
Assessment of Genetic Diversity and Population Structure of the Endangered Astragalus exscapus subsp. transsilvanicus through DNA-Based Molecular Markers


Abstract
:1. Introduction
2. Results
2.1. Morphologic Description of Eight Populations of A. exscapus subsp. transsilvanicus
2.2. SRAP Marker Polymorphism
2.3. Genetic Diversity Analysis
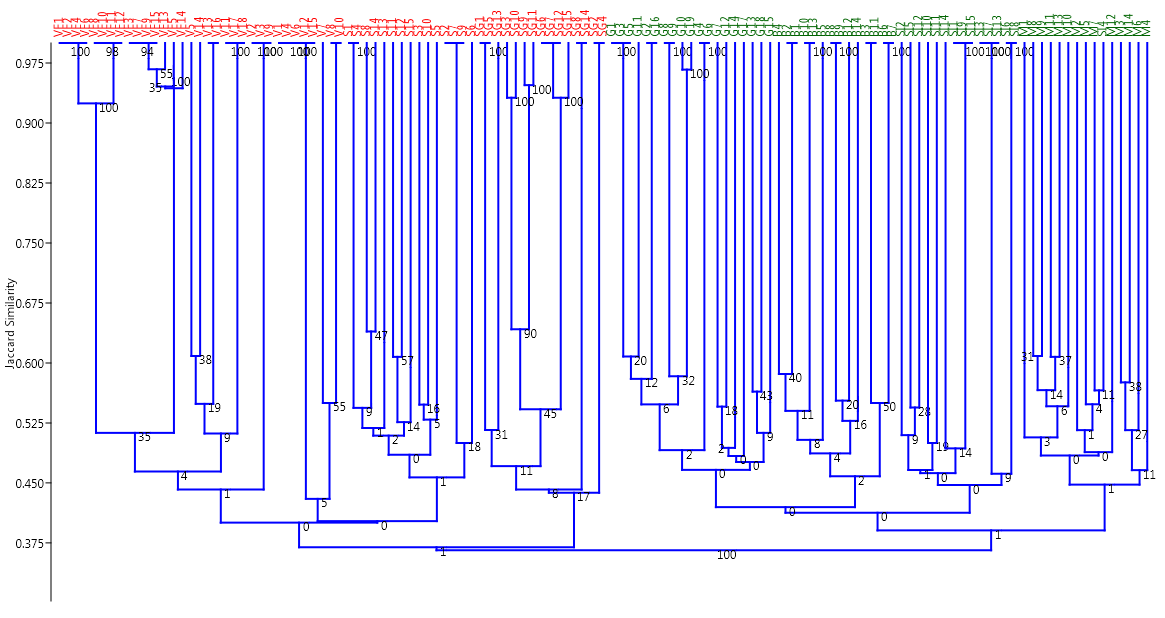
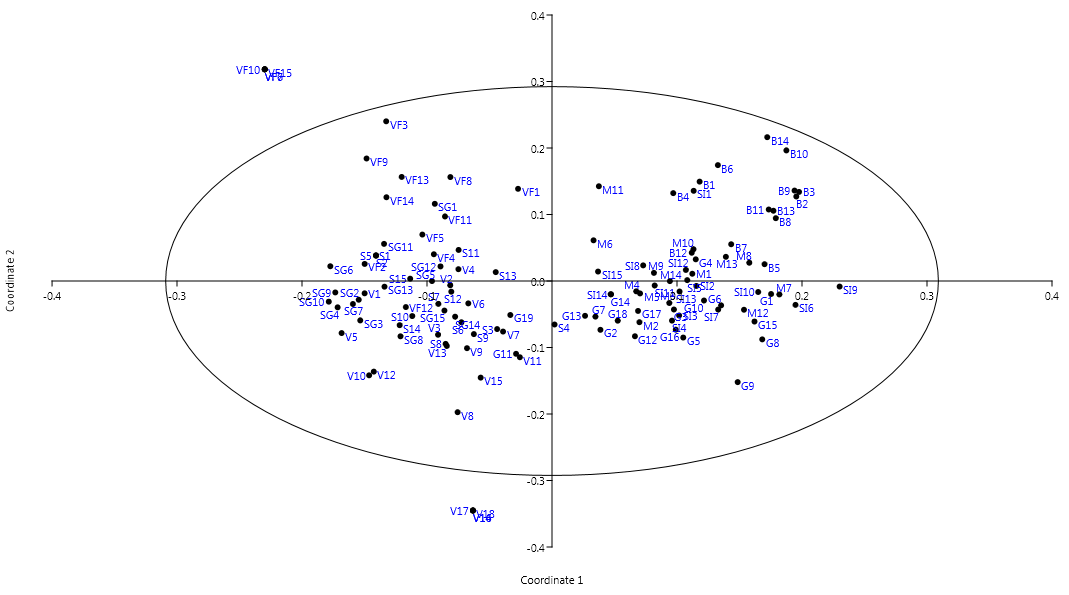
2.4. Cluster Analysis and PCoA
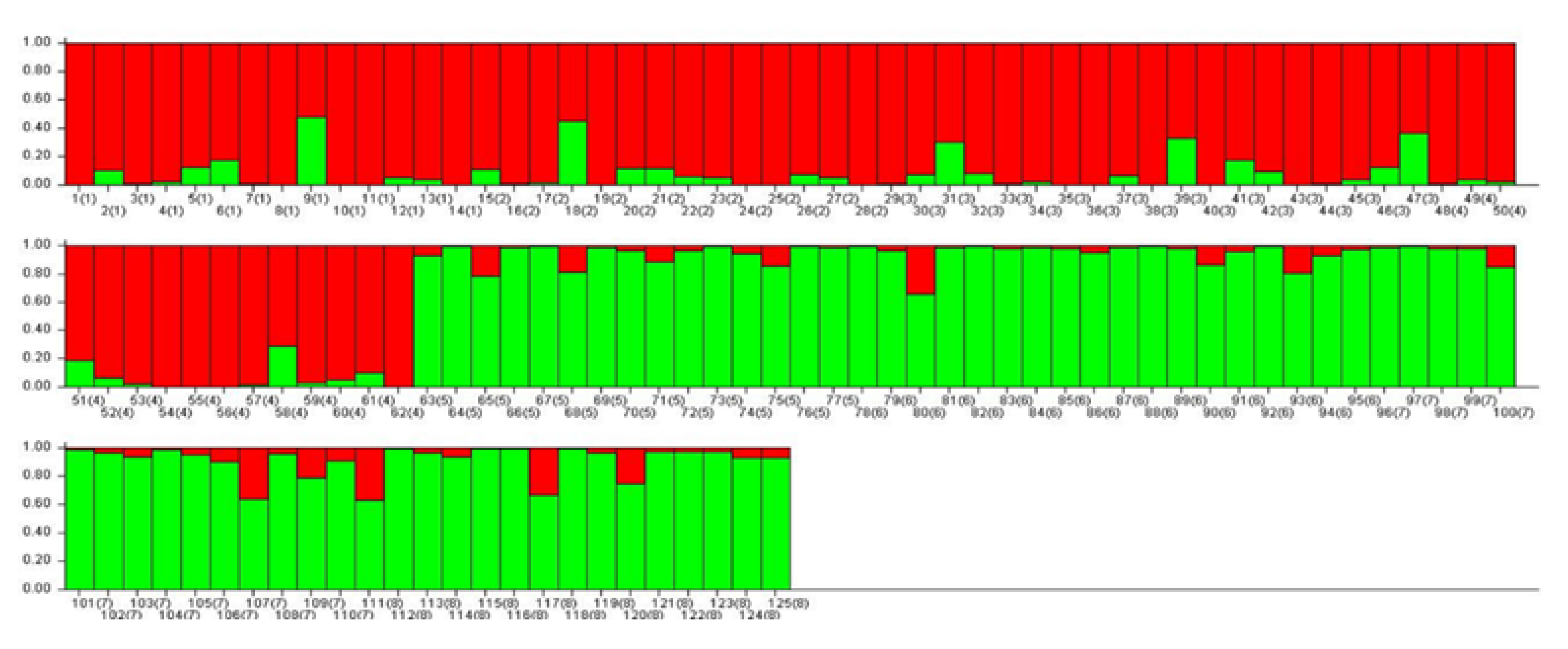
2.5. Population Structure Analysis
3. Discussion
3.1. Effectiveness of SRAP Markers in A. exscapus subsp. transsilvanicus
3.2. Genetic Diversity of A. exscapus subsp. transsilvanicus Populations
3.3. Population Structure of A. exscapus subsp. transsilvanicus
3.4. Conservation Implications
4. Materials and Methods
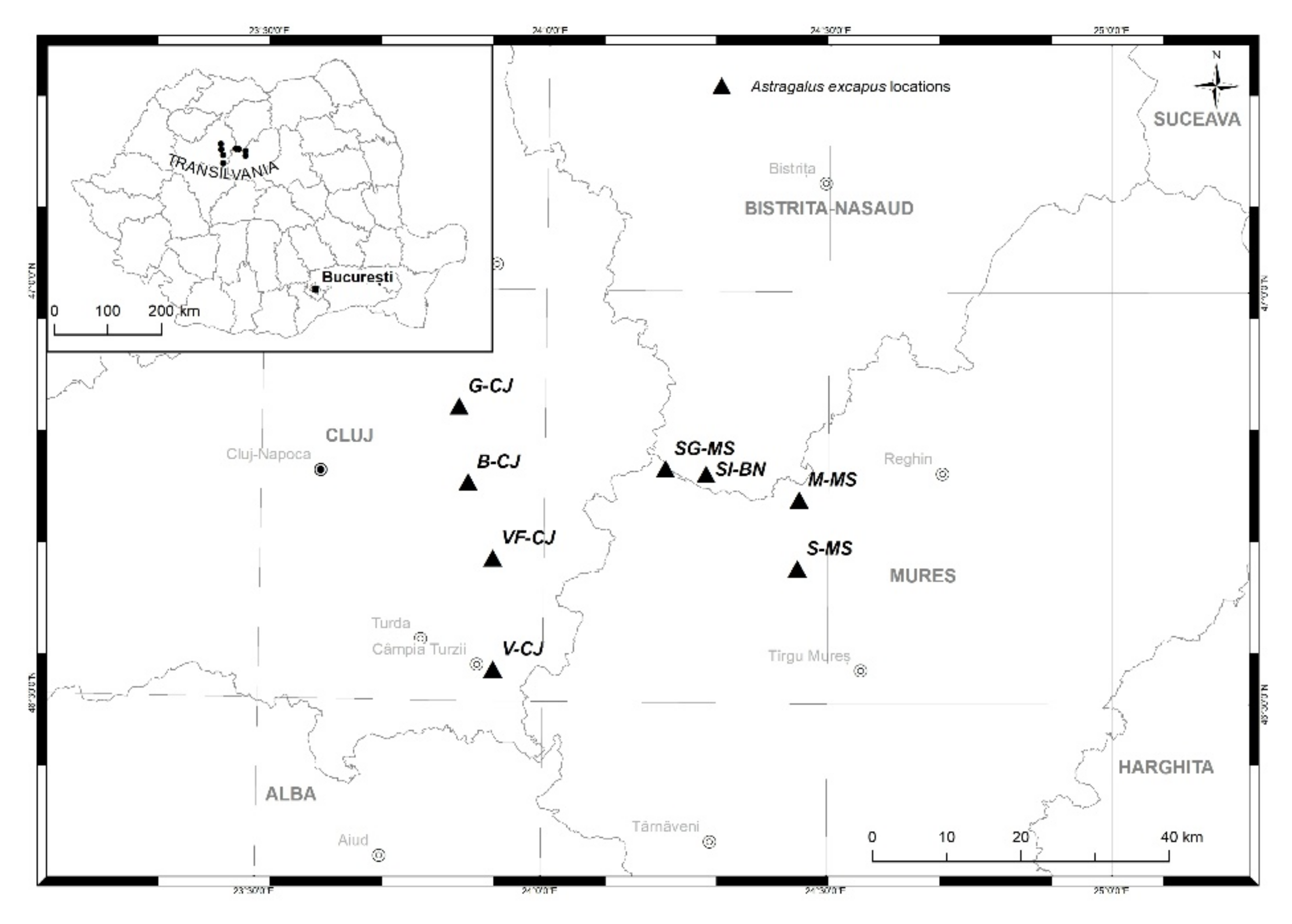
4.1. Plant Material
4.2. DNA Isolation
4.3. SRAP Analysis
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rabinowitz, D.; Cairns, S.; Dillon, T. Seven forms of rarity and their frequency in the flora of the British Isles. In Conservation Biology: The Science of Scarcity and Diversity; Soule, M.E., Ed.; Sinauer Associates: Sunderland, MA, USA, 1986; pp. 182–204. ISBN 0-87893-794-3. [Google Scholar]
- Becker, T.; Voss, N.; Durka, W. Pollen limitation and inbreeding depression in an “old rare” bumblebee-pollinated grassland herb. Plant Biol. 2011, 13, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Becker, T. Explaining Rarity of the Dry Grassland Perennial Astragalus exscapus. Folia Geobot. Phytotaxon. 2010, 45, 303–321. [Google Scholar] [CrossRef] [Green Version]
- Kukuła, K.; Okarma, H.; Pawłowski, J.; Perzanowski, K.; Ruzicka, T.; Sandor, J.; Stanova, V.; Tasenkevich, L.; Vlasin, M. Carpathian List of Endangered Species; WWF International Danube-Carpathian Programme: Vienna, Austria; Institute of Nature Conservation, Polish Academy of Sciences: Krakow, Poland, 2003; pp. 1–64. ISBN 83–918914–0–2. [Google Scholar]
- Vojnits, A.M.; Acs, E. A population of the hungarian zephyr blue. Holarct. Lepid. 1995, 2, 23–26. [Google Scholar]
