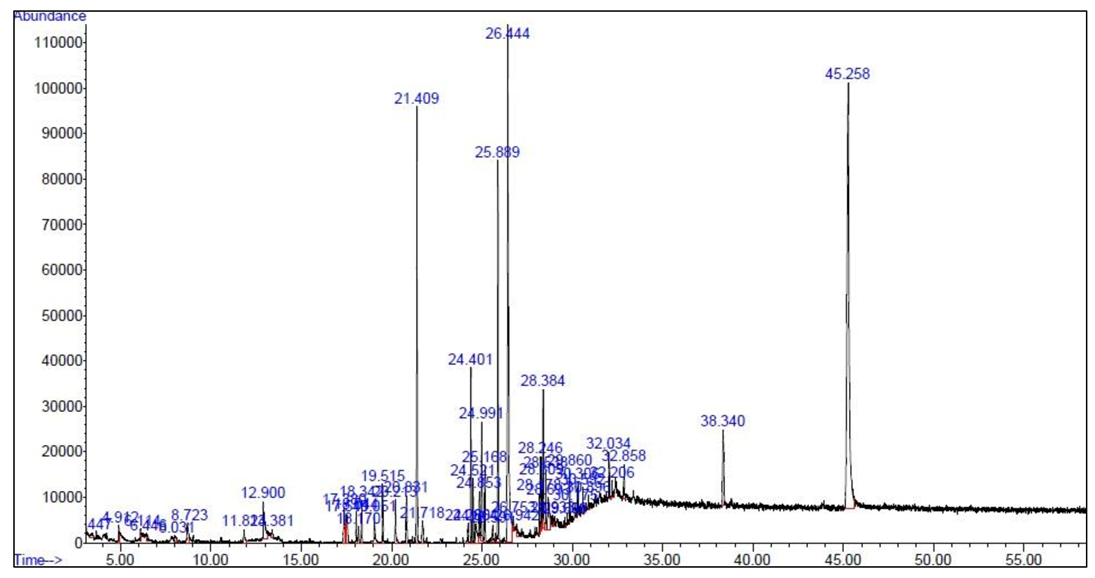
3.1. Results of GC-MS
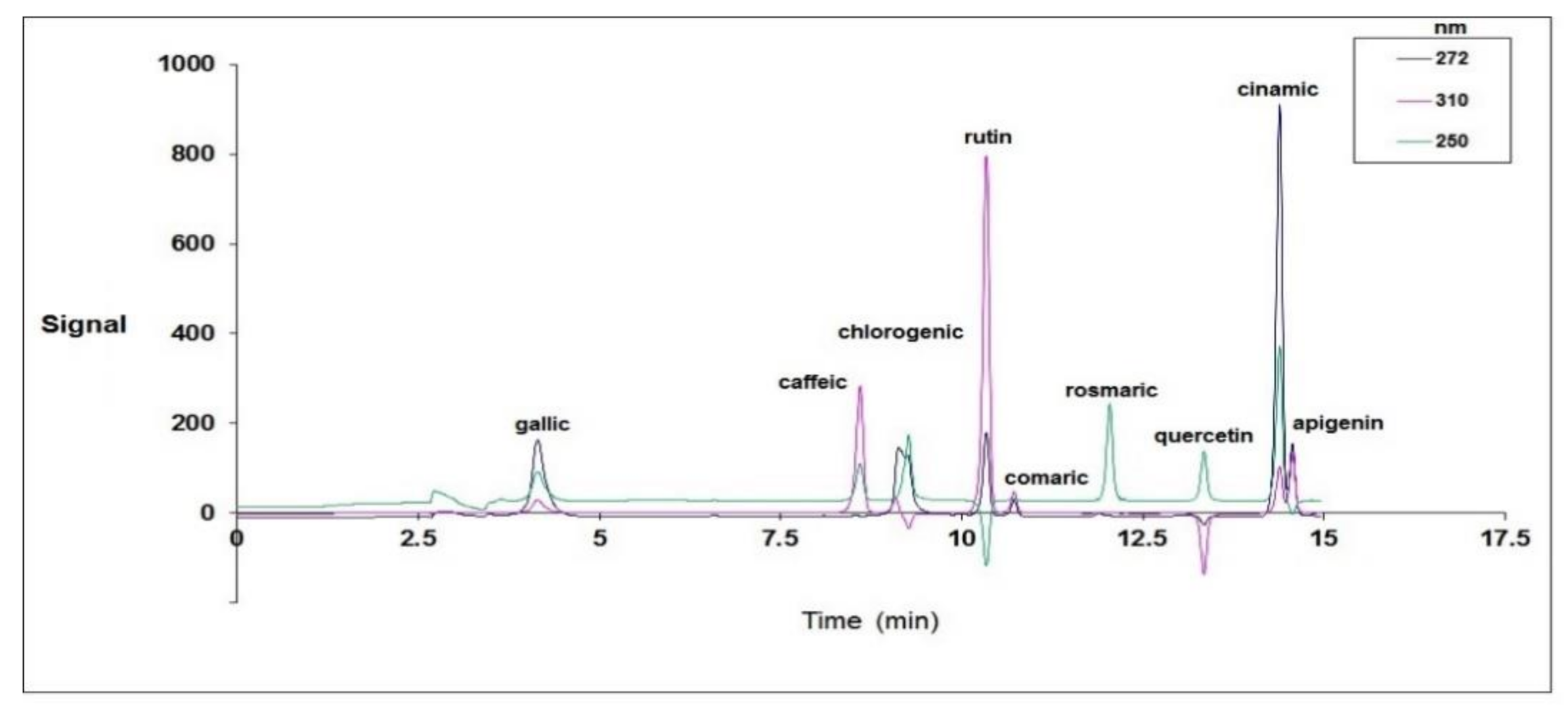
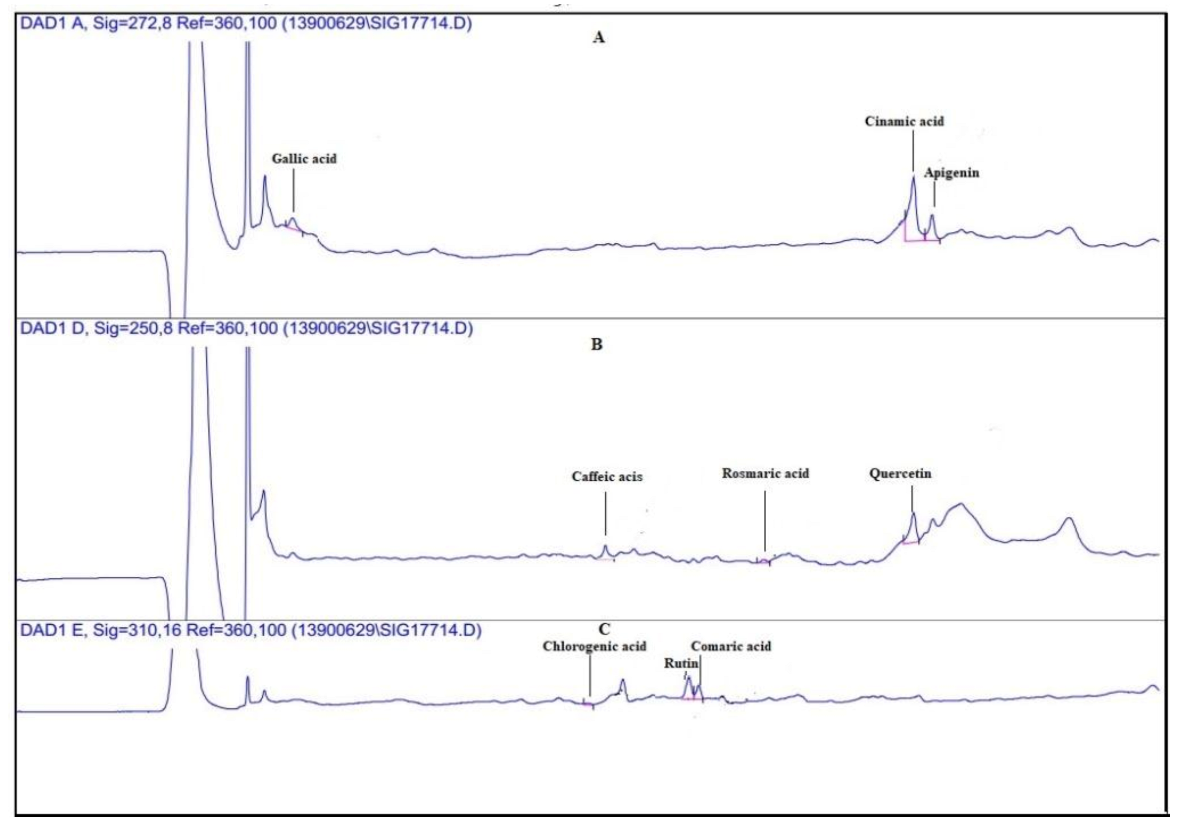
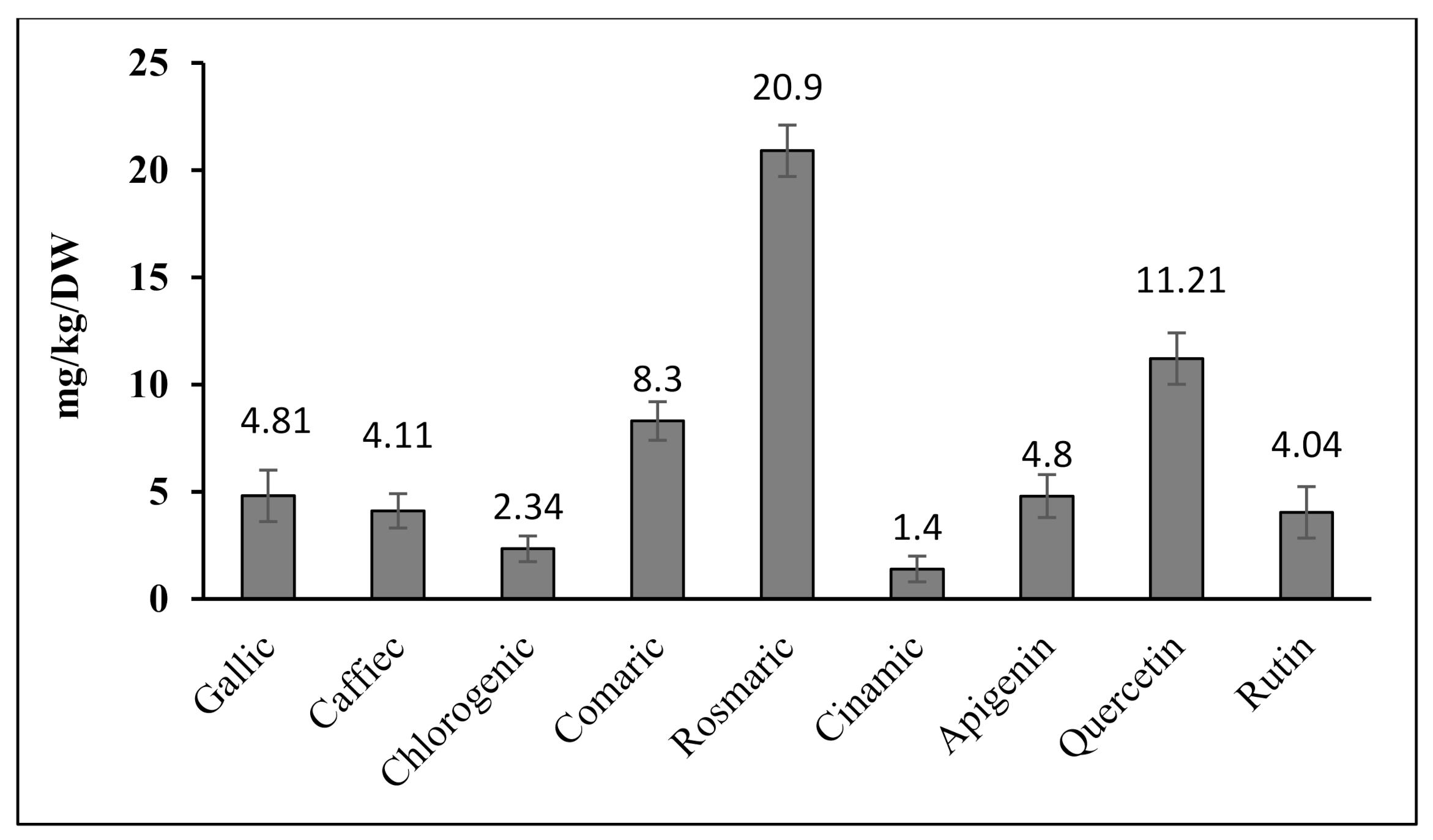
3.2. Quantitative Analysis of Phenolic Compounds Determined Using HPLC
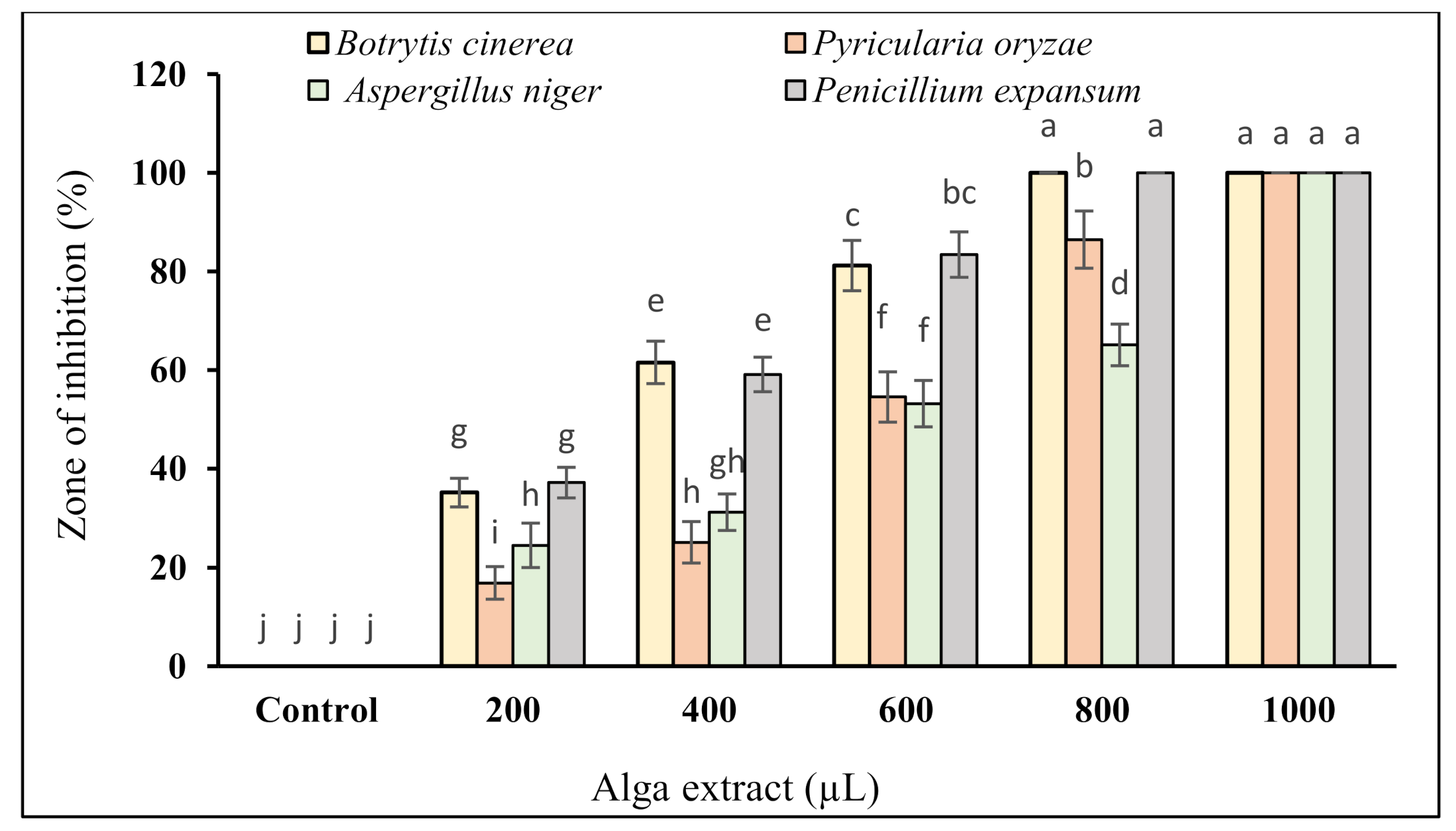
3.3. Antifungal Activity
Fungistatic
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, R.E. Phycology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Mishra, V.K.; Temelli, F.; Ooraikul, B.; Shacklock, P.F.; Craigie, J.S. Lipids of the red alga, Palmaria palmata. Bot. Mar. 1993, 36, 169–174. [Google Scholar] [CrossRef]
- Mohamed, F.; Namitha, K.K.; Chidambara Murthy, K.N.; Mahadeva Swamy, M.; Sarada, R.; Khanam, S.; Subbarao, P.V.; Ravishankar, G.A. Chemical composition, iron bioavailability and antioxidant activity of Kappsphycus alvarezi. J. Agric. Chem. 2005, 53, 792–797. [Google Scholar]
- Plaza, M.; Santoyo, S.; Jaime, L.; Reina, G.G.B.; Herrero, M.; Senorans, F.J.; Lbanez, E. Screening for bioactive compounds from algae. J. Pharmac. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Al-Saif, S.S.A.; Abd-Alraouf, N.; El-Wazanani, H.A.; Aref, I.A. Antibacterial substances from marine algae isolated from Jeddah coast of Red sea, Saudi Arabia. Sau. J. Biolog. Sci. 2014, 14, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Righini, H.; Roberti, R.; Baraldi, E. Use of algae in strawberry management. J. Appl. Phyco. 2018, 30, 3551–3564. [Google Scholar] [CrossRef]
- Soares, A.R.; Robaina, M.C.S.; Mendes, G.S.; Silva, T.S.L.; Gestinari, L.M.S.; Pamplona, O.S.; Yoneshigue-Valentin, Y.; Kaiser, C.R.; Romanos, M.T.V. Antiviral activity of extracts from Brazilian seaweeds against herpes simplex virus. Rev. Bras. Farmacogn. Braz. J. Pharmacogn. 2012, 22, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Sagar, A.; Sayyed, R.Z.; Ramteke, P.W.; Sharma, S.; Marraiki, N.; Elgorban, A.E.; Syed, A. ACC deaminase and antioxidant enzymes producing halophilic Enterobacter sp. PR14 promotes the growth of rice and millets under salinity stress. Physiol. Mol. Biol. Plants 2020, 26, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; Enshasy, H.E. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; El Enshasy, H.; Gafur, A.; et al. Bacterial Plant Biostimulants: A Sustainable Way towards Improving Growth, Productivity, and Health of Crops. Sustainability 2021, 21, 2856. [Google Scholar] [CrossRef]
- Saharan, V.; Sharma, G.; Yadav, M.; Choudhary, M.K.; Sharma, S.S.; Pal, A.; Biswas, P. Synthesis and in vitro antifungal efficacy of Cu–chitosan nanoparticles against pathogenic fungi of tomato. Int. J. Biol. Macromol. 2015, 75, 346–353. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant. Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lee, S.; Wang, J.; Ma, J.; Bianco, T.; Jia, Y. Current advances on genetic resistance to rice blast disease. In Rice-Germplasm, Genetics, and Improvement; Yan, W., Bao, J., Eds.; InTech: London, UK, 2014; pp. 195–217. [Google Scholar]
