
The Effects of Ozone on Visual Attraction Traits of Erodium paularense (Geraniaceae) Flowers: Modelled Perception by Insect Pollinators

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Ozone Exposure and Growing Conditions
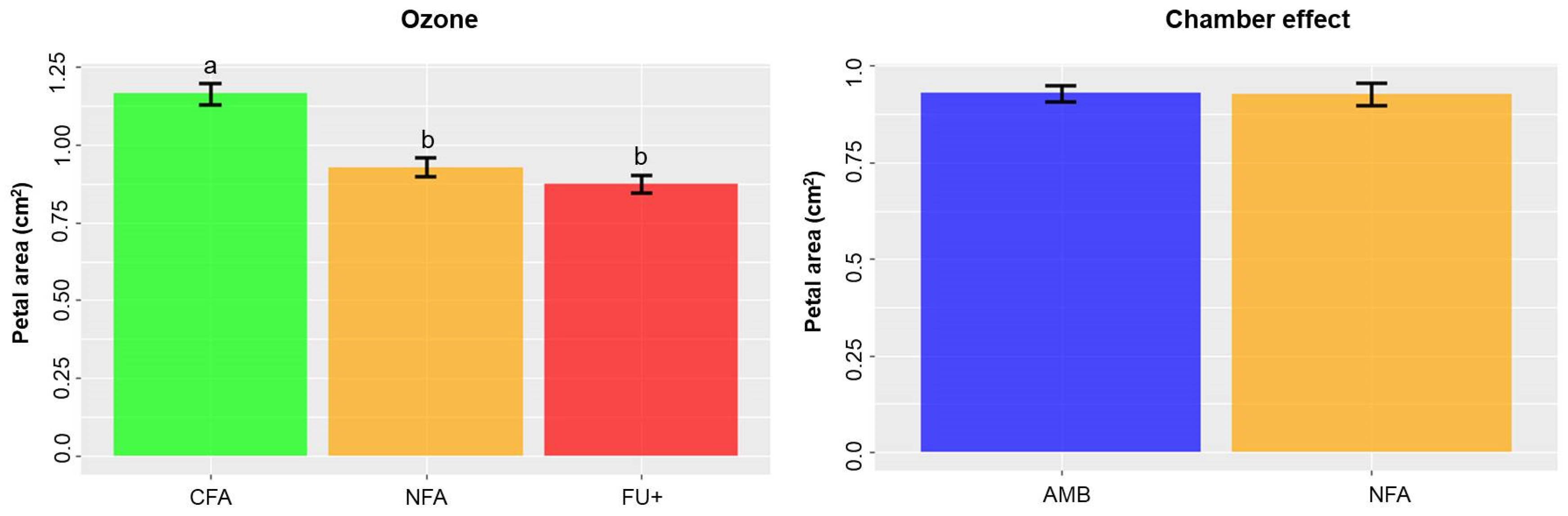
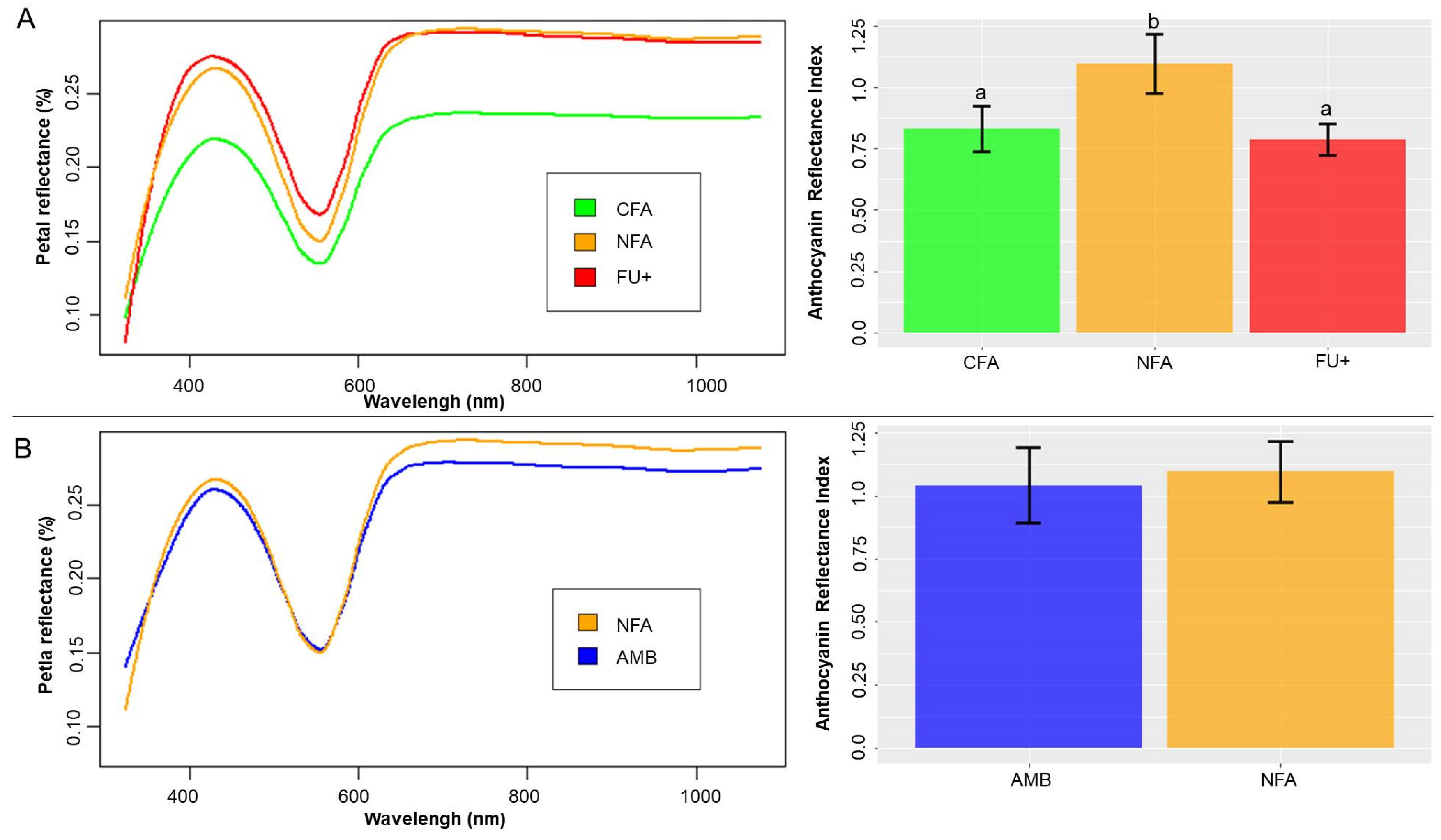
2.2. Ozone and Chamber Effects on Petal Area, Reflectance and ARI
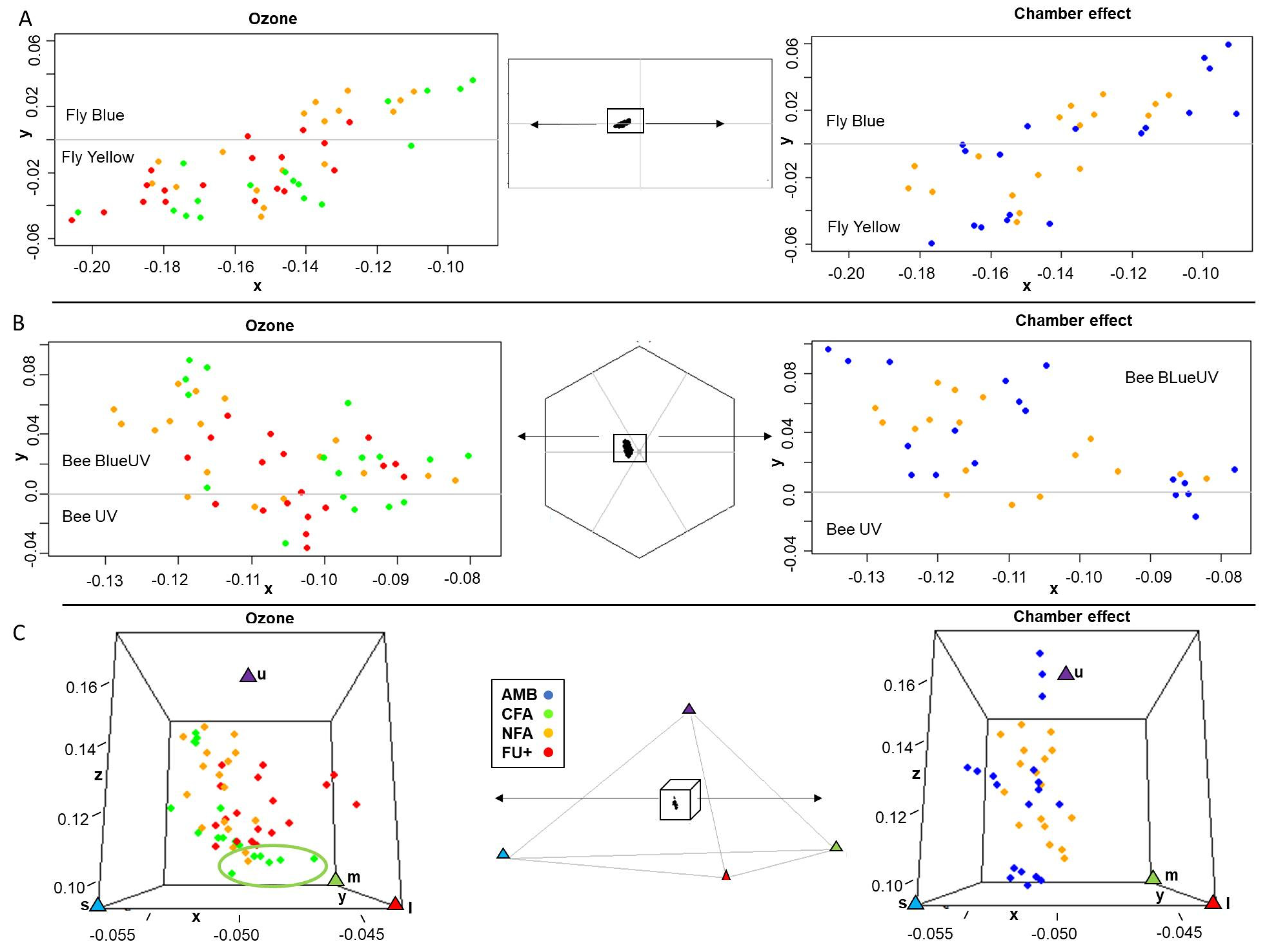
2.3. Ozone and Chamber Effects on Pollinator Color Perception
3. Discussion
4. Materials and Methods
4.1. OTC Experiment
4.2. Plant Material
4.3. Spectral Reflectance Measurements, Anthocyanin Index and Petal Area
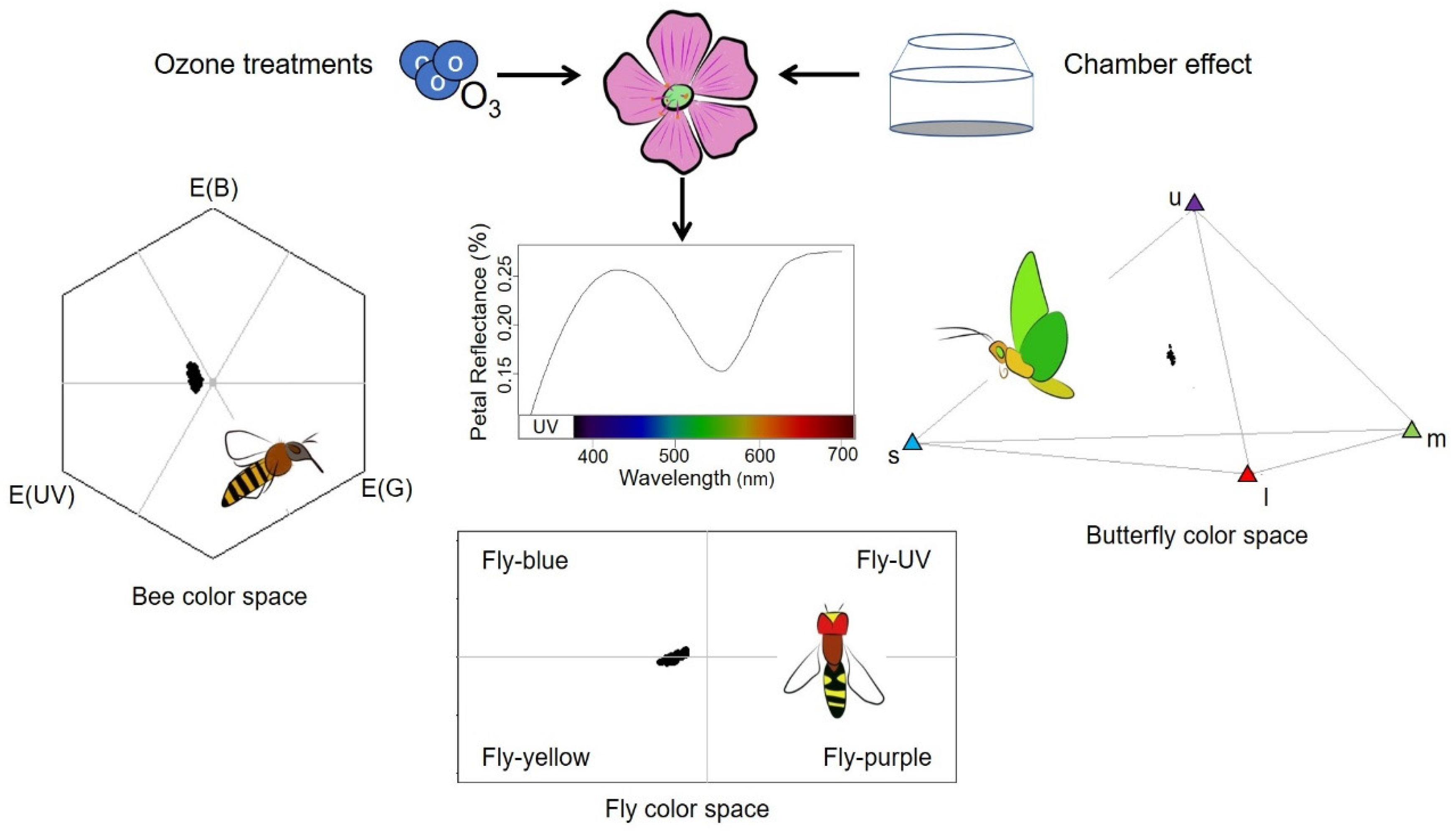
4.4. Pollinator Color Perception
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ainsworth, E.A.; Yendrek, C.R.; Sitch, S.; Collins, W.J.; Emberson, L.D. The Effects of Tropospheric Ozone on Net Primary Productivity and Implications for Climate Change. Annu. Rev. Plant Biol. 2012, 63, 637–661. [Google Scholar] [CrossRef] [Green Version]
- Chappelka, A.H.; Samuelson, L.J. Ambient ozone effects on forest trees of the eastern United States: A review. New Phytol. 1998, 139, 91–108. [Google Scholar] [CrossRef]
- Leisner, C.P.; Ainsworth, E.A. Quantifying the effects of ozone on plant reproductive growth and development. Glob. Chang. Biol. 2011, 18, 606–616. [Google Scholar] [CrossRef]
- Mills, G.; Sharps, K.; Simpson, D.; Pleijel, H.; Frei, M.; Burkey, K.; Emberson, L.; Uddling, J.; Broberg, M.; Feng, Z.; et al. Closing the global ozone yield gap: Quantification and cobenefits for multistress tolerance. Glob. Chang. Biol. 2018, 24, 4869–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elvira, S.; Fernández, I.G.; Alonso, R.; Sanz, J.; Bermejo-Bermejo, V. Ozone levels in the Spanish Sierra de Guadarrama mountain range are above the thresholds for plant protection: Analysis at 2262, 1850, and 995 m a.s.l. Environ. Monit. Assess. 2016, 188, 593. [Google Scholar] [CrossRef]
- Monks, P.S.; Archibald, A.T.; Colette, A.; Cooper, O.; Coyle, M.; Derwent, R.; Fowler, D.; Granier, C.; Law, K.S.; Mills, G.E.; et al. Tropospheric ozone and its precursors from the urban to the global scale from air quality to short-lived climate forcer. Atmos. Chem. Phys. Discuss. 2015, 15, 8889–8973. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, E.; De Marco, A.; Beddows, D.; Harrison, R.M.; Manning, W.J. Ozone levels in European and USA cities are increasing more than at rural sites, while peak values are decreasing. Environ. Pollut. 