- Shao, B.-M.; Xu, W.; Dai, H.; Tu, P.; Li, Z.; Gao, X.-M. A study on the immune receptors for polysaccharides from the roots of Astragalus membranaceus, a Chinese medicinal herb. Biochem. Biophys. Res. Commun. 2004, 320, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S. Review of the Botanical Characteristics, Phytochemistry, and Pharmacology of Astragalus membranaceus (Huangqi). Phytother. Res. 2014, 1283, 1275–1283. [Google Scholar] [CrossRef]
- Ganzera, M.; Bedir, E.; Calis, I.; Khan, I.A. Separation of Astragalus saponins by reversed phase high performance liquid chromatography and evaporative light scattering detection. Chromatographia 2001, 53, 131–134. [Google Scholar] [CrossRef]
- Tin, M.M.Y.; Cho, C.-H.; Chan, K.; James, A.E.; Ko, J.K.S. Astragalus saponins induce growth inhibition and apoptosis in human colon cancer cells and tumor xenograft. Carcinogenesis 2007, 28, 1347–1355. [Google Scholar] [CrossRef]
- Auyeung, K.K.W.; Cho, C.H.; Ko, J.K.S. A novel anticancer effect of Astragalus saponins: Transcriptional activation of NSAID-activated gene. Int. J. Cancer 2009, 125, 1082–1091. [Google Scholar] [CrossRef]
- Guo, Z.; Lou, Y.; Kong, M.; Luo, Q.; Liu, Z.; Wu, J. A systematic review of phytochemistry, pharmacology and pharmacokinetics on astragali radix: Implications for astragali radix as a personalized medicine. Int. J. Mol. Sci. 2019, 20, 1463. [Google Scholar] [CrossRef] [Green Version]
- Bădărău, A.S.; Dezsi, S.; Comes, O. Cercetări biogeografice asupra speciilor stepice -silvostepice de Astragalus L. din depresiunea Transilvaniei. Studia Univ. Babes-Bolyai Geogr. XLV 2000, 2, 3. [Google Scholar]
- Selyutina, I.Y.; Konichenko, E.S.; Dorogina, O.V.; Sandanov, D.V. Genetic diversity of the endangered endemic milkvetch Astragalus sericeocanus Gontsch., Fabaceae from the Lake Baikal region. Biochem. Syst. Ecol. 2016, 68, 163–169. [Google Scholar] [CrossRef]
- Vicente, M.J.; Segura, F.; Aguado, M.; Migliaro, D.; Franco, J.A.; Martínez-Sánchez, J.J. Genetic diversity of Astragalus nitidiflorus, a critically endangered endemic of SE Spain, and implications for its conservation. Biochem. Syst. Ecol. 2011, 39, 175–182. [Google Scholar] [CrossRef]