- Bautista-Baños, S. Postharvest Decay: Control Strategies; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2014; p. 383. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer Science + Business Media, LLC: New York, NY, USA, 2009; p. 519. [Google Scholar]
- Palencia, E.R.; Hinton, D.M.; Bacon, C.W. The black Aspergillus species of maize and peanuts and their potential for myctoxin production. Toxins 2010, 2, 399–416. [Google Scholar] [CrossRef]
- Ziani, K.; Fernandez-Pan, I.; Royo, M.; Maté, J.I. Antifungal activity of films and solutions based on chitosan against typical seed fungi. Food Hydrocolloids 2009, 23, 2309–2314. [Google Scholar] [CrossRef]
- Kwon, J.H.; Jeong, S.G.; Hong, S.B.; Chae, Y.S.; Park, C.S. Occurrence of blue mold on sweet persimmon (Diospyros kaki) caused by Penicillium expansum. Res. Plant Dis. 2006, 12, 290–293. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.L.; Zhang, Q.; Sun, X.Y.; Jiang, L.; Zhang, R.; Sun, G.Y.; Zha, Y.L.; Biggs, A.R. Etiology of moldy core, core browning, and core rot of Fuji apple in China. Plant Dis. 2013, 97, 510–516. [Google Scholar] [CrossRef] [Green Version]
- Tannous, J.; Atoui, A.; El Khoury, A.; Francis, Z.; Oswald, I.P.; Puel, O.; Lteif, R. A study on the physicochemical parameters for Penicillium expansum growth and patulin production: Effect of temperature, pH, and water activity. Food Sci. Nutr. 2016, 4, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; Enshasy, D.; Dailin, J.; Suriani, N.L. Recent understanding of soil Acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Arora, H.; Sharma, A.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z.; Datta, R. Pythium damping-off and root rot of Capsicum annuum L.: Impacts, Diagnosis, and Management. Microorganisms 2021, 9, 823. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: A review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, B.S.; Jabran, K.; Mahajan, G. Rice Production Worldwide; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2017; p. 563. [Google Scholar]
- Raven, P.H.; Evert, R.F.; Eichom, S.E. Biology of Plants, 5th ed.; Worth Publishers: New York, NY, USA, 1992. [Google Scholar]
- Saidani, K.; Bedjou, F.; Benabdesslam, F.; Touati, N. Antifungal activity of methanolic extracts of four Algerian marine algae species. Afr. J. Biotechnol. 2012, 11, 9496–9500. [Google Scholar] [CrossRef]
