
Effect of Selenium Application and Growth Stage at Harvest on Hydrophilic and Lipophilic Antioxidants in Lamb’s Lettuce (Valerianella locusta L. Laterr.)

Abstract
:1. Introduction
2. Results
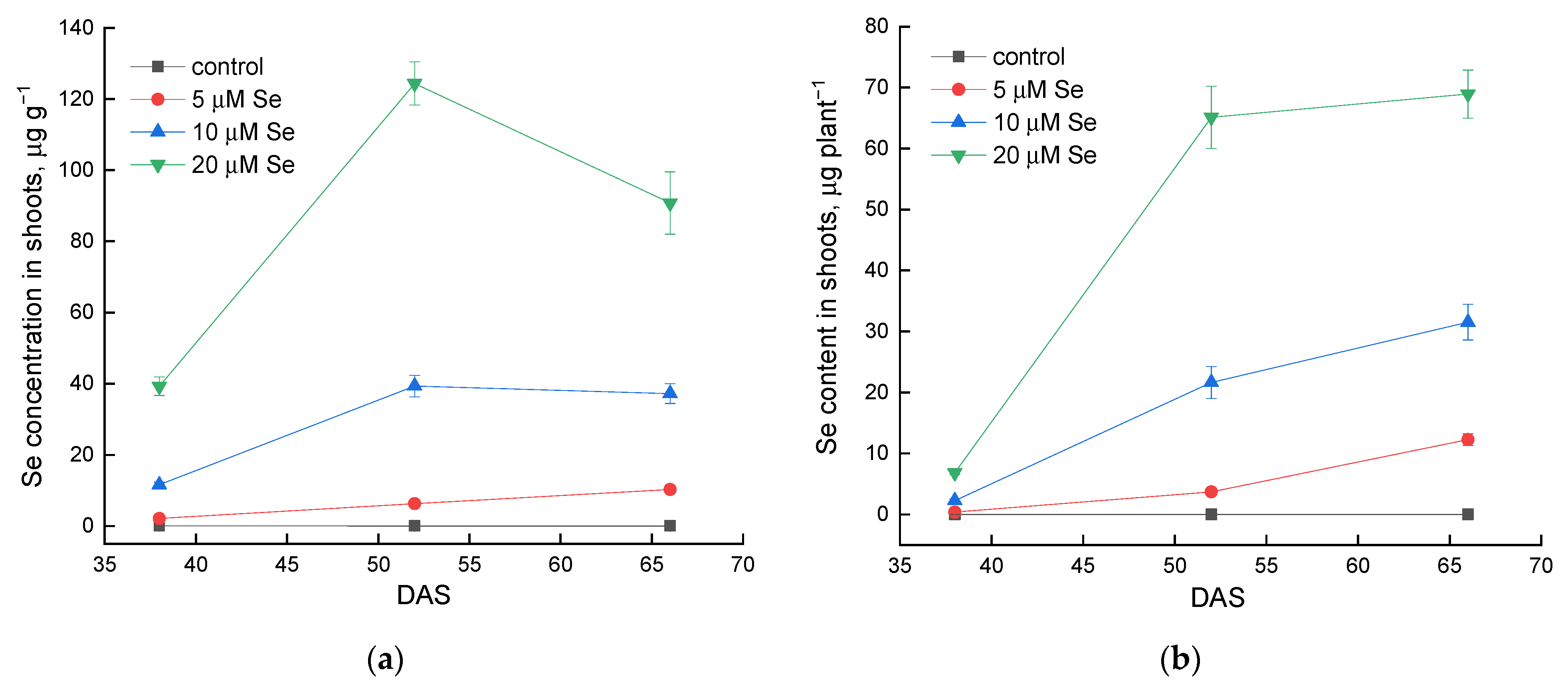
2.1. Effect of Se Application and Growth Stage at Harvest on Plant Yield and Accumulation of Se
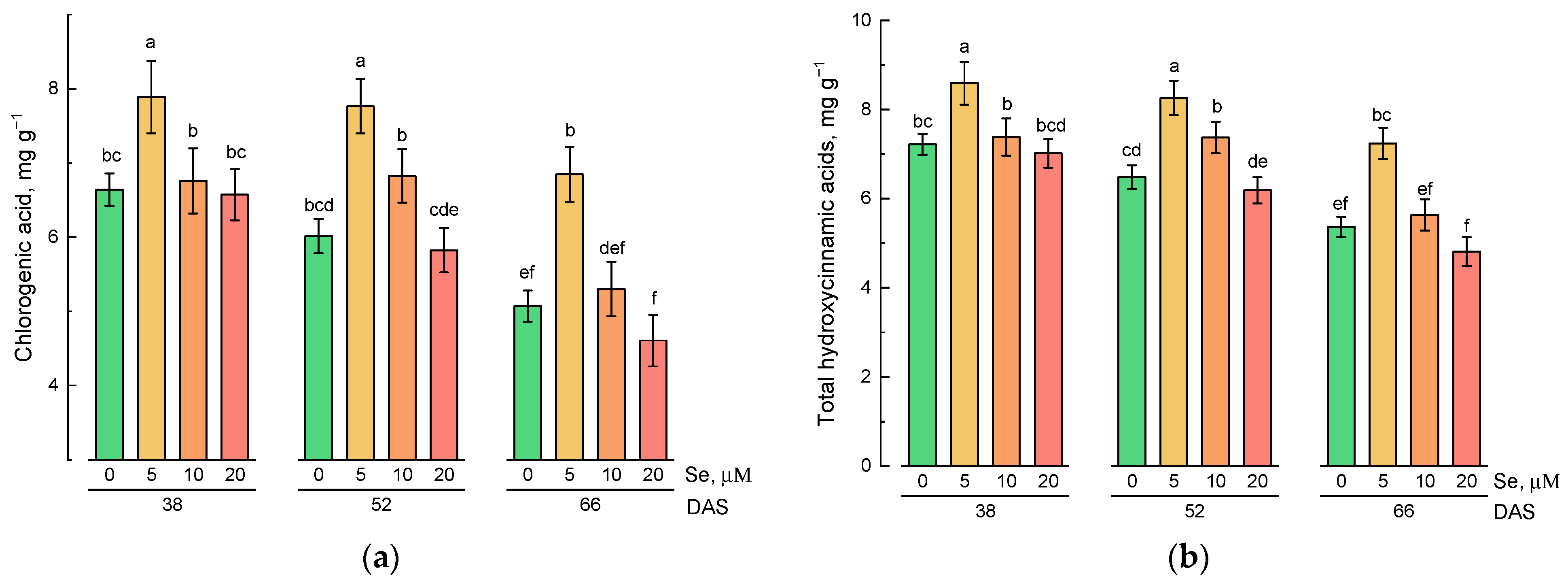
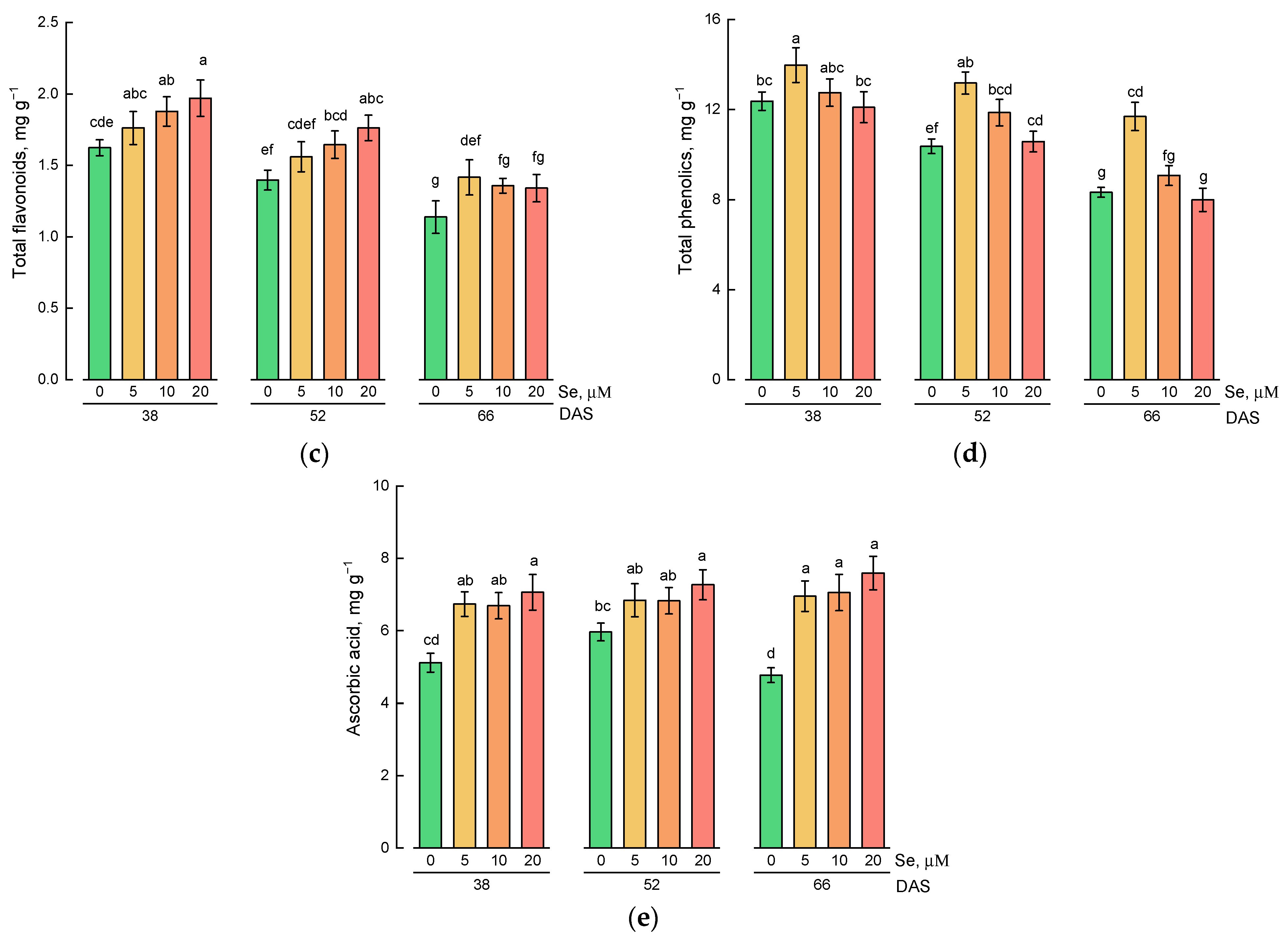
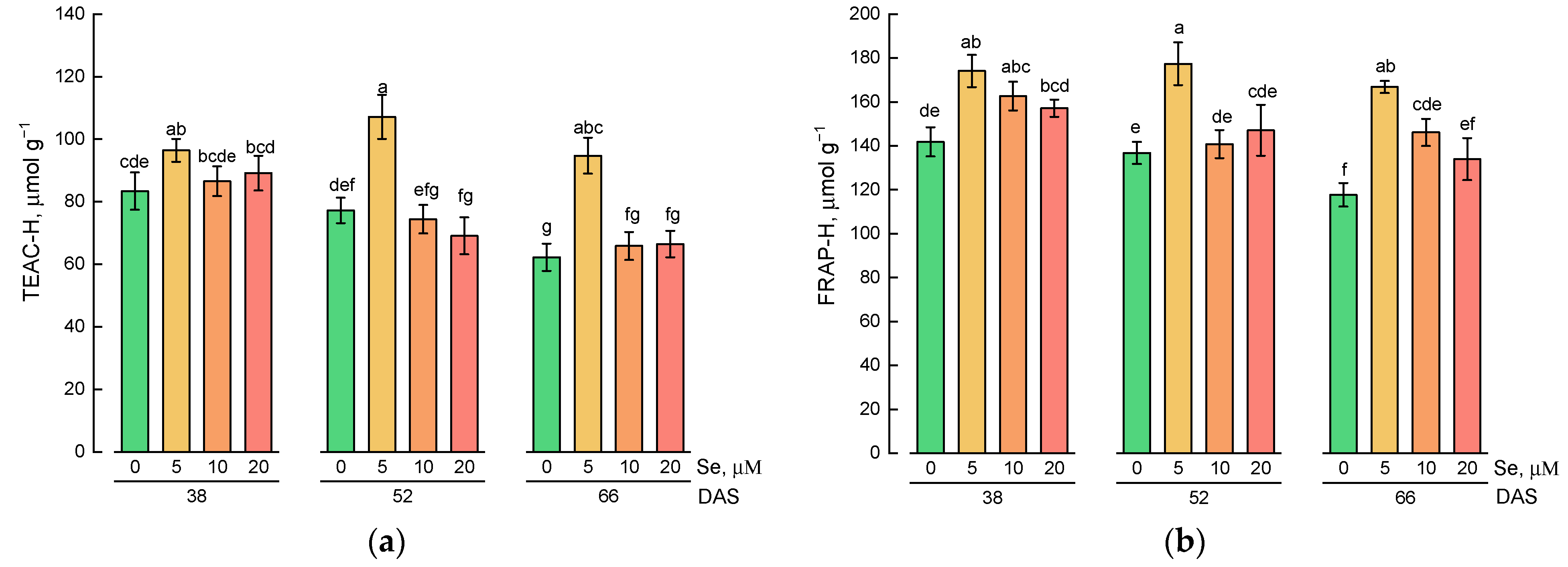
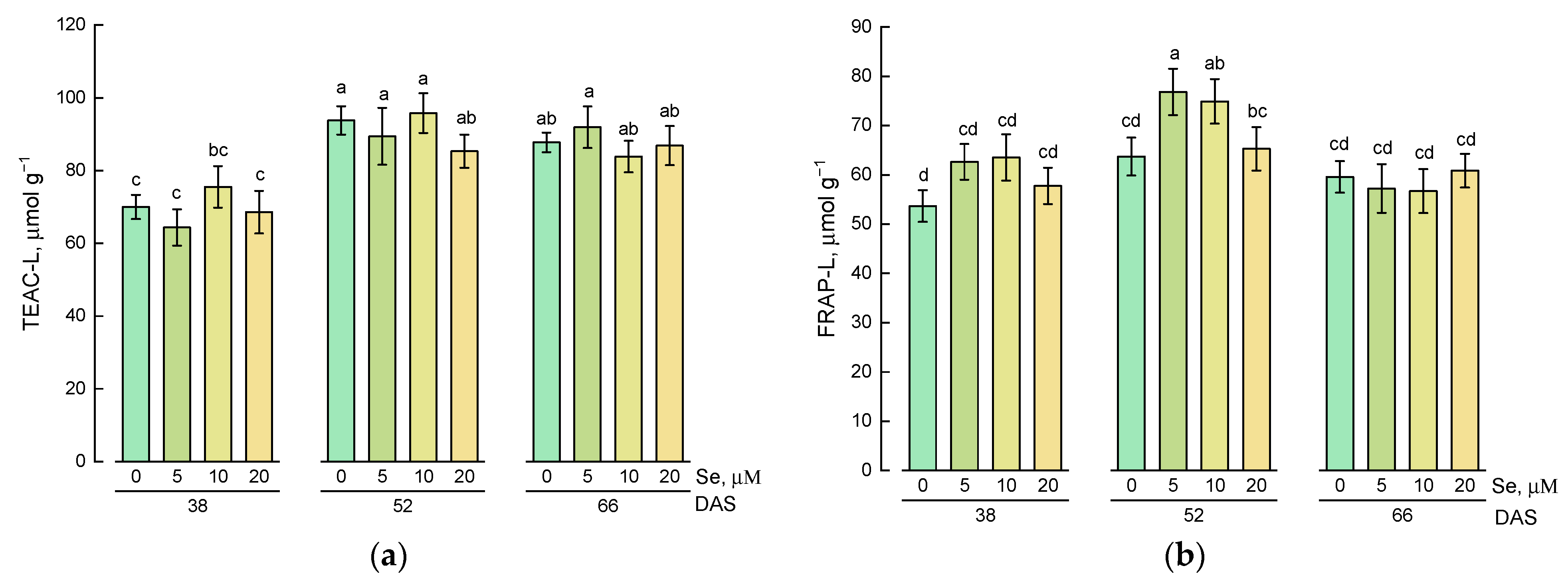
2.2. Effect of Se Application and Growth Stage at Harvest on the Content of Hydrophilic Antioxidative Compounds
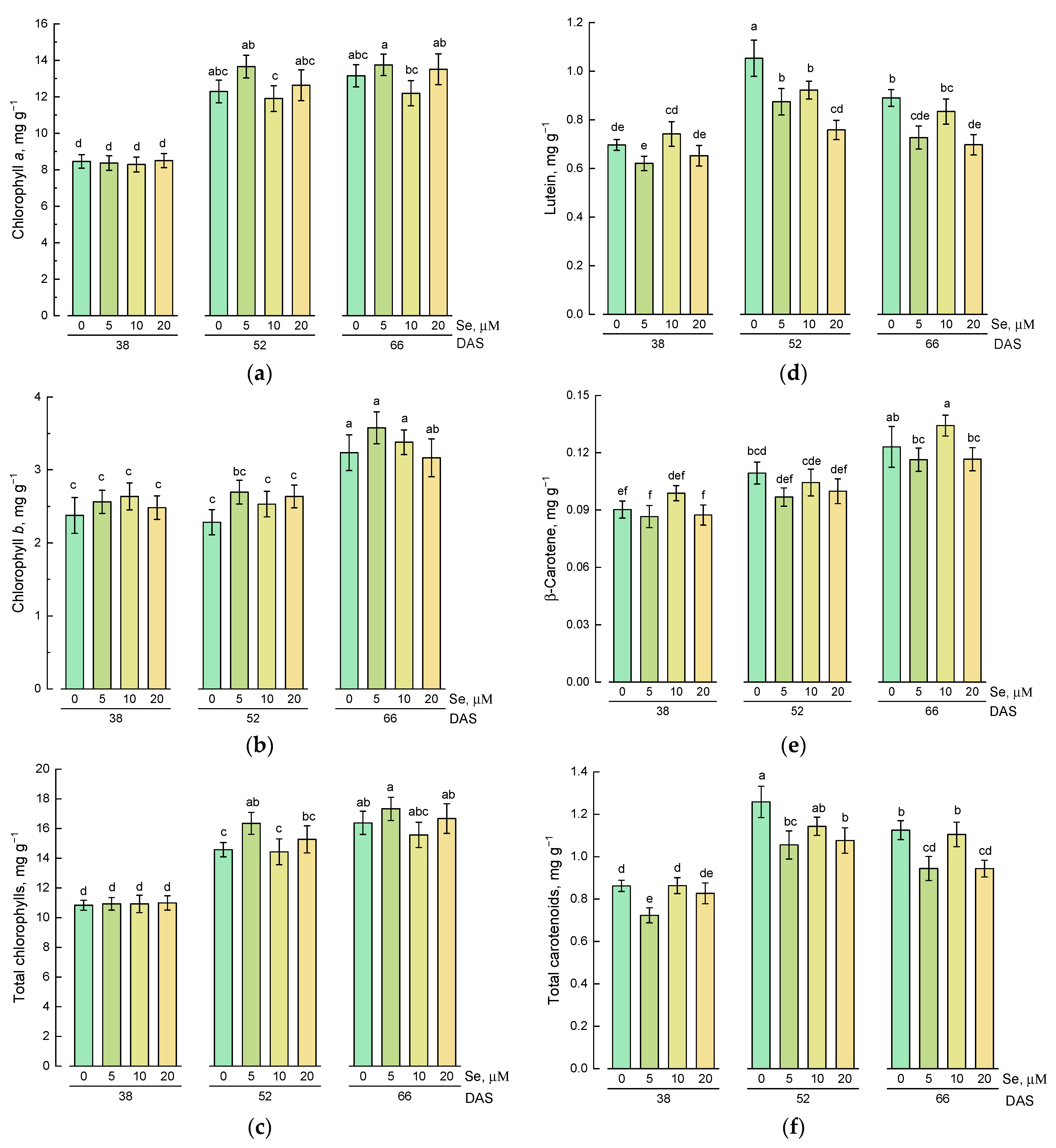
2.3. Effect of Se Application and Growth Stage at Harvest on Lipophilic Antioxidative Compounds
3. Discussion
3.1. Effect of Exogenous Se Application on Se Accumulation in Lamb’s Lettuce Shoots and Plant Yield
3.2. Effect of Exogenous Se Application on the Content of Hydrophilic and Lipophilic Antioxidants in Lamb’s Lettuce Shoots
3.3. Changes in the Content of Hydrophilic and Lipophilic Antioxidants during Plant Growth
4. Materials and Methods
4.1. Plant Materials
4.2. Determination of Se Concentration
4.3. Determination of Vitamin C
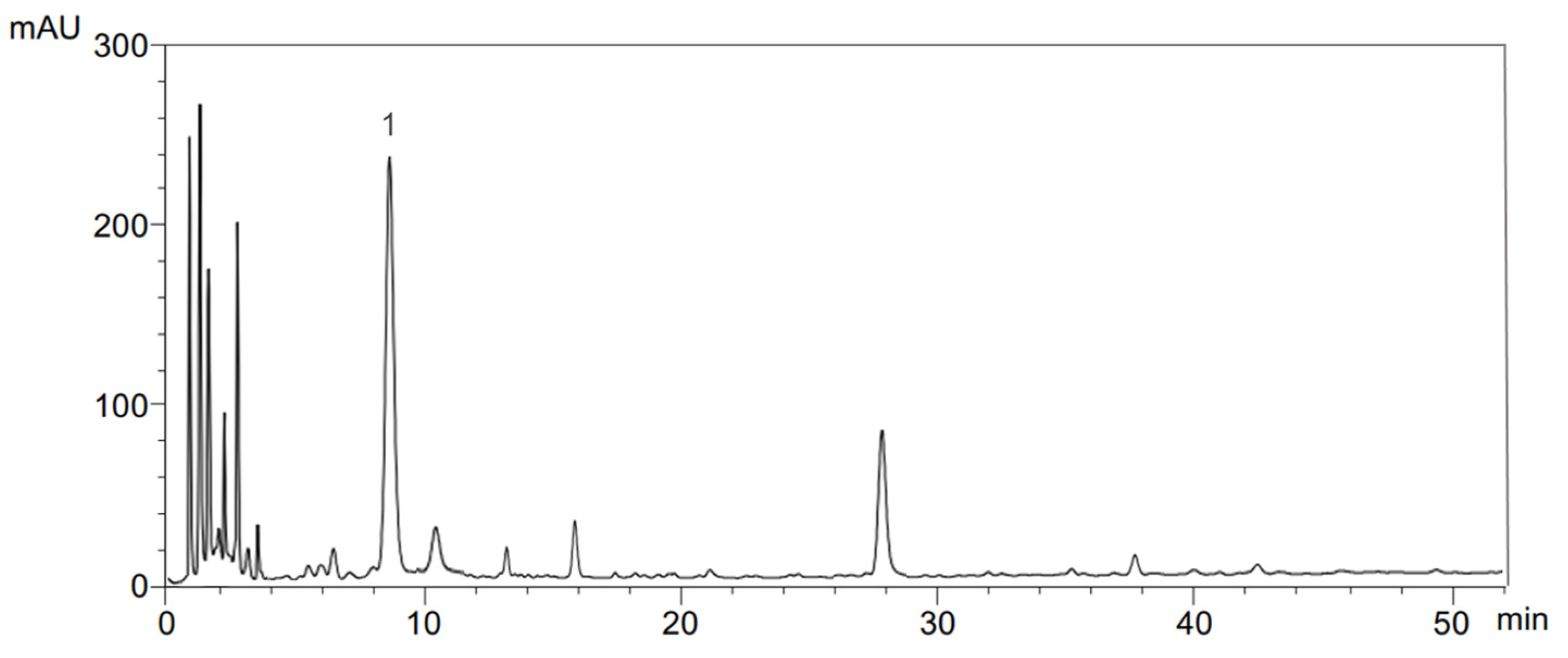
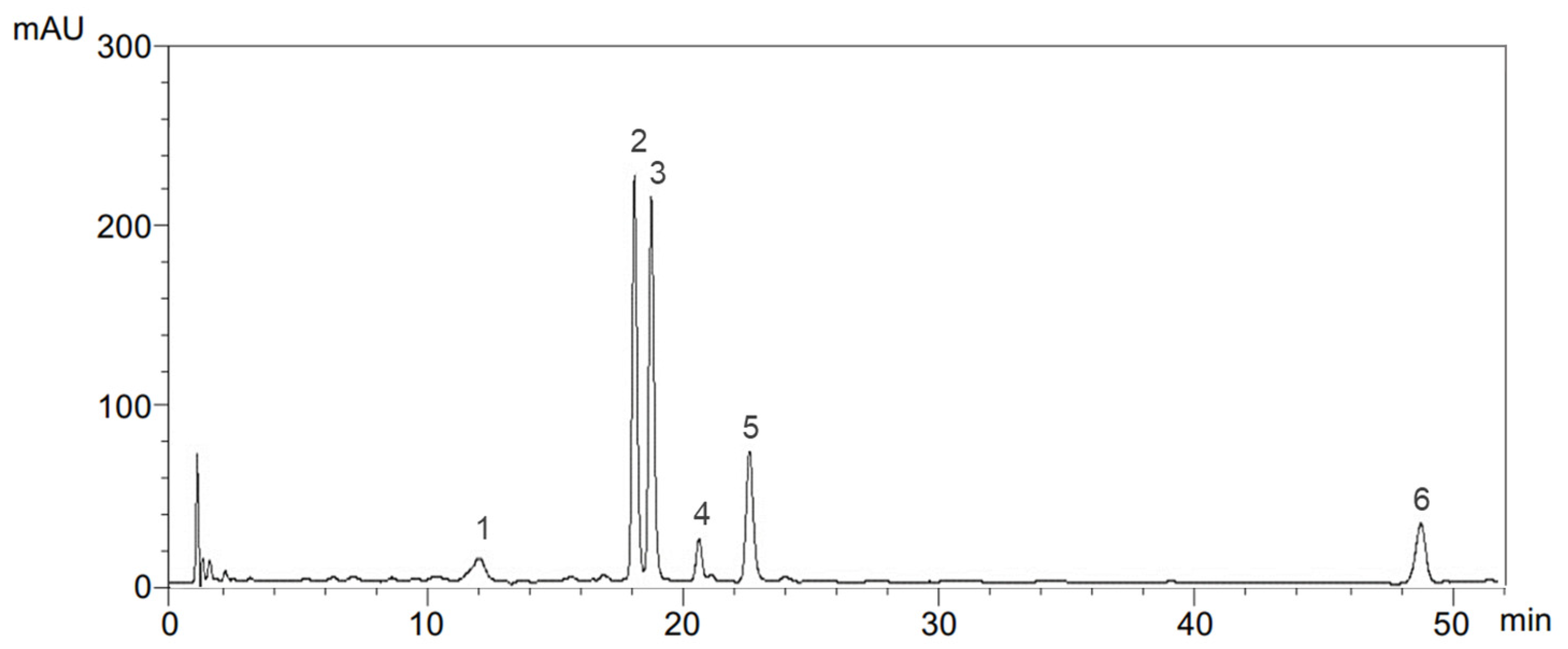
4.4. Determination of Phenolic Compounds
4.5. Determination of Chlorophylls and Carotenoids
4.6. Determination of the Antioxidant Activity of Hydrophilic and Lipophilic Extracts
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Level | Chlorogenic Acid mg g−1 | THA mg g−1 | TFC mg g−1 | TPC mg g−1 | AsA mg g−1 | TEAC -H µmol g−1 | FRAP-H µmol g−1 |
|---|---|---|---|---|---|---|---|---|
| Main effects 1 | ||||||||
| Se concentration (Se) | 0 (control) | 5.907 c | 6.358 c | 1.386 c | 10.353 c | 5.284 c | 74.293 b | 132.102 c |
| 5 µM | 7.499 a | 8.034 a | 1.579 b | 12.946 a | 6.845 b | 99.434 a | 172.775 a | |
| 10 µM | 6.295 b | 6.796 b | 1.626 ab | 11.228 b | 6.859 b | 75.601 b | 149.857 b | |
| 20 µM | 5.667 c | 6.007 c | 1.691 a | 10.221 c | 7.309 a | 74.910 b | 146.069 b | |
| Harvest (H) | 38 DAS | 6.964 a | 7.220 a | 1.808 a | 12.797 a | 6.401 a | 88.887 a | 158.935 a |
| 52 DAS | 6.606 b | 6.483 b | 1.591 b | 11.495 b | 6.727 a | 81.971 b | 150.513 b | |
| 66 DAS | 5.456 c | 5.370 c | 1.313 c | 9.269 c | 6.595 a | 72.320 c | 141.069 c | |
| Significance | Se | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * |
| H | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.072 ns | <0.001 * | <0.001 * | |
| Se * H | 0.078 ns | 0.079 ns | 0.114 ns | 0.011 * | 0.009 * | <0.001 * | 0.006 * |
| Factors | Level | Chlorophyll a mg g−1 | Chlorophyll b mg g−1 | Total Chlorophyllsmg g−1 | Luteinmg g−1 | β-Carotenemg g−1 | Total carotenoidsmg g−1 | TEAC-L µmol g−1 | FRAP-L µmol g−1 |