2014, 192, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Querol, X.; Alastuey, A.; Gangoiti, G.; Perez, N.; Lee, H.K.; Eun, H.R.; Park, Y.; Mantilla, E.; Escudero, M.; Titos, G.; et al. Phenomenology of summer ozone episodes over the Madrid Metropolitan Area, central Spain. Atmos. Chem. Phys. Discuss. 2018, 18, 6511–6533. [Google Scholar] [CrossRef] [Green Version]
- Querol, X.; Alastuey, A.; Pandolfi, M.; Reche, C.; Pérez, N.; Minguillón, M.; Moreno, T.; Viana, M.; Escudero, M.; Orio, A.; et al. 2001–2012 trends on air quality in Spain. Sci. Total Environ. 2014, 490, 957–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agathokleous, E.; Feng, Z.; Oksanen, E.; Sicard, P.; Wang, Q.; Saitanis, C.J.; Araminiene, V.; Blande, J.D.; Hayes, F.; Calatayud, V.; et al. Ozone affects plant, insect, and soil microbial communities: A threat to terrestrial ecosystems and biodiversity. Sci. Adv. 2020, 6, eabc1176. [Google Scholar] [CrossRef]
- Fuhrer, J.; Martin, M.V.; Mills, G.; Heald, C.L.; Harmens, H.; Hayes, F.; Sharps, K.; Bender, J.; Ashmore, M.R. Current and future ozone risks to global terrestrial biodiversity and ecosystem processes. Ecol. Evol. 2016, 6, 8785–8799. [Google Scholar] [CrossRef] [Green Version]
- Cristofanelli, P.; Bonasoni, P. Background ozone in the southern Europe and Mediterranean area: Influence of the transport processes. Environ. Pollut. 2009, 157, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Millán, M.; Salvador, R.; Mantilla, E.; Artnano, B. Meteorology and photochemical air pollution in Southern Europe: Experimental results from EC research projects. Atmos. Environ. 1996, 30, 1909–1924. [Google Scholar] [CrossRef]
- Mills, G.; Wagg, S.; Harmens, H. Ozone Pollution: Impacts on Ecosystem Services and Biodiversity; NERC/Centre for Ecology & Hydrology: Bangor, UK, 2013; p. 104. [Google Scholar]
- Di Gregorio, A.; Jansen, L.J.M. Land Cover Classification System (LCCS): Classification Concepts and User Manual; Food and Agriculture Organization: Rome, Italy, 2000. [Google Scholar]
- Hayes, F.; Harmens, H.; Mills, G.; Bender, J.; Grünhage, L. Ozone critical levels for (semi-) natural vegetation dominated by perennial grassland species. Environ. Sci. Pollut. Res. 2020, 28, 15090–15098. [Google Scholar] [CrossRef]