- Bagheri, A.; Abbasi, S.; Mahmoodi, M.; Roofigar, A.A.; Blattner, F.R. Genetic structure and conservation status of Astragalus subrecognitus (Fabaceae): A very rare and narrow endemic species. Plant Ecol. Evol. 2020, 153, 101–107. [Google Scholar] [CrossRef]
- Li, G.; Quiros, C.F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica. TAG Theor. Appl. Genet. 2001, 103, 455–461. [Google Scholar] [CrossRef]
- Robarts, D.W.H.; Wolfe, A.D. Sequence-related amplified polymorphism (SRAP) markers: A potential resource for studies in plant molecular biology(1.). Appl. Plant Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef]
- Ma, M.; Wang, T.; Lu, B. Assessment of genetic diversity in Amomum tsao-ko Crevost & Lemarié, an important medicine food homologous crop from Southwest China using SRAP and ISSR markers. Genet. Resour. Crop Evol. 2021, 68, 2655–2667. [Google Scholar] [CrossRef]
- Weising, K.; Nybom, H.; Wolff, K.; Kahl, G. DNA Fingerprinting in Plants; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-0-8493-1488-9. [Google Scholar]
- Alzahib, R.H.; Migdadi, H.M.; Al Ghamdi, A.A.; Alwahibi, M.S.; Afzal, M.; Elharty, E.H.; Alghamdi, S.S. Exploring genetic variability among and within hail tomato landraces based on sequence-related amplified polymorphism markers. Diversity 2021, 13, 135. [Google Scholar] [CrossRef]
- Borsai, O.; Hârta, M.; Szabo, K.; Kelemen, C.D.; Andrecan, F.A.; Codrea, M.M.; Clapa, D. Evaluation of genetic fidelity of in vitro-propagated blackberry plants using RAPD and SRAP molecular markers. Hortic. Sci. 2020, 47, 21–27. [Google Scholar] [CrossRef]
- Clapa, D.; Borsai, O.; Hârța, M.; Bonta, V.; Szabo, K.; Coman, V.; Bobiș, O. Micropropagation, genetic fidelity and phenolic compound production of Rheum rhabarbarum L. Plants 2020, 9, 656. [Google Scholar] [CrossRef]
- Ren, X.; Huang, J.; Liao, B.; Zhang, X.; Jiang, H. Genomic affinities of Arachis genus and interspecific hybrids were revealed by SRAP markers. Genet. Resour. Crop Evol. 2010, 57, 903–913. [Google Scholar] [CrossRef]