|---|---|---|---|---|---|---|---|---|---|
| Main effects 1 | |||||||||
| Se concentration (Se) | 0 (control) | 11.300 ab | 2.632 b | 13.932 b | 0.879 a | 0.108 ab | 1.082 a | 83.829 a | 58.987 b |
| 5 µM | 11.923 a | 2.944 a | 14.867 a | 0.740 b | 0.0999 c | 0.908 b | 81.883 a | 65.559 a | |
| 10 µM | 10.794 b | 2.849 a | 13.643 b | 0.833 a | 0.112 a | 1.037 a | 85.036 a | 65.054 a | |
| 20 µM | 11.550 a | 2.761 ab | 14.311 ab | 0.702 b | 0.101 bc | 0.949 b | 80.246 a | 61.300 ab | |
| Harvest (H) | 38 DAS | 8.405 c | 2.514 b | 10.919 c | 0.678 c | 0.0908 b | 0.819 c | 69.594 b | 59.402 b |
| 52 DAS | 12.620 b | 2.536 b | 15.157 b | 0.902 a | 0.103 b | 1.333 a | 91.070 a | 70.181 a | |
| 66 DAS | 13.151 a | 3.339 a | 16.490 a | 0.787 b | 0.123 a | 1.030 b | 87.582 a | 58.592 b | |
| Significance | Se | <0.001 * | 0.003 * | 0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.126 ns | <0.001 * |
| H | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | |
| Se * H | 0.098 ns | 0.268 ns | 0.131 ns | <0.001 * | 0.193 ns | 0.031 * | 0.015 * | 0.001 * |
Appendix B


References
- Ramos-Bueno, R.P.; Rincón-Cervera, M.A.; González-Fernández, M.J.; Guil-Guerrero, J.L. Phytochemical Composition and Antitumor Activities of New Salad Greens: Rucola (Diplotaxis tenuifolia) and Corn Salad (Valerianella locusta). Plant Foods Hum. Nutr. 2016, 71, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Hernández, V.; Botella, M.Á.; Hellín, P.; Cava, J.; Fenoll, J.; Mestre, T.; Martínez, V.; Flores, P. Phenolic and Carotenoid Profile of Lamb’s Lettuce and Improvement of the Bioactive Content by Preharvest Conditions. Foods 2021, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Alvino, A.; Barbieri, G. Vegetables of temperate climates: Leafy vegetables. In The Encyclopedia of Food and Health, 1st ed.; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; Volume 5, pp. 393–400. [Google Scholar]
- Manzocco, L.; Foschia, M.; Tomasi, N.; Maifreni, M.; Dalla Costa, L.; Marino, M.; Cortella, G.; Cesco, S. Influence of hydroponic and soil cultivation on quality and shelf life of ready-to-eat lamb's lettuce (Valerianella locusta L. Laterr). J. Sci. Food Agric. 2011, 91, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Iwamoto, T.; Carey, E.E. Antioxidant capacity of leafy vegetables as affected by high tunnel environment, fertilisation and growth stage. J. Sci. Food Agric. 2007, 87, 2692–2699. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Variation in Macronutrient Content, Phytochemical Constitution and In Vitro Antioxidant Capacity of Green and Red Butterhead Lettuce Dictated by Different Developmental Stages of Harvest Maturity. Antioxidants 2020, 9, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mediani, A.; Abas, F.; Ping, T.C.; Khatib, A.; Lajis, N.H. Influence of growth stage and season on the antioxidant constituents of Cosmos caudatus. Plant Foods Hum. Nutr. 2012, 67, 344–350. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Giordano, M.; Kyriacou, M.C.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Mineral and Antioxidant Attributes of Petroselinum crispum at Different Stages of Ontogeny: Microgreens vs. Baby Greens. Agronomy 2021, 11, 857. [Google Scholar] [CrossRef]