- Sahoo, D.; Yarish, C. Mariculture of seaweeds. In Phycological Methods: Algal Culturing Techniques; Andersen, R., Ed.; Academic Press: New York, NY, USA, 2005; pp. 219–237. [Google Scholar]
- Sohrabipour, J.; Rabii, R. A list of marine algae of sea shores of the Persian Gulf and Oman Sea in the Hormozgan province. Iran. J. Bot. 1999, 8, 131–162. [Google Scholar]
- Reshma, P.; Naik, M.K.; Aiyaz, M.; Niranjana, S.R.; Chennappa, G.; Shaikh, S.S.; Sayyed, R.Z. Induced systemic resistance by 2,4diacetylphloroglucinol positive fluorescent Pseudomonas strains against rice sheath blight. Indian J. Exp. Biol. 2018, 56, 207–212. [Google Scholar]
- Zope, V.P.; Jadhav, H.P.; Sayyed, R.Z. Neem cake carrier prolongs shelf life of biocontrol fungus Trichoderma viridae. Indian J. Exp. Biol. 2019, 57, 372–375. [Google Scholar]
- Liu, S. Study on the commercial cultivation of Gracilaria in south. Chi. J. Oce. Lim. 1987, 5, 281–283. [Google Scholar]
- Kumar, C.S.; Dronamraju, V.; Sarada, L.; Ramasamy, R. Seaweed extracts control the leaf spot disease of the medicinal plant Gymnema sylvestre. Ind. J. Sci. Technol. 2008, 1, 1–5. [Google Scholar] [CrossRef]
- Seal, T. Quantitative HPLC analysis of phenolic acids, flavonoids, and ascorbic acid in four different solvent extracts of two wild edible leaves, Sonchus arvensis and Oenanthe linearis of North-Eastern region in India. J. Appl. Pharmacol. Sci. 2016, 6, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Shahi, S.K.; Patra, M.; Shukla, A.C.; Dikshit, A. Use of essential oil as botanical-pesticide against postharvest spoilage in Malus pumilo fruits. Biocontrol 2003, 48, 223–232. [Google Scholar] [CrossRef]
- Husain, A.; Mumtaz, A.; Siddiqui, N. Synthesis, reactions and biological activity of 3-arylidene-5-(4-methylphenyl)-2(3H)-furanones. J. Serb. Chem. Soc. 2009, 74, 103–115. [Google Scholar] [CrossRef]
- Mansur Ali, S.; Ahmed Khan, N.; Sagathevan, K.; Anwar, A.; Siddiqui, R. Biologically active metabolite(s) from haemolymph of red-headed centipede Scolopendra subspinipes possess broad spectrum antibacterial activity. AMB Express 2019, 9, 1–17. [Google Scholar]
- Paerl, R.W.; Bertrand, E.M.; Rowland, E.; Schatt, P.; Mehiri, M.; Niehaus, T.D.; Hanson, A.D.; Riemann, L.; Bouget, F.Y. Carboxythiazole is a key microbial nutrient currency and critical component of thiamin biosynthesis. Sci. Rep. 2018, 8, 8876. [Google Scholar] [CrossRef] [PubMed]
- Magi, G.; Marini, E.; Facinelli, B. Antimicrobial activity of essential oils and carvacrol, and synergy of carvacrol and erythromycin, against clinical, erythromycin-resistant Group A Streptococci. Front. Microbiol. 2015, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, P.I.A.M.; Wan Ahmad, W.Y.; Ibrahim, N.; Abu Bakar, M. Antibacterial activity and major constituents of polyalthia cinnamomea basic fraction. Sains Malays. 2018, 47, 2063–2071. [Google Scholar] [CrossRef]