- Sanz, J.; Fernández, I.G.; Elvira, S.; Muntifering, R.; Alonso, R.; Bermejo-Bermejo, V. Setting ozone critical levels for annual Mediterranean pasture species: Combined analysis of open-top chamber experiments. Sci. Total Environ. 2016, 571, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Rämö, K.; Kanerva, T.; Ojanperä, K.; Manninen, S. Growth onset, senescence, and reproductive development of meadow species in mesocosms exposed to elevated O3 and CO2. Environ. Pollut. 2007, 145, 850–860. [Google Scholar] [CrossRef]
- Sanz, J.; Bermejo, V.; Muntifering, R.; González-Fernández, I.; Gimeno, B.; Elvira, S.; Alonso, R. Plant phenology, growth and nutritive quality of Briza maxima: Responses induced by enhanced ozone atmospheric levels and nitrogen enrichment. Environ. Pollut. 2011, 159, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.; Muntifering, R.; Lin, J.; Weigel, H. Growth and nutritive quality of Poa pratensis as influenced by ozone and competition. Environ. Pollut. 2006, 142, 109–115. [Google Scholar] [CrossRef]
- Bergweiler, C.; Manning, W. Inhibition of flowering and reproductive success in spreading dogbane (Apocynum androsaemifolium) by exposure to ambient ozone. Environ. Pollut. 1999, 105, 333–339. [Google Scholar] [CrossRef]
- Danielsson, H.; Gelang, J.; Pleijel, H. Ozone sensitivity, growth and flower development in Phleum genotypes of different geographic origin in the Nordic countries. Environ. Exp. Bot. 1999, 42, 41–49. [Google Scholar] [CrossRef]
- Drogoudi, P.; Ashmore, M.R. Does elevated ozone have differing effects in flowering and deblossomed strawberry? New Phytol. 2000, 147, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Franzaring, J.; Tonneijck, A.; Kooijman, A.; Dueck, T. Growth responses to ozone in plant species from wetlands. Environ. Exp. Bot. 2000, 44, 39–48. [Google Scholar] [CrossRef]
- Hayes, F.; Williamson, J.; Mills, G. Ozone pollution affects flower numbers and timing in a simulated BAP priority calcareous grassland community. Environ. Pollut. 2012, 163, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.; Black, V.; Black, C.; Roberts, J. Direct Effects of Ozone on the Reproductive Development of Brassica Species. J. Plant Physiol. 1996, 148, 172–178. [Google Scholar] [CrossRef]
- Taylor, M.D.; Sinn, J.P.; Davis, D.D.; Pell, E.J. The impact of ozone on a salt marsh cordgrass (Spartina alterniflora). Environ. Pollut. 2002, 120, 701–705. [Google Scholar] [CrossRef]
- Duque, L.; Poelman, E.H.; Steffan-Dewenter, I. Effects of ozone stress on flowering phenology, plant-pollinator interactions and plant reproductive success. Environ. Pollut. 2021, 272, 115953. [Google Scholar] [CrossRef]
