
Silicon Mitigates Negative Impacts of Drought and UV-B Radiation in Plants


Abstract
:1. Introduction
2. Silicon
3. Drought
3.1. Plant Damage by Drought
3.2. Plant Protection from Drought
3.3. Drought and Silicon

{kind=link}
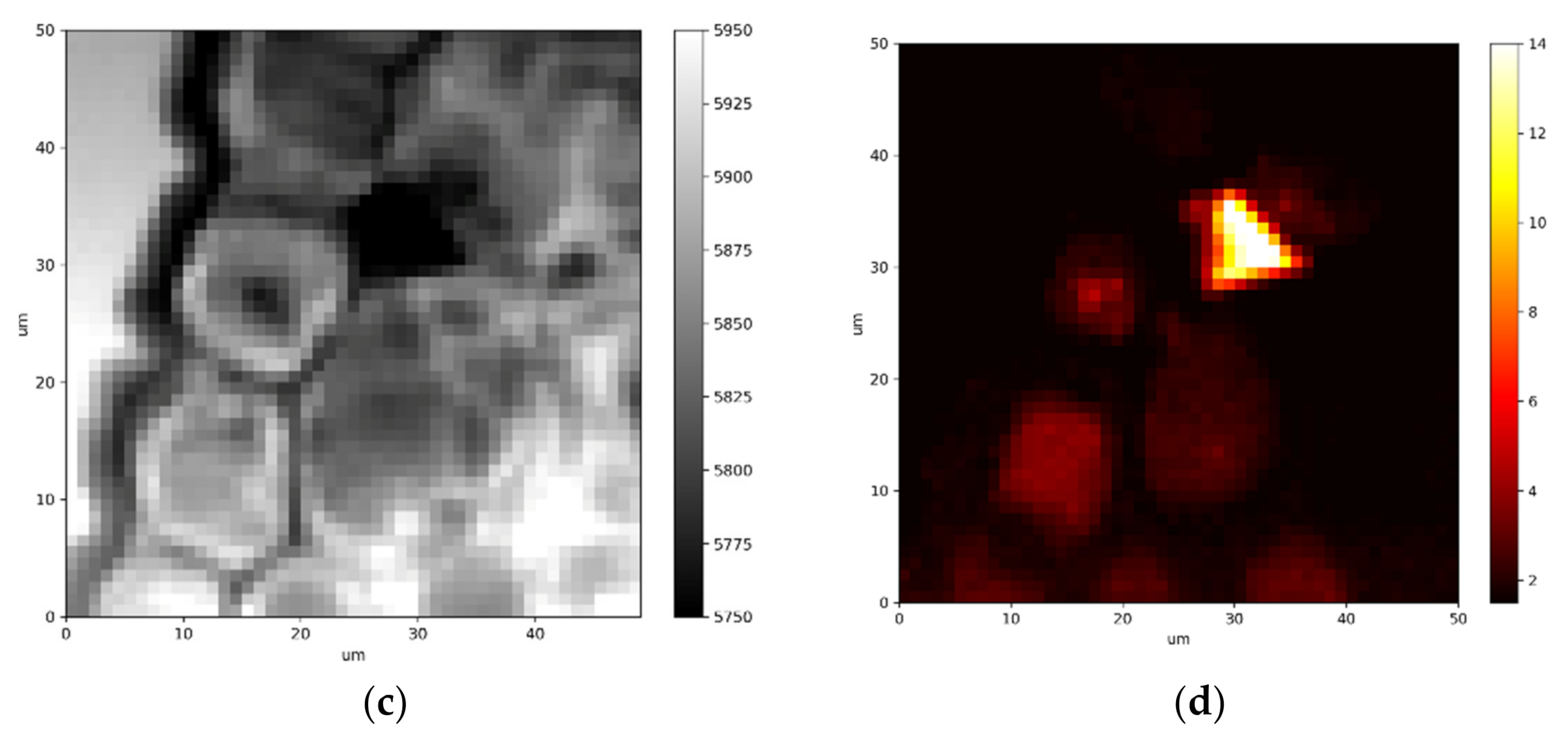{kind=link}
| Plant Species | Silicon Treatment | Drought Duration | Results (Si Treated vs. Not Si Treated) | Reference | |
|---|---|---|---|---|---|
| Source | Concentration | ||||
| Rice (Oryza sativa L.) | Calcium and magnesium silicate | 0, 350 kg Si ha−1 | - | Decreased proline content | Mauad et al., 2016 [47] |
| Sugarcane (Saccharum officinarum L.) | Calcium magnesium silicate | 0, 600 kg Si ha−1 | 30/60 days | Increased ascorbate acid peroxidase (30 days), proline (30 days); decreased malondialdehyde | Bezerra et al., 2019 [44] |
| Wheat (Triticum aestivum L.) | Potassium silicate | 0, 2, 4, 6 mM Si | 45 days | Increased total soluble sugars, proline, ascorbic acid, glutathione, net photosynthetic rate, plant growth, relative water content; decreased malondialdehyde content | Alzahrani et al., 2018 [46] |
| Sodium silicate | 1.5 mM Si/0.2, 0.9, 1.8 mM Si | 6 weeks (soil)/4 weeks (hydroponics) | Increased Si concentration | Thorne et al., 2021 [53] | |
| Maize (Zea mays L.) | Sodium silicate | 0, 2, 4 mM Si | Two summer seasons | Increased growth, number of leaves per plant, photosynthesis efficiency, stomatal conductance, cell membrane integrity; decreased accumulation of Ni2+, Cd2+, Cr3+ | Abd El-Mageed et al., 2020 [51] |
| Kentucky bluegrass (Poa pratensis L.) | Sodium silicate | 0, 200, 400, and 80 0 mg Si L−1 | 20 days | Decreased the leaf C/N ratio; increased photosynthesis, leaf water content, relative growth rate, leaf green color, root/shoot ratio, and turf quality | Chen et al., 2014 [49] |
| Tomato (Solanum lycopersicum L.) | Potassium silicate | 2.5 mM Si | 7 days | Increased growth, chlorophyll and carotenoid content, maximum photochemical efficiency, effective quantum efficiency, actual photochemical quantum efficiency, photochemical quenching coefficient, and electron transport rate; up-regulation of genes related to photosynthesis | Zhang et al., 2018 [48] |
| Sodium silicate | 0.6 mM Si | 12 days | Increased net photosynthetic rate; decreased reactive oxygen species | Cao et al., 2020 [45] | |
| 0.6, 1.2, 1.8 mM Si | 12 days | Increased maximum photochemical efficiency, electron transport rate, net photosynthetic rate | Cao et al., 2015 [50] | ||