- Vyas, D.H.; Tala, S.D.; Akbari, J.D.; Dhaduk, M.F.; Joshi, H.S. Synthesis, antimicrobial, and antitubercular activity of some cyclohexenone and indazole derivatives. Ind. J. Chem. 2009, 48, 1405–1410. [Google Scholar]
- Mujeeb, F.; Bajpai, P.; Pathak, N. Phytochemical evaluation, antimicrobial activity, and determination of bioactive components from leaves of Aegle marmelos. Biol. Med. Res. Int. 2014, 2014, 1–11. [Google Scholar]
- Al-Bari, M.A.A.; Sayeed, M.A.; Rahman, M.S.; Mossadik, M.A. Characterization and antimicrobial activities of a phthalic acid derivative produced by Streptomyces bangladeshiensis—A novel species collected in Bangladesh. Res. J. Medic. Med. Sci. 2006, 1, 77–81. [Google Scholar]
- Uma, M.; Jothinayaki, S.; Kumaravel, S.; Kalaiselvi, P. Determination of bioactive components of Plectranthus amboinicus Lour by GC–MS analysis. N.Y. Sci. J. 2011, 4, 66. [Google Scholar]
- Singh, H.B.; Handique, A.K. Antifungal activity of the essential oil of Hyptis suaveolens and its efficacy in biocontrol measures in combination with Trichoderma harzianum. J. Essent. Oil Res. 2007, 9, 683–687. [Google Scholar] [CrossRef]
- Kim, Y.W.; Chun, H.J.; Kim, I.; Liu, H.; Shick Ahn, W. Antimicrobial and antifungal effects of green tea extracts against microorganisms causing vaginitis. Food Sci. Biotechnol. 2013, 22, 713–719. [Google Scholar] [CrossRef]
- Nikitina, L.E.; Startseva, V.A.; Dorofeeva, L.; Artemova, N.P.; Kuznetsov, I.V.; Lisovskaya, S.A.; Glushko, N.P. Antifungal activity of bicylic monoterpenoids and terpeneslfides. Chem. Nat. Comp. 2010, 46, 28–32. [Google Scholar] [CrossRef]
- Kumara Swamy, M.; Arumugam, G.; Kaur, R.; Ghasemzadeh, A.; Yusoff, M.M.; Sinniah, U.R. GC-MS-based metabolite profiling, antioxidant and antimicrobial properties of different solvent extracts of Malaysian Plectranthus amboinicus leaves. Evidence Based Compl. Altern. Medi. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Qi, S.H.; Xu, Y.; Gao, J.; Qian, P.Y.; Zhang, S. Antibacterial and antilarval compounds from the marine bacterium Pseudomonas rhizosphaerae. Ecol. Environ. Microbiol. 2009, 59, 229–233. [Google Scholar]
- Chandrasekaran, M.; Senthilkumar, A.; Venkatesalu, V. Antibacterial and antifungal efficacy of fatty acid methyl esters from the leaves of Sesuvium portulacastrum L. Eur. Rev. Medicol. Pharmacol. Sci. 2011, 15, 775–780. [Google Scholar]