- Kohut, R.J.; Laurence, J.A.; Colavito, L.J. The influence of ozone exposure dynamics on the growth and yield of kidney bean. Environ. Pollut. 1988, 53, 79–88. [Google Scholar] [CrossRef]
- Retzlaff, W.A.; Williams, L.E.; Dejong, T.M. Growth and Yield Response of Commercial Bearing-age’ Casselman’ Plum Trees to Various Ozone Partial Pressures. ASA 1997, 26, 858–865. [Google Scholar] [CrossRef]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination Syndromes and Floral Specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Van Der Pijl, L. Ecological aspects of flower evolution. I. Phyletic evolution. Evolution 1960, 14, 403–416. [Google Scholar] [CrossRef]
- Simpson, B.B.; Neff, J.L. Floral Rewards: Alternatives to Pollen and Nectar. Ann. Mo. Bot. Gard. 1981, 68, 301. [Google Scholar] [CrossRef]
- Ribeiro, H.; Duque, L.; Sousa, R.; Abreu, I. Ozone effects on soluble protein content of Acer negundo, Quercus robur and Platanus spp. pollen. Aerobiologia 2013, 29, 443–447. [Google Scholar] [CrossRef]
- Stabler, D. Impacts of Ozone Pollution on Nectar and Pollen Quality and Their Significance for Pollinators. Doctoral Dissertation, University of Newcastle, Newcastle upon Tyne, UK, 2016. [Google Scholar]
- Saunier, A.; Blande, J.D. The effect of elevated ozone on floral chemistry of Brassicaceae species. Environ. Pollut. 2019, 255, 113257. [Google Scholar] [CrossRef]
- Blande, J.D.; Holopainen, J.K.; Niinemets, Ü. Plant volatiles in polluted atmospheres: Stress responses and signal degradation. Plant Cell Environ. 2014, 37, 1892–1904. [Google Scholar] [CrossRef] [Green Version]
- Farré-Armengol, G.; Penuelas, J.; Li, T.; Yli-Pirilä, P.; Filella, I.; Llusia, J.; Blande, J.D. Ozone degrades floral scent and reduces pollinator attraction to flowers. New Phytol. 2016, 209, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFrederick, Q.S.; Fuentes, J.D.; Roulston, T.; Kathilankal, J.C.; Lerdau, M. Effects of air pollution on biogenic volatiles and ecological interactions. Oecologia 2009, 160, 411–420. [Google Scholar] [CrossRef]
- McFrederick, Q.S.; Kathilankal, J.C.; Fuentes, J.D. Air pollution modifies floral scent trails. Atmos. Environ. 2008, 42, 2336–2348. [Google Scholar] [CrossRef]