- Uddin, M.K.; Juraimi, A.S.; Ali, M.E.; Ismail, M.R. Evaluation of Antioxidant Properties and Mineral Composition of Purslane (Portulaca oleracea L.) at Different Growth Stages. Int. J. Mol. Sci. 2012, 13, 10257–10267. [Google Scholar] [CrossRef]
- Yadav, L.P.; Koley, T.K.; Tripathi, A.; Singh, S. Antioxidant Potentiality and Mineral Content of Summer Season Leafy Greens: Comparison at Mature and Microgreen Stages Using Chemometric. Agric. Res. 2019, 8, 165–175. [Google Scholar] [CrossRef]
- Długosz-Grochowska, O.; Wojciechowska, R.; Kruczek, M.; Habela, A. Supplemental lighting with LEDs improves the biochemical composition of two Valerianella locusta (L.) cultivars. Hortic. Environ. Biotechnol. 2017, 58, 441–449. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Nocerino, S.; Rouphael, Y.; Colla, G.; El-Nakhel, C.; Mori, M. Nitrogen Use and Uptake Efficiency and Crop Performance of Baby Spinach (Spinacia oleracea L.) and Lamb’s Lettuce (Valerianella locusta L.) Grown under Variable Sub-Optimal N Regimes Combined with Plant-Based Biostimulant Application. Agronomy 2020, 10, 278. [Google Scholar] [CrossRef] [Green Version]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Schiavon, M.; Nardi, S.; Dalla Vecchia, F.; Ertani, A. Selenium biofortification in the 21 st century: Status and challenges for healthy human nutrition. Plant Soil 2020, 453, 245–270. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.; Moldovan, A.; Kekina, H.; Kharchenko, V.; Sekara, A.; Vasileva, V.; Skrypnik, L.; Tallarita, A.; Caruso, G. Joint Biofortification of Plants with Selenium and Iodine: New Field of Discoveries. Plants 2021, 10, 1352. [Google Scholar] [CrossRef] [PubMed]
- García Márquez, V.; Morelos Moreno, Á.; Benavides Mendoza, A.; Medrano Macías, J. Ionic Selenium and Nanoselenium as Biofortifiers and Stimulators of Plant Metabolism. Agronomy 2020, 10, 1399. [Google Scholar] [CrossRef]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current knowledge on selenium biofortification to improve the nutraceutical profile of food: A comprehensive review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef]
- Wrobel, K.; Esperanza, M.G.; Barrientos, E.Y.; Escobosa, A.R.C.; Wrobel, K. Different approaches in metabolomic analysis of plants exposed to selenium: A comprehensive review. Acta Physiol. Plant. 2020, 42, 125. [Google Scholar] [CrossRef]