- Togashi, N.; Shiraishi, A.; Nishizaka, M.; Matsuoka, K.; Endo, K.; Hamashima, H.; Inoue, Y. Antibacterial activity of long-chain fatty alcohols against Staphylococcus aureus. Molecules 2007, 12, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pejin, B.; Savic, A.; Sokovic, M.; Glamoclija, J.; Ciric, A.; Nikolic, M. Further in vitro evaluation of the antiradical and antimicrobial activity of phytol. Nat. Prod. Res. 2014, 28, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Sinniah, U.R. A comprehensive review on the phytochemical constituents and pharmacological activities of Pogostemon cablin Benth.: An aromatic medicinal plant of industrial importance. Molecules 2015, 20, 8521–8547. [Google Scholar] [CrossRef] [Green Version]
- Shobier, A.H.; Abdel, S.A.; Barakat, G.M. GC/MS spectroscopic approach and antifungal potential of bioactive extracts produced by marine macroalgae. Egy. J. Aqu. Res. 2016, 42, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Keskin, D.; Ceyhan, N.; Ugur, A.; Durgan Dbeys, A. Antimicrobial activity and chemical constituents of West Anatolia olive (Olea europaea L.) leaves. J. Food. Env. 2012, 10, 99–102. [Google Scholar]
- Rateb, H.S.; Ahmed, H.E.; Ahmed, S.; Ihmaid, S.; Afifi, H.S. Discovery of novel phthalimide analogs: Synthesis, antimicrobial and antitubercular screening with molecular docking studies. EXCLI J. 2016, 15, 781–796. [Google Scholar]
- Saeidniaa, S.; Permehb, P.; Goharia, A.R.; Mashinchian-Moradib, A. Gracilariopsis persica from Persian Gulf contains bioactive sterols. IJPR 2012, 11, 845–849. [Google Scholar]
- Nirupama, R. Fungal disease of white muscardiane in silkworm, Bombyx mori L. Mun. Ent. Zool. 2014, 9, 870–875. [Google Scholar]
- Soliman, A.S.; Ahmed, A.Y.; Abdel-Ghafour, A.; El-Sheekh, M.; Sobhy, H. Antifungal bio-efficacy of the red algae Gracilaria confervoides extracts against three pathogenic fungi of the cucumber plant. Middle East. J. Appl. 2018, 8, 727–735. [Google Scholar]
- Kim, S.K.; Karadeniz, F. Anti-HIV activity of extracts and compounds from marine algae. Adv. Food. Nutr. Res. 2011, 64, 255–265. [Google Scholar] [PubMed]
- Zarei Jeliani, Z.; Mashjoor, S.; Soleimani, S.; Pirian, K.; Sedaghat, F. Antioxidant activity and cytotoxicity of organic extracts from three species of green macroalgae of Ulvaceae from Persian Gulf. Modares J. Biotech. 2018, 9, 59–67. [Google Scholar]
- Abdelhamid, A.; Jouini, M.; Bel Haj Amor, H.; Mzoughi, Z.; Dridi, M.; Ben Said, R.; Bouraoui, A. Phytochemical analysis and evaluation of the antioxidant, anti-inflammatory, and anti-nociceptive potential of phlorotannin-rich fractions from three Mediterranean brown seaweeds. Mar. Biotechnol. 2018, 20, 60–74. [Google Scholar] [CrossRef]
- Komari, S.S.; Kumar, V.D.; Priyanka, B. Antifungal efficacy of seaweed extracts against fungal pathogen of silkworm, Bombyx mori L. Int. J. Agric. Res. 2017, 12, 123–129. [Google Scholar] [CrossRef]