- Giurfa, M.; Lehrer, M. Honeybee vision and floral displays: From detection to close-up recognition. In Cognitive Ecology of Pollination: Animal Behaviour and Floral Evolution; Chittka, L., Thomson, J., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 61–82. [Google Scholar] [CrossRef]
- Roshchina, V.V. The Autofluorescence Response of Flower Cells from Saintpaulia Ionantha as the Biosensor Reaction to Ozone. Comput. Biol. Bioinform. 2017, 4, 60–66. [Google Scholar] [CrossRef]
- Arnold, S.E.J.; Savolainen, V.; Chittka, L. Flower colours along an alpine altitude gradient, seen through the eyes of fly and bee pollinators. Arthropod-Plant Interact. 2009, 3, 27–43. [Google Scholar] [CrossRef]
- Ohashi, K.; Makino, T.T.; Arikawa, K. Floral colour change in the eyes of pollinators: Testing possible constraints and correlated evolution. Funct. Ecol. 2015, 29, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- González-Benito, E.; Martin, C.; Iriondo, J. Autecology and conservation of Erodium paularense Fdez. Glez. & Izco. Biol. Conserv. 1995, 72, 55–60. [Google Scholar] [CrossRef]
- Martín, C.; González-Benito, M.E.; Iriondo, J.M. The use of genetic markers in the identification and characterization of three recently discovered populations of a threatened plant species. Mol. Ecol. 1999, 8, S31–S40. [Google Scholar] [CrossRef]
- Moreno, J.C. Lista Roja 2008 de la Flora Vascular Española; Dirección General de Medio Natural y PolíticaForestal (Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la Conservaciónde Plantas): Madrid, Spain, 2008. (In Spanish) [Google Scholar]
- Albert, M.J.; Escudero, A.; Iriondo, J. Female Reproductive Success of Narrow Endemic Erodium paularense in Contrasting Microhabitats. Ecology 2001, 82, 1734–1747. [Google Scholar] [CrossRef]
- Albert, M.J. Biología de la Conservación de Erodium Paularense Fern. Gonz. & Izco (Geraniaceae). Doctoral Thesis, Universidad Politécnica of Madrid, Madrid, Spain, 2003. (In Spanish). [Google Scholar]
- Foot, J.P.; Caporn, S.J.M.; Lee, J.A.; Ashenden, T.W. The effect of long-term ozone fumigation on the growth, physiology and frost sensitivity of Calluna vulgaris. New Phytol. 1996, 133, 503–511. [Google Scholar] [CrossRef]
- Ansari, N.; Agrawal, M.; Agrawal, S.B. An assessment of growth, floral morphology, and metabolites of a medicinal plant Sida cordifolia L. under the influence of elevated ozone. Environ. Sci. Pollut. Res. 2020, 28, 832–845. [Google Scholar] [CrossRef]