- Hârţa, M.; Borsai, O.; Muntean, C.M.; Dina, N.E.; Fǎlǎmaş, A.; Olar, L.E.; Szabo, K.; Pamfil, D.; Ştefan, R. Assessment of genetic relationships between streptocarpus x hybridus v. Parents and f1 progenies using srap markers and ft-ir spectroscopy. Plants 2020, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Levi, A.; Wechter, P.; Massey, L.; Carter, L.; Hopkins, D. An Extended Genetic Linkage Map for Watermelon Based on a Testcross and a BC Population. Am. J. Plant Sci. 2011, 2, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Talebi, M.; Kazemi, M.; Sayed-Tabatabaei, B.E. Molecular diversity and phylogenetic relationships of Pistacia vera, Pistacia atlantica subsp. mutica and Pistacia khinjuk using SRAP markers. Biochem. Syst. Ecol. 2012, 44, 179–185. [Google Scholar] [CrossRef]
- Cai, X.; Feng, Z.; Zhang, X.; Xu, W.; Hou, B.; Ding, X. Genetic diversity and population structure of an endangered Orchid (Dendrobium loddigesii Rolfe) from China revealed by SRAP markers. Sci. Hortic. 2011, 129, 877–881. [Google Scholar] [CrossRef]
- Liu, Z.; Shu, Q.; Wang, L.; Yu, M.; Hu, Y.; Zhang, H.; Tao, Y.; Shao, Y. Genetic diversity of the endangered and medically important Lycium ruthenicum Murr. revealed by sequence-related amplified polymorphism (SRAP) markers. Biochem. Syst. Ecol. 2012, 45, 86–97. [Google Scholar] [CrossRef]
- Rebrean, F.; Fustos, A.; Tǎut, I.; Szabo, K.; Hȃrţa, M.; Pamfil, D.; Rebrean, M.; Sălăgean, T. Genetic diversity of Acer pseudoplatanus L. populations from Transylvania. Rev. Bras. Bot. 2019, 42, 643–650. [Google Scholar] [CrossRef]
- Jones, M.R.; Winkler, D.E.; Massatti, R. The demographic and ecological factors shaping diversification among rare Astragalus species. Divers. Distrib. 2021, 27, 1407–1421. [Google Scholar] [CrossRef]
- Bobo-Pinilla, J.; López-González, N.; Peñas, J. Conservation of genetic diversity in Mediterranean endemic species: Arenaria balearica (Caryophyllaceae). Plant Ecol. Evol. 2020, 153, 348–360. [Google Scholar] [CrossRef]
- Travis, S.E.; Maschinski, J.; Keim, P. An analysis of genetic variation in Astragalus cremnophylax var. cremnophylax, a critically endangered plant, using AFLP markers. Mol. Ecol. 1996, 5, 735–745. [Google Scholar] [CrossRef]
- Walker, G.F.; Metcalf, A.E. Bulletin of the Southern California Academy of Sciences Genetic Variation in the Endangered Astragalus jaegerianus (Fabaceae, Papilionoideae): A Geographically Restricted Species. Bull. South. Calif. Acad. Sci. 2008, 107, 158–177. [Google Scholar]