- Singh, M.J.; Raadha, C.K. Studies on the antimicrobial potency of the marine algae-Gracilaria corticata var cylindrical and Hydroclathrus clathratus. Adv. Life Sci. Health 2015, 2, 50–55. [Google Scholar]
- Dayuti, S. Antibacterial activity of red algae (Gracilaria verrucosa) extract against Escherichia coli and Salmonella typhimurium. Earth Environ. Sci. 2018, 137, 1–5. [Google Scholar]
- Kolanjinathan, K.; Stella, D. Pharmacological effect of Gracilaria corticata solvent extracts against human pathogenic bacteria and fungi. Int. J. Pharmacol. Biol. Arch. 2011, 2, 1722–1728. [Google Scholar]
- Perry, N.B.; Blunt, J.W.; Munro, M.H. Cytotoxic and antifungal 1, 4-naphthoquinone and related compounds from a New Zealand brown algae. Landsburgia quercifolia. J. Nat. Prod. 1991, 54, 978–985. [Google Scholar] [CrossRef]
- Elena, M.; Francisco, Y.; Erickson, K.L. Mailiohydrin, a cytotoxic chamigrene dibromohydrin from a Phillippine Laurencia species. J. Nat. Prod. 2003, 64, 790–791. [Google Scholar]
- Smit, A.J. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q.; Hara, Y.; Shimamura, T. Inhibition of penicillinase by epigallocatechin gallate resulting in restoration of antibacterial activity of penicillin against penicillinase-producing Staphylococcus aureus. Antim. Agents Chemo. 2002, 46, 2266–2268. [Google Scholar] [CrossRef] [Green Version]
- Shiota, S.; Shimizu, M.; Mizusima, T.; Ito, H.; Hatano, T.; Yoshida, T.; Tsuchiya, T. Restoration of the effectiveness of beta-lactams on methicillin-resistant Staphylococcus aureus by tellimagrandin I from rose red. FEMS Microbiol. Lett. 2000, 185, 135–138. [Google Scholar]
- Ekambaram, P.S.; Perumal, S.S.; Balakrishnan, A.; Marappan, N.; Gajendran, S.S.; Viswanathan, V. Antibacterial synergy between rosmarinic acid and antibiotics against methicillin-resistant Staphylococcus aureus. J. Int. Ethnopharm. 2016, 5, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Arima, H.; Ashida, H.; Danno, G. Rutinen hanced antibacterial activities of flavonoids against Bacillus cereus and Salmonella enteritidis. Biosci. Biotechnol. Biochem. 2002, 66, 1009–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, V.M.; Carraro, E.; Auler, M.E.; Khalil, N.M. Quercetin and rutin as potential agents antifungal against Cryptococcus spp. Braz. J. Biol. 2016, 76, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Skocibusic, M.; Bezic, N.; Dunkic, V. Food Chemistry. Phytochemical composition and antimicrobial activities of the essential oil from Satureja subspicata Vis. Grow. Croatia 2006, 96, 20–28. [Google Scholar]