- Gómez, J.M.; Perfectti, F.; Armas, C.; Narbona, E.; González-Megías, A.; Navarro, L.; DeSoto, L.; Torices, R. Within-individual phenotypic plasticity in flowers fosters pollination niche shift. Nat. Commun. 2020, 11, 4019. [Google Scholar] [CrossRef]
- de Jong, T.J.; Waser, N.M.; Klinkhamer, P.G. Geitonogamy: The neglected side of selfing. Trends Ecol. Evol. 1993, 8, 321–325. [Google Scholar] [CrossRef]
- Rudall, P.J. Colourful cones: How did flower colour first evolve? J. Exp. Bot. 2020, 71, 759–767. [Google Scholar] [CrossRef]
- Martin, C.; Gerats, T. Control of Pigment Biosynthesis Genes during Petal Development. Plant Cell 1993, 5, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Kay, Q.; Daoud, H.; Stirton, C. Pigment distribution, light reflection and cell structure in petals. Bot. J. Linn. Soc. 1981, 83, 57–83. [Google Scholar] [CrossRef]
- Boyer, M.; Miller, J.; Belanger, M.; Hare, E.; Wu, J. Senescence and spectral reflectance in leaves of northern pin oak (Quercus palustris Muenchh). Remote Sens. Environ. 1988, 25, 71–87. [Google Scholar] [CrossRef]
- Menzel, R.; Shmida, A. The Ecology of flower colours and the natural colour vision of insect pollinators: The Israeli flora as a study case. Biol. Rev. 1993, 68, 81–120. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Chivkunova, O.B. Light-stress-induced pigment changes and evidence for anthocyanin photoprotection in apples. J. Photochem. Photobiol. B Biol. 2000, 55, 155–163. [Google Scholar] [CrossRef]
- Arnold, S.E.J.; Faruq, S.; Savolainen, V.; McOwan, P.W.; Chittka, L. FReD: The Floral Reflectance Database—A Web Portal for Analyses of Flower Colour. PLoS ONE 2010, 5, e14287. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental Significance of Anthocyanins in Plant Stress Responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- de Rezende, F.M.; Furlan, C.M. Anthocyanins and tannins in ozone-fumigated guava trees. Chemosphere 2009, 76, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Gravano, E.; Bussotti, F.; Strasser, R.J.; Schaub, M.; Novak, K.; Skelly, J.; Tani, C. Ozone symptoms in leaves of woody plants in open-top chambers: Ultrastructural and physiological characteristics. Physiol. Plant. 2004, 121, 620–633. [Google Scholar] [CrossRef]
- Koukol, J.; Dugger, W.M. Anthocyanin formation as a response to ozone and smog treatment in Rumex crispus L. Plant Physiol. 1967, 42, 1023–1024. [Google Scholar] [CrossRef] [Green Version]