- Rogenski, C.; Smith, M.; Esselman, E. ISSR Marker Diversity of the Threatened Astragalus crassicarpus var. trichocalyx in Illinois Populations. Trans. Ill. State Acad. Sci. 2009, 102, 149–159. [Google Scholar]
- Baskauf, C.J.; Burke, J.M. Population genetics of astragalus bibullatus (Fabaceae) using AFLPs. J. Hered. 2009, 100, 424–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Wall, W.A.; Douglas, N.A.; Hoffmann, W.A.; Wentworth, T.R.; Gray, J.B.; Xiang, Q.Y.J.; Knaus, B.K.; Hohmann, M.G. Evidence of population bottleneck in Astragalus michauxii (Fabaceae), a narrow endemic of the southeastern United States. Conserv. Genet. 2014, 15, 153–164. [Google Scholar] [CrossRef]
- Alexander, J.A.; Liston, A.; Popovich, S.J. Genetic diversity of the narrow endemic Astragalus oniciformis (Fabaceae). Am. J. Bot. 2004, 91, 2004–2012. [Google Scholar] [CrossRef] [Green Version]
- Gafta, D.; Mountford, J.O. Manual de interpretare a habitatelor Natura 2000 din România [Romanian Manual for Interpretation of EU Habitats]; Risoprint: Cluj-Napoca, Romania, 2008. [Google Scholar]
- Bădărău, A.S.; Coldea, G.; Groza, G.; Gudasz, C. The biogeography of an endemic element from the Transylvanian Plain, Astragalus exscapus ssp. transsylvanicus. Studia Univ. Babes-Bolyai Geogr. 1999, 44, 45–59. (In Romanian) [Google Scholar]
- Rus, T. Aplicarea legii de reformă agrară din 1921 în aşezările judeţului Mureş. Marisia. 1979, 9, 443–474. [Google Scholar]
- Pascu, Ș. Voievodatul Transilvaniei; Dacia Ed.: Cluj.-Napoca, Romania, 1972; Volume 1. [Google Scholar]
- Pascu, Ș. Voievodatul Transilvaniei; Dacia Ed.: Cluj-Napoca, Romania, 1979; Volume 2. [Google Scholar]
- Feurdean, A.; Marinova, A.; Nielsen, A.B.; Liakka, J.; Veress, D.; Hutchinsons, M.; Braun, M.; Timar-Gabor, A.; Astalos, C.; Mosburgger, V.; et al. Origin of the forest steppe and exceptional grassland diversity in Transylvania (central-eastern Europe). J. Biogeogr. 2015, 42, 951–963. [Google Scholar] [CrossRef] [Green Version]
- Peterman, W.E.; Feist, S.M.; Semlitsch, R.D.; Eggert, L.S. Conservation and management of peripheral populations: Spatial and temporal influences on the genetic structure of wood frog (Rana sylvatica) populations. Biol. Conserv. 2013, 158, 351–358. [Google Scholar] [CrossRef]