- Malagoli, M.; Schiavon, M.; Pilon-Smits, E.A. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasi, N.; Pinton, R.; Gottardi, S.; Mimmo, T.; Scampicchio, M.; Cesco, S. Selenium fortification of hydroponically grown corn salad (Valerianella locusta). Crop and Pasture Science 2015, 66, 1128–1136. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Dresler, S.; Rubinowska, K.; Matraszek-Gawron, R.; Woch, W.; Hasanuzzaman, M. Selenium biofortification enhances the growth and alters the physiological response of lamb's lettuce grown under high temperature stress. Plant Physiol. Biochem. 2018, 127, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H. On the Ecology of Selenium Accumulation in Plants. Plants 2019, 8, 197. [Google Scholar] [CrossRef] [Green Version]
- Saffaryazdi, A.; Lahouti, M.; Ganjeali, A.; Bayat, H. Impact of selenium supplementation on growth and selenium accumulation on spinach (Spinacia oleracea L.) plants. Not. Sci. Biol. 2012, 4, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Bo, L.E.I.; Bian, Z.H.; Yang, Q.C.; Jun, W.A.N.G.; Cheng, R.F.; Kun, L.I.; Liu, W.; Zhnag, Y.; Fang, H.; Tong, Y.X. The positive function of selenium supplementation on reducing nitrate accumulation in hydroponic lettuce (Lactuca sativa L.). J. Integr. Agric. 2018, 17, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Golubkina, N.; Kekina, H.; Caruso, G. Yield, Quality and Antioxidant Properties of Indian Mustard (Brassica juncea L.) in Response to Foliar Biofortification with Selenium and Iodine. Plants 2018, 7, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mroczek-Zdyrska, M.; Strubińska, J.; Hanaka, A. Selenium improves physiological parameters and alleviates oxidative stress in shoots of lead-exposed Vicia faba L. minor plants grown under phosphorus-deficient conditions. J. Plant Growth Regul. 2017, 36, 186–199. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Niazi, N.K.; Khalid, S.; Murtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 2018. 234, 915–934. [Google Scholar] [CrossRef]
- Regni, L.; Micheli, M.; Del Pino, A.M.; Palmerini, C.A.; D’Amato, R.; Facchin, S.L.; Famiani, F.; Peruzzi, A.; Mairech, H.; Proietti, P. The First Evidence of the Beneficial Effects of Se-Supplementation on In Vitro Cultivated Olive Tree Explants. Plants 2021, 10, 1630. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Rosellini, I.; Pezzarossa, B. Uptake and partitioning of selenium in basil (Ocimum basilicum L.) plants grown in hydroponics. Sci. Hortic. 2017, 225, 271–276. [Google Scholar] [CrossRef]
- Skrypnik, L.; Novikova, A.; Tokupova, E. Improvement of Phenolic Compounds, Essential Oil Content and Antioxidant Properties of Sweet Basil (Ocimum basilicum L.) Depending on Type and Concentration of Selenium Application. Plants 2019, 8, 458. [Google Scholar] [CrossRef] [Green Version]