- Rozpądek, P.; Nosek, M.; Ślesak, I.; Kunicki, E.; Dziurka, M.; Miszalski, Z. Ozone fumigation increases the abundance of nutrients in Brassica vegetables: Broccoli (Brassica oleracea var. italica) and Chinese cabbage (Brassica pekinensis). Eur. Food Res. Technol. 2014, 240, 459–462. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Murphy, M.R.C.; Cardoso, A.A.; Jordan, G.J.; Brodribb, T.J. Similar geometric rules govern the distribution of veins and stomata in petals, sepals and leaves. New Phytol. 2018, 219, 1224–1234. [Google Scholar] [CrossRef] [Green Version]
- Costa, V.B.S.; Pimentel, R.M.M.; Chagas, M.G.S.; Alves, G.D.; Castro, C.C. Petal micromorphology and its relationship to pollination. Plant Biol. 2017, 19, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, P.; Günthardt-Goerg, M.S.; Menard, T.; Baumgarten, M.; Matyssek, R.; Schaub, M. Macro- and microscopic leaf injury triggered by ozone stress in beech foliage (Fagus sylvatica L.). Ann. For. Sci. 2019, 76, 71. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology; Springer: Berlin/Heidelberg, Germany, 2003; p. 514. ISBN 978-3-540-43516-7. [Google Scholar]
- Hegland, S.J.; Nielsen, A.; Lázaro, A.; Bjerknes, A.-L.; Totland, Ø. How does climate warming affect plant-pollinator interactions? Ecol. Lett. 2009, 12, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Scaven, V.L.; Rafferty, N.E. Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions. Curr. Zool. 2013, 59, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dela, G.; Or, E.; Ovadia, R.; Nissim-Levi, A.; Weiss, D.; Oren-Shamir, M. Changes in anthocyanin concentration and composition in ‘Jaguar’ rose flowers due to transient high-temperature conditions. Plant Sci. 2003, 164, 333–340. [Google Scholar] [CrossRef]
- Nozaki, K.; Takamura, T.; Fukai, S. Effects of high temperature on flower colour and anthocyanin content in pink flower genotypes of greenhouse chrysanthemum (Chrysanthemum morifoliumRamat). J. Hortic. Sci. Biotechnol. 2006, 81, 728–734. [Google Scholar] [CrossRef]
- Shaked-Sachray, L.; Weiss, D.; Reuveni, M.; Nissim-Levi, A.; Oren-Shamir, M. Increased anthocyanin accumulation in aster flowers at elevated temperatures due to magnesium treatment. Physiol. Plant. 2002, 114, 559–565. [Google Scholar] [CrossRef]
- Heck, W.W.; Taylor, O.C.; Adams, R.; Bingham, G.; Miller, J.; Preston, E.; Weinstein, L. Assessment of Crop Loss from Ozone. J. Air Pollut. Control Assoc. 1982, 32, 353–361. [Google Scholar] [CrossRef]
- Team, R.C. R Core Team: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 June 2021).