- Lesica, P.; Allendorf, F.W. When are Peripheral Populations valuable for Conservation? Conserv. Biol. 1995, 9, 753–760. [Google Scholar] [CrossRef]
- Szabo, K.; Sisea, C.R.; Pop, R.; Bodea, M.; Berindean, I.V.; Badarau, A.S. Pamfil Doru Preliminary Testing of SRAP Primers in Order to Establish Genetic Diversity of Astragalus exscapus L. Bull. Hortic. 2015, 72, 188–190. [Google Scholar] [CrossRef]
- Roldan-Ruiz, I.; Dendauw, J.; van Bockstaele, E.; Depicker, A.; De Loose, M. AFLP markers reveal high polymorphic rates in ryegrasses (Lolium spp.). Mol. Breed. 2000, 6, 125–134. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Yeh, F.C. Population genetic analysis of co-dominant and dominant marker and quantitative traits. Belgian J. Bot. Bot. 1997, 130, 129–157. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; Bridgett, M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Plant Height (mm) | Number of Leaves/Plant | Number of Flowers/Plant | Inflorescence Diameter (mm) | ||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SE | Range | Mean ± SE | Range | Mean ± SE | Range | Mean ± SE | Range | |
| M | 230.9 ± 23.7 | 195–265 | 26.1 ± 18.4 | 9–73 | 44.6 ± 37.7 | 15–143 | 68.8 ± 37.1 | 9–140 |
| B | 158.8 ± 40.5 | 95–250 | 19.5 ± 9.4 | 7–45 | 28.8 ± 17.8 | 4–65 | 53.0 ± 16.8 | 19–76 |
| G | 159.6 ± 44.3 | 84–260 | 21.5 ± 14.4 | 5–52 | 22.3 ± 16.4 | 8–73 | 51.5 ± 21.9 | 22–105 |
| SI | 188.3 ± 49.9 | 93–270 | 17.8 ± 11.5 | 8–52 | 29.2 ± 24.1 | 8–89 | 57.8 ± 21.4 | 23–110 |
| S | 260.6 ± 62.8 | 140–323 | 34.3 ± 21.0 | 12–82 | 76.8 ± 60.0 | 3–227 | 102.4 ± 43.9 | 16–175 |
| V | 211.7 ± 59.3 | 120–320 | 22.5 ± 12.2 | 6–43 | 41.4 ± 16.3 | 12–75 | 89.4 ± 34.8 | 28–147 |
| SG | 163.5 ± 51.0 | 59–255 | 22.2 ± 17.9 | 7–62 | 27.8 ± 14.6 | 13–47 | 82.8 ± 47.6 | 28–134 |
| VF | 298.7 ± 27.1 | 240–332 | 39.5 ± 23.1 | 8–94 | 73.8 ± 52.2 | 13–191 | 100.3 ± 38.6 | 30–175 |
| SRAP Primers | Forward Primer (5′−3′) | Reverse Primer (5′−3′) | NTB | NPB | Size (bp) | PPB (%) | PIC |
|---|---|---|---|---|---|---|---|