- Borghi, E.; Morace, G.; Borgo, F.; Rajendran, R.; Sherry, L.; Nile, C.; Ramage, G. New strategic insights into managing fungal biofilms. Front. Microbiol. 2015, 6, 1077. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.G. Novel antifungal mechanism of resveratrol: Apoptosis inducer in Candida albicans. Curr. Microbiol. 2015, 70, 383–389. [Google Scholar] [CrossRef]
- Hwang, I.; Lee, J.; Jin, H.G.; Woo, E.R.; Lee, D.G. Amentoflavone stimulates mitochondrial dysfunction and induces apoptotic cell death in Candida albicans. Mycopathologia 2012, 173, 207–218. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Lee, D.G. The antimicrobial peptide arenicin-1 promotes generation of reactive oxygen species and induction of apoptosis. Biochim. Biophys. Acta 2011, 1810, 1246–1250. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Salvador, V.A.G. Evaluation of Apoptosis and Necrosis in Saccharomyces cerevisiae during Wine Fermentations. Ph.D. Thesis, Universidade Técnica de Lisboa, Lisbon, Portugal, 2009. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retension Time | Name of the Compound | Area % | Bioactivity | Reference |
|---|---|---|---|---|
| 3.447 | Boron, trihydro (N-methylmethanamine-, (T-4)- | 0.36 | ||
| 4.912 | 2 (3H)-Furanone, dihydro-4-hydroxy- | 0.45 | Antibacterial | [35] |
| 6.114 | 1-Hepten-3-ol | 0.21 | - | |
| 6.646 | D-Ribonic acid, 2,3-O-(ethoxymethylene)- | 0.37 | - | |
| 8.031 | 2-Octyldodecan-1-ol | 0.19 | - | |
| 8.723 | Docosane | 0.88 | Antifangal | [36] |
| 11.824 | 2-Decyne | 0.46 | - | |
| 12.900 | 5-Thiazoleethanol, 4-methyl- | 2.12 | Antifungal, anti-inflammatory, anti-allergic | [37] |
| 13.381 | Carvacrol | 0.25 | Anti-inflammatory, antioxidant, antitumor, antibacterial | [38] |
| 17.380 | 2-(1,4,4-Trimethyl-cyclohex-2-enyl)-ethanol | 1.21 | ||
| 17.541 | Decamethylpentasiloxane | 0.83 | Antibacterial, antifungal | [39] |
| 18.044 | 2-cyclohexene-1-one | 0.62 | Antibacterial | [40] |
| 18.170 | 10-Methyl-9-oxabicyc lo[6.4.0]dodecan-1(8)-ene | 0.43 | - | |
| 18.342 | 2-(4H)-Benzofuranone, 5,6,7,7a-tetrahydro-4,4,7a-trimethyl- | 0.76 | Antimicrobial preservative, antifungal, antibacterial | [41] |
| 19.051 | 2-Propenoic acid, 3-(1-aziridinyl)-,methyl ester | 0.85 | ||
| 19515 | Diethyl Phthalate | 1.06 | Antioxidant | [42] |
| 20.213 | Methanone, diphenyl- | 0.98 | Antibacterial, antifungal | [43] |
| 20.831 | Cyclododecasiloxane, tetracosamethyl- | 1.37 | Antimicobial, antirheumatic antispasmodic | [44] |
| 21.409 | Heptadecane | 7.71 | Anticancer, anti-inflammatory | [38] |
| 21.718 | n-Dodecanal | 0.66 | Antibacterial | [41] |
| 24.230 | Cyclododecasiloxane, Tetracosamethyl- | 0.39 | Antimicobial, antirheumatic antispasmodic | [44] |
| 24.662 | Caffeine | 0.97 | Antibacterial, antifungal | [45] |
| 24.401 | Bicyclo[3.1.1] heptane, 2,6,6-trimethyl Pinane | 3.17 | Antifungal | [46] |