- Maia, R.; Gruson, H.; Endler, J.A.; White, T.E. PAVO 2: New tools for the spectral and spatial analysis of colour in R. Methods Ecol. Evol. 2019, 10, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Merzlyak, M.N.; Solovchenko, A.E.; Gitelson, A.A. Reflectance spectral features and non-destructive estimation of chlorophyll, carotenoid and anthocyanin content in apple fruit. Postharvest Biol. Technol. 2003, 27, 197–211. [Google Scholar] [CrossRef]
- Bourne, R. ImageJ. In Fundamentals of Digital Imaging in Medicine; Springer: London, UK, 2009; Volume 9, pp. 185–188. [Google Scholar] [CrossRef]
- Koshitaka, H.; Kinoshita, M.; Vorobyev, M.; Arikawa, K. Tetrachromacy in a butterfly that has eight varieties of spectral receptors. Proc. R. Soc. B Boil. Sci. 2008, 275, 947–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chittka, L. The colour hexagon: A chromaticity diagram based on photoreceptor excitations as a generalized representation of colour opponency. J. Comp. Physiol. A 1992, 170, 533–543. [Google Scholar] [CrossRef]
- Schad, D.J.; Vasishth, S.; Hohenstein, S.; Kliegl, R. How to capitalize on a priori contrasts in linear (mixed) models: A tutorial. J. Mem. Lang. 2020, 110, 104038. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 4 October 2020).
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Simpson, G.; Solymos, P.; Stevenes, M.H.H.; Wagner, H. Vegan: 2012 Community Ecology Package. R Package Version 2.0-2. Available online: http://CRAN.R-project.org/package=vegan (accessed on 1 June 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Ozone Exposure and N | AOT40 | 7–15 h. O3 Mean | Daily O3 Mean | ||
| Treatment | N LM or LMM | N PERM. | (nL L−1 h) | (nL L−1) | (nL L−1) |
| AMB | 17 | 18 | 2922.7 | 41.3 | 36 |
| CFA | 17 | 17 | 23.7 | 21.7 | 17.3 |
| NFA | 15 | 17 | 2333.2 | 40.1 | 35.2 |
| FU+ | 17 | 18 | 12121.5 | 64.6 | 48.8 |
| (B) Growth Conditions | |||||
| Mean Tª (°C) | Max Tª (°C) | RH (%) | VPD (kPa) | PAR µmol (m−2 s−1) | |
| Ambient | 15.8 | 28.6 | 52.5 | 0.78 | 841 |
| OTC | 19.1 | 33.6 | 56.5 | 1.06 | 698.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prieto-Benítez, S.; Ruiz-Checa, R.; Bermejo-Bermejo, V.; Gonzalez-Fernandez, I. The Effects of Ozone on Visual Attraction Traits of Erodium paularense (Geraniaceae) Flowers: Modelled Perception by Insect Pollinators. Plants 2021, 10, 2750. https://doi.org/10.3390/plants10122750
Prieto-Benítez S, Ruiz-Checa R, Bermejo-Bermejo V, Gonzalez-Fernandez I. The Effects of Ozone on Visual Attraction Traits of Erodium paularense (Geraniaceae) Flowers: Modelled Perception by Insect Pollinators. Plants. 2021; 10(12):2750. https://doi.org/10.3390/plants10122750
Chicago/Turabian StylePrieto-Benítez, Samuel, Raquel Ruiz-Checa, Victoria Bermejo-Bermejo, and Ignacio Gonzalez-Fernandez. 2021. "The Effects of Ozone on Visual Attraction Traits of Erodium paularense (Geraniaceae) Flowers: Modelled Perception by Insect Pollinators" Plants 10, no. 12: 2750. https://doi.org/10.3390/plants10122750