| Me1/Em6 | TGAGTCCAAACCGGATA | GACTGCGTACGAATTGCA | 11 | 11 | 167–1107 | 100 | 0.42 |
| Me2/Em1 | TGAGTCCAAACCGGAGC | GACTGCGTACGAATTAAT | 10 | 9 | 187–879 | 90 | 0.28 |
| Me2/Em6 | TGAGTCCAAACCGGAGC | GACTGCGTACGAATTGCA | 9 | 8 | 161–1263 | 88.9 | 0.23 |
| Me3/Em3 | TGAGTCCAAACCGGAAT | GACTGCGTACGAATTGAC | 10 | 10 | 211–1144 | 100 | 0.36 |
| Me4/Em2 | TGAGTCCAAACCGGACC | GACTGCGTACGAATTTGC | 11 | 11 | 218–1013 | 100 | 0.40 |
| Me4/Em4 | TGAGTCCAAACCGGACC | GACTGCGTACGAATTTGA | 12 | 12 | 217–910 | 100 | 0.48 |
| Me4/Em5 | TGAGTCCAAACCGGACC | TGAGTCCAAACCGGATA | 16 | 15 | 236–1733 | 93.75 | 0.25 |
| Me5/Em2 | TGAGTCCAAACCGGAAG | GACTGCGTACGAATTTGC | 10 | 9 | 204–857 | 90.00 | 0.26 |
| Me5/Em6 | TGAGTCCAAACCGGAAG | GACTGCGTACGAATTGCA | 11 | 10 | 182–1068 | 90.90 | 0.25 |
| Me6/Em1 | TGAGTCCAAACCGGACA | GACTGCGTACGAATTAAT | 16 | 16 | 237–1700 | 100 | 0.37 |
| Me6/Em8 | TGAGTCCAAACCGGACA | GACTGCGTACGAATTCAC | 14 | 14 | 210–1548 | 100 | 0.31 |
| Me8/Em2 | TGAGTCCAAACCGGACT | GACTGCGTACGAATTTGC | 10 | 10 | 235–1200 | 100 | 0.36 |
| Me8/Em3 | TGAGTCCAAACCGGACT | GACTGCGTACGAATTGAC | 9 | 9 | 220–1373 | 100 | 0.46 |
| Me1/Em2 | TGAGTCCAAACCGGATA | GACTGCGTACGAATTTGC | 15 | 15 | 175–1411 | 100 | 0.32 |
| Total | 164 | 159 | - | - | - | ||
| Mean | 11.71 | 11.36 | 96.7 | 0.34 |
| Population | Code | N | PBP (%) | Na | Ne | I | He |
|---|---|---|---|---|---|---|---|
| Milǎșel (MS) | M | 14.00 | 75.69% | 1.576 | 1.389 | 0.358 | 0.234 |
| Cojocna-Dl.Beleni (CJ) | B | 14.00 | 72.22% | 1.493 | 1.349 | 0.324 | 0.210 |
| Gǎdǎlin (CJ) | G | 19.00 | 85.41% | 1.743 | 1.403 | 0.382 | 0.247 |
| Silivașu de câmpie (BN) | SI | 15.00 | 85.42% | 1.764 | 1.374 | 0.370 | 0.235 |
| Sǎbed (MS) | S | 15.00 | 72.92% | 1.514 | 1.355 | 0.325 | 0.212 |
| Viișoara (CJ) | V | 18.00 | 78.47% | 1.611 | 1.363 | 0.343 | 0.221 |
| Sǎrmǎșel Garǎ (MS) | SG | 15.00 | 78.47% | 1.611 | 1.390 | 0.363 | 0.236 |
| Valea Florilor (CJ) | VF | 15.00 | 72.92% | 1.528 | 1.373 | 0.349 | 0.227 |
| Average | 15.50 | 77.69% | 1.605 | 1.374 | 0.352 | 0.228 | |
| Species-level | 99.31 | 1.993 | 1.445 | 0.425 | 0.272 |
| Source of Variation | DF | SS | MS | Est. Var. | % | PhiPT |
|---|---|---|---|---|---|---|
| Within populations | 116 | 2259.464 | 19.478 | 19.478 | 83% | |