- Puccinelli, M.; Pezzarossa, B.; Rosellini, I.; Malorgio, F. Selenium Enrichment Enhances the Quality and Shelf Life of Basil Leaves. Plants 2020, 9, 801. [Google Scholar] [CrossRef] [PubMed]
- Hawrylak-Nowak, B. Enhanced selenium content in sweet basil (Ocimum basilicum) by foliar fertilization. Veg. Crop. Res. Bul. 2008, 69, 63–72. [Google Scholar] [CrossRef]
- Liu, D.; Li, H.; Wang, Y.; Ying, Z.; Bian, Z.; Zhu, W.; Liu, W.; Yang, L.; Jiang, D. How Exogenous Selenium Affects Anthocyanin Accumulation and Biosynthesis-Related Gene Expression in Purple Lettuce. Pol. J. Environ. Stud. 2017, 26, 717–722. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Pannico, A.; El-Nakhel, C.; Kyriacou, M.C.; Giordano, M.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Combating micronutrient deficiency and enhancing food functional quality through selenium fortification of select lettuce genotypes grown in a closed soilless system. Front. Plant Sci. 2019, 10, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poldma, P.; Moor, U.; Tonutare, T.; Herodes, K.; Rebane, R. Selenium treatment under field conditions affects mineral nutrition, yield and antioxidant properties of bulb onion (Allium cepa L.). Acta Sci. Pol-Hortoru. 2013, 12, 167–181. [Google Scholar]
- Zhu, Z.; Zhang, Y.; Liu, J.; Chen, Y.; Zhang, X. Exploring the effects of selenium treatment on the nutritional quality of tomato fruit. Food Chem. 2018, 252, 9–15. [Google Scholar] [CrossRef]
- Babajani, A.; Iranbakhsh, A.; Ardebili, Z.O.; Eslami, B. Differential growth, nutrition, physiology, and gene expression in Melissa officinalis mediated by zinc oxide and elemental selenium nanoparticles. Environ. Sci. Pollut. Res. 2019, 26, 24430–24444. [Google Scholar] [CrossRef] [PubMed]
- Mimmo, T.; Tiziani, R.; Valentinuzzi, F.; Lucini, L.; Nicoletto, C.; Sambo, P.; Scampicchio, M.; Pii, Y.; Cesco, S. Selenium biofortification in Fragaria× ananassa: Implications on strawberry fruits quality, content of bioactive health beneficial compounds and metabolomic profile. Front. Plant Sci. 2017, 8, 1887. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.J.; Rosales, M.A.; Blasco, B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Sabatino, L.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C. Effect of Selenium Enrichment and Type of Application on Yield, Functional Quality and Mineral Composition of Curly Endive Grown in a Hydroponic System. Agronomy 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Newman, R.; Waterland, N.; Moon, Y.; Tou, J.C. Selenium Biofortification of Agricultural Crops and Effects on Plant Nutrients and Bioactive Compounds Important for Human Health and Disease Prevention—A Review. Plant Foods Hum. Nutr. 2019, 74, 449–460. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmiański, J.; Wiśniewski, R. Determination of triterpenoids, carotenoids, chlorophylls, and antioxidant capacity in Allium ursinum L. at different times of harvesting and anatomical parts. Eur. Food Res. Technol. 2018, 244, 1269–1280. [Google Scholar] [CrossRef] [Green Version]