| Mango (Mangifera indica L.) | Potassium silicate | 1.5 mM Si | - | Increased indole-3-acetic acid, gibberellic acid, cytokinin levels, relative growth rate, net assimilation rate, relative water content, chlorophyll and carotenoids contents; decreased abscisic acid levels, and peroxidase, catalase, superoxide dismutase activities | Helaly et al., 2017 [6] |
| Cantaloupe (Cucumis melo var. reticulatus L. Naud. cv. Galia) | Silicic acid | 0, 100, 200 and 400 kg Si ha−1 | 30 days | Increasing growth and number of leaves per plant, fruit length, diameter, flesh thickness, and overall fruit yield | Alam et al., 2020 [52] |
| Oilseed rape (Brassica napus L.) | Orthosilicic acid tetraethyl ester | 3.4 mM Si | 10 days | Improved water uptake | Saja-Garbarz et al., 2021 [54] |
4. Ultraviolet Radiation
4.1. Plant Damage by UV-B Radiation
4.2. Plant Protection from UV Radiation
4.3. UV-B and Silicon
| Plant Species | Silicon Treatment | UV Treatment | Results (Si Treated vs. Not Si Treated) | Reference | ||
|---|---|---|---|---|---|---|
| Source | Concentration | Level | Duration | |||
| Wheat (Triticum aestivum L.) | Potassium silicate | 0, 100, 200, 400 mg SiO2/kg substrate | 25%/50% ozone depletion | - | Increased antioxidant compound contents, Si concentration in leaves, total biomass, and contents of chlorophyll (a + b), soluble sugars, anthocyanins, flavonoids; reduced superoxide radical (O2−) production and malondialdehyde content | Yao et al., 2011 [3] |
| Si, Si nanoparticles | 10 µM Si/Si nanoparticles | Ambient and enhanced UV | 2 days, 6/8 h/day | Increased fresh and dry mass | Tripathi et al., 2017 [84] | |
| Rice (Oryza sativa L.) | Silicic acid | 1.5 mM Si | UV-B, 10 W | 30 h | Increased soluble and insoluble UV-absorbing compounds in epidermis; brown spots in plants without Si | Li et al., 2004 [87] |
| Sodium silicate | 0/5 mM Si | UV-B, 10.27 W m−2 | 15/30 min | Decreased reactive oxygen species, thiobarbituric acid reactive substances, total phenols and flavonoid content | Mihaličova Malčovska et al., 2014 [5] | |
| Soybean (Glycine max L.) | Sodium silicate | 1.7 mM Si | 30% ozone depletion | 1 week, 8 h/day | Increased P and Mg uptake and dry mass (in one cultivar) | Shen et al., 2009 [86] |
| 1 week | Increase in net photosynthetic rate; decrease in proline content, lipid peroxidation, osmolyte leakage | Shen et al., 2010 [4] | ||||
| 1.7/2.55 mM Si | 15%, 30% ozone depletion | 1 week, 8 h/day | Decreased catalase, superoxide dismutase and peroxidase activities | Shen et al., 2010 [85] | ||
5. Silicon, UV-B, and Drought
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kramer, G.F.; Norman, H.A.; Krizek, D.T