| 24.521 | 11-Dodecen-2-one | 1.13 | ||
| 24.853 | E-10-Methyl-11-tetradecen-1-ol acetate | 0.76 | ||
| 24.991 | Isobutyl phthalate | 2.25 | Antibacterial, antifungal | [42] |
| 25.168 | Neophytadine | 1.66 | Antimicrobial, anti-inflammatory | [47] |
| 25.591 | 2-Propenoic acid, 2-methyl- | 0.43 | Antibacterial, antifungal | [48] |
| 25.889 | Palmitic acid methyl ester | 6.66 | Antibacterial, antifungal | [41] |
| 26.444 | Palmitic acid | 17.11 | Antibacterial, antifungal | [49] |
| 26.753 | Oleic acid | 0.24 | Antibacterial, antifungal | [49] |
| 26.942 | 1-Decanol, 2-hexyl- | 0.46 | Antimicrobial | [50] |
| 28.178 | Linolelaidic acid, methyl ester | 0.66 | Antibacterial, antifungal | [41] |
| 28.246 | Oleic acid methyl ester | 1.18 | Antibacterial, antifungal | [51] |
| 28.309 | Oleic acid methyl ester | 0.94 | Antibacterial, antifungal | [41] |
| 28.384 | Phytol | 2.34 | Antibacterial, antifungal | [52] |
| 28.538 | Stearic acid methyl ester | 1.01 | Antibacterial, antifungal | [41] |
| 28.693 | 9-Hexadecenoic acid | 1.73 | Antibacterial, inflammator | [52] |
| 28.933 | 9-Hexadecenoic acid | 0.32 | Antibacterial, inflammator | [52] |
| 29.608 | E-11-Tetradecenoic acid | 0.25 | Antibacterial, antifungal | [52] |
| 29.860 | Hexadecanedioic acid | 0.93 | Antimicrobial, anti-inflammatory | [52] |
| 30.175 | 9-Borabicyclo[3.3.1]nonane, dimethylamino)propyl] | 0.27 | - | |
| 0.306 | (-)-18-noramborx | 0.45 | - | |
| 30.592 | Trisiloxane, 1,1,1,3,5,5,5-heptamethyl | 0.53 | Antifungal | [53] |
| 30.896 | Hexamethylcyclotrisiloxane | 0.28 | Antimicrobial | [54] |
| 32.034 | 1,3-Bis(trimethylsilyl)benzene | 0.79 | - | |
| 32.206 | p-Bis(trimethylsilyl)benzene | 0.47 | - | |
| 32.858 | Phthalic acid,bis(2-ethylhexyl) ester | 0.75 | Antibacterial, antifungal | [35] |
| 38.340 | N-Methyl-1-adamantane acetamide | 2.52 | Antimicrobial | [55] |
| 45.258 | Cholest-5-en-3-ol (3.beta.)- | 27.64 | Antibacterial | [56,57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pourakbar, L.; Moghaddam, S.S.; Enshasy, H.A.E.; Sayyed, R.Z. Antifungal Activity of the Extract of a Macroalgae, Gracilariopsis persica, against Four Plant Pathogenic Fungi. Plants 2021, 10, 1781. https://doi.org/10.3390/plants10091781
Pourakbar L, Moghaddam SS, Enshasy HAE, Sayyed RZ. Antifungal Activity of the Extract of a Macroalgae, Gracilariopsis persica, against Four Plant Pathogenic Fungi. Plants. 2021; 10(9):1781. https://doi.org/10.3390/plants10091781
Chicago/Turabian StylePourakbar, Latifeh, Sina Siavash Moghaddam, Hesham Ali El Enshasy, and R. Z. Sayyed. 2021. "Antifungal Activity of the Extract of a Macroalgae, Gracilariopsis persica, against Four Plant Pathogenic Fungi" Plants 10, no. 9: 1781. https://doi.org/10.3390/plants10091781