- Malorgio, F.; Diaz, K.E.; Ferrante, A.; Mensuali-Sodi, A.; Pezzarossa, B. Effects of selenium addition on minimally processed leafy vegetables grown in a floating system. J. Sci. Food Agric. 2009, 89, 2243–2251. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium Biofortification Impacts the Nutritive Value, Polyphenolic Content, and Bioactive Constitution of Variable Microgreens Genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefsrud, M.G.; Kopsell, D.A.; Kopsell, D.E.; Randle, W.M. Kale carotenoids are unaffected by, whereas biomass production, elemental concentrations, and selenium accumulation respond to, changes in selenium fertility. J. Agric. Food Chem. 2006, 54, 1764–1771. [Google Scholar] [CrossRef]
- Sams, C.E.; Panthee, D.R.; Charron, C.S.; Kopsell, D.A.; Yuan, J.S. Selenium regulates gene expression for glucosinolate and carotenoid biosynthesis in Arabidopsis. J. Am. Soc. Hortic. Sci. 2011, 136, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Heo, H.J.; Kim, Y.J.; Chung, D.; Kim, D.O. Antioxidant capacities of individual and combined phenolics in a model system. Food Chem. 2007, 104, 87–92. [Google Scholar] [CrossRef]
- Olszowy-Tomczyk, M. Synergistic, antagonistic and additive antioxidant effects in the binary mixtures. Phytochem. Rev. 2020, 19, 63–103. [Google Scholar] [CrossRef]
- White, P.J. Selenium metabolism in plants. Biochim. Biophys. Acta (BBA) Gen. Subj. 2018, 1862, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Del Bano, M.J.; Lorente, J.; Castillo, J.; Benavente-García, O.; Del Rio, J.A.; Ortuño, A.; Quirin, K.-W.; Gerard, D. Phenolic diterpenes, flavones, and rosmarinic acid distribution during the development of leaves, flowers, stems, and roots of Rosmarinus officinalis. Antioxidant activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef] [PubMed]
- Lisiewska, Z.; Kmiecik, W.; Korus, A. Content of vitamin C, carotenoids, chlorophylls and polyphenols in green parts of dill (Anethum graveolens L.) depending on plant height. J. Food Compost. Anal. 2006, 19, 134–140. [Google Scholar] [CrossRef]
- Nordmark, L.; Gertsson, U.; Olsson, K.; Olsson, M.E. Content of bioactive compounds in baby-leaves as affected by season and growth stage at harvest. Acta Hortic. 2014, 1040, 201–206. [Google Scholar] [CrossRef]
- Lefsrud, M.; Kopsell, D.; Wenzel, A.; Sheehan, J. Changes in kale (Brassica oleracea L. var. acephala) carotenoid and chlorophyll pigment concentrations during leaf ontogeny. Sci. Hortic. 2007, 112, 136–141. [Google Scholar] [CrossRef]