| Among populations | 7 | 558.584 | 79.798 | 3.900 | 17% | 0.167 ** |
| Total | 123 | 2818.048 | 23.378 | 100% |
| Population | M | B | G | SI | S | V | SG | VF |
|---|---|---|---|---|---|---|---|---|
| M | - | |||||||
| B | 0.159 | - | ||||||
| G | 0.188 | 0.166 | - | |||||
| SI | 0.199 | 0.164 | 0.158 | - | ||||
| S | 0.438 | 0.406 | 0.438 | 0.480 | - | |||
| V | 0.246 | 0.205 | 0.218 | 0.227 | 0.348 | - | ||
| SG | 0.248 | 0.214 | 0.211 | 0.233 | 0.361 | 0.113 | - | |
| VF | 0.302 | 0.264 | 0.292 | 0.291 | 0.449 | 0.285 | 0.243 | - |
| Population. | Code | No. of Samples | Latitude (N) | Longitude (W) | Area (m2) | Census Size |
|---|---|---|---|---|---|---|
| Milǎșel, Mureș County | M | 14 | 46°44′52″ | 24°26′55″ | 654 | 109 |
| Cojocna, Beleni hill, Cluj County | B | 15 | 46°45′57″ | 23°51′52″ | 1148 | 395 |
| Gǎdǎlin, Cluj County | G | 19 | 46°51′28″ | 23°50′50″ | 525 | 68 |
| Silivașu de câmpie, Bistrița Năsăud County | SI | 15 | 46°46′41″ | 24°17′04″ | 5546 | 1109 |
| Săbed, Mureș County | S | 15 | 46°39′52″ | 24°26′46″ | 1205 | 182 |
| Viișoara, Cluj County | V | 18 | 46°33′28″ | 23°47′48″ | 806 | 128 |
| Sǎrmǎșel Garǎ, Mureș County | SG | 15 | 46°47′03″ | 24°12′45″ | 463 | 93 |
| Valea florilor, Cluj County | VF | 15 | 46°40′25″ | 23°54′35″ | 3706 | 335 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabo, K.; Pamfil, D.; Bădărău, A.S.; Hârţa, M. Assessment of Genetic Diversity and Population Structure of the Endangered Astragalus exscapus subsp. transsilvanicus through DNA-Based Molecular Markers. Plants 2021, 10, 2732. https://doi.org/10.3390/plants10122732
Szabo K, Pamfil D, Bădărău AS, Hârţa M. Assessment of Genetic Diversity and Population Structure of the Endangered Astragalus exscapus subsp. transsilvanicus through DNA-Based Molecular Markers. Plants. 2021; 10(12):2732. https://doi.org/10.3390/plants10122732
Chicago/Turabian StyleSzabo, Katalin, Doru Pamfil, Alexandru Sabin Bădărău, and Monica Hârţa. 2021. "Assessment of Genetic Diversity and Population Structure of the Endangered Astragalus exscapus subsp. transsilvanicus through DNA-Based Molecular Markers" Plants 10, no. 12: 2732. https://doi.org/10.3390/plants10122732