- Kurkova, T.; Skrypnik, L.; Zalieckiene, E. Features of plant material pre-treatment for the selenium determination by atomic absorption and fluorometric methods. Chemija 2008, 19, 40–43. [Google Scholar]
- Galani, J.H.Y.; Patel, J.S.; Patel, N.J.; Talati, J.G. Storage of Fruits and Vegetables in Refrigerator Increases their Phenolic Acids but Decreases the Total Phenolics, Anthocyanins and Vitamin C with Subsequent Loss of their Antioxidant Capacity. Antioxidants 2017, 6, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Santos, E.C.E.; Viganó, J.; Neves, D.A.; Martínez, J.; Godoy, H.T. Vitamin C in camu-camu [Myrciaria dubia (HBK) McVaugh]: Evaluation of extraction and analytical methods. Food Res. Int. 2019, 115, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Neugart, S.; Majer, P.; Schreiner, M.; Hideg, É. Blue light treatment but not green light treatment after pre-exposure to UV-B stabilizes flavonoid glycoside changes and corresponding biological effects in three different Brassicaceae sprouts. Front. Plant Sci. 2020, 11, 611247. [Google Scholar] [CrossRef]
- Štefan, M.B.; Rodríguez, J.V.; Blažeković, B.; Kindl, M.; Vladimir-Knežević, S. Total hydroxycinnamic acids assay: Prevalidation and application on Lamiaceae species. Food Anal. Methods 2014, 7, 326–336. [Google Scholar] [CrossRef]
- Sevket, A.L.P.; Ercisli, S.; Jurikova, T.; Cakir, O.; Gozlekci, S. Bioactive content of rose hips of different wildly grown Rosa dumalis genotypes. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 472–476. [Google Scholar] [CrossRef] [Green Version]
- Padhi, E.M.; Liu, R.; Hernandez, M.; Tsao, R.; Ramdath, D.D. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J. Funct. Foods 2017, 38, 602–611. [Google Scholar] [CrossRef]
- Frede, K.; Schreiner, M.; Baldermann, S. Light quality-induced changes of carotenoid composition in pak choi Brassica rapa ssp. chinensis. J. Photochem. Photobiol. B Biol. 2019, 193, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Skrypnik, L.; Novikova, A. Response Surface Modeling and Optimization of Polyphenols Extraction from Apple Pomace Based on Nonionic Emulsifiers. Agronomy 2020, 10, 92. [Google Scholar] [CrossRef] [Green Version]






| Growth Stage at Harvest | Se Concentration, µM | Se Concentration µg g−1 | Se Content µg Plant−1 | FW Shoots g Plant−1 | DW Shoots g Plant−1 | Dry Matter Content % |
|---|---|---|---|---|---|---|
| 38 DAS | 0 | 0.051 f 1 | 0.009 f | 2.21 e | 0.186 d | 8.42 a |
| 5 | 2.11 ef | 0.388 f | 2.19 e | 0.186 d | 8.51 a | |
| 10 | 11.58 d | 2.27 ef | 2.23 e | 0.195 d | 8.78 a | |
| 20 | 39.29 c | 6.86 de | 2.16 e | 0.175 d | 8.17 a | |
| 52 DAS | 0 | 0.018 f | 0.009 f | 6.64 d | 0.533 c | 8.03 a |
| 5 | 6.29 def | 3.71 ef | 7.18 d | 0.591 c | 8.23 a | |
| 10 | 39.3 c | 21.6 c | 6.77 d | 0.550 c | 8.13 a | |
| 20 | 124.4 a | 65.1 a | 6.58 d | 0.524 c | 7.97 a | |
| 66 DAS | 0 | 0.011 f | 0.010 f | 11.62 b | 0.899 b | 7.76 a |
| 5 | 10.7 de | 12.28 d | 14.72 a | 1.201 a | 8.14 a | |
| 10 | 37.2 c | 31.5 b | 10.99 bc | 0.850 b | 7.73 a | |
| 20 | 90.8 b | 68.9 a | 9.77 c | 0.763 b | 7.80 a | |
| Main effects 2 | ||||||
| Se concentration (Se) | 0 (control) | 0.0264 d | 0.009 d | 6.82 b | 0.539 b | 8.07 a |
| 5 µM | 6.22 c | 5.46 c | 8.0. a | 0.660 a | 8.30 a | |
| 10 µM | 29.34 b | 18.48 b | 6.66 bc | 0.531 b | 8.21 a | |
| 20 µM | 84.83 a | 46.98 a | 6.17 c | 0.488 b | 7.98 a | |
| Harvest (H) | 38 DAS | 13.26 c | 2.38 c | 2.20 c | 0.186 c | 8.47 a |
| 52 DAS | 42.52 a | 22.63 b | 6.79 b | 0.550 b | 8.09 ab | |
| 66 DAS | 34.58 b | 28.20 a | 11.78 a | 0.929 a | 7.86 b | |
| Significance | Se | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.424 ns |
| H | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.005 * | |
| Se * H | <0.001 * | <0.001 * | <0.001 * | <0.001 * | 0.850 ns |
| Parameters | THA 1 | TFC | TPC | AsA | TEAC-H | FRAP-H |
|---|---|---|---|---|---|---|
| ChlA | 0.99 ** | 0.52 * | 0.96 ** | 0.06 ns | 0.80 ** | 0.75 ** |
| THA | 0.54 * | 0.97 ** | 0.03 ns | 0.79 ** | 0.74 ** | |
| TFC | 0.68 ** | 0.35 * | 0.47 * | 0.52 * | ||
| TPC | 0.08 ns | 0.79 ** | 0.76 ** | |||
| AsA | 0.19 ns | 0.47 * | ||||
| TEAC-H | 0.85 ** |
| Parameters | Chl b 1 | TChl | Lut | β-Car | TCar | TEAC-L | FRAP-L |
|---|---|---|---|---|---|---|---|
| Chl a | 0.56 * | 0.98 ** | 0.51 * | 0.64 ** | 0.66 ** | 0.82 ** | 0.30 * |
| Chl b | 0.67 ** | −0.04 ns | 0.70 ** | 0.10 ns | 0.35 * | −0.18 ns | |
| TChl | 0.45 * | 0.69 ** | 0.61 ** | 0.80 ** | 0.23 ns | ||
| Lut | 0.42 * | 0.92 ** | 0.74 ** | 0.44 * | |||
| β-Car | 0.57 * | 0.56 * | −0.11 ns | ||||
| TCar | 0.80 ** | 0.35 * | |||||
| TEAC-L | 0.48 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skrypnik, L.; Styran, T.; Savina, T.; Golubkina, N. Effect of Selenium Application and Growth Stage at Harvest on Hydrophilic and Lipophilic Antioxidants in Lamb’s Lettuce (Valerianella locusta L. Laterr.). Plants 2021, 10, 2733. https://doi.org/10.3390/plants10122733
Skrypnik L, Styran T, Savina T, Golubkina N. Effect of Selenium Application and Growth Stage at Harvest on Hydrophilic and Lipophilic Antioxidants in Lamb’s Lettuce (Valerianella locusta L. Laterr.). Plants. 2021; 10(12):2733. https://doi.org/10.3390/plants10122733
Chicago/Turabian StyleSkrypnik, Liubov, Tatiana Styran, Tamara Savina, and Nadezhda Golubkina. 2021. "Effect of Selenium Application and Growth Stage at Harvest on Hydrophilic and Lipophilic Antioxidants in Lamb’s Lettuce (Valerianella locusta L. Laterr.)" Plants 10, no. 12: 2733. https://doi.org/10.3390/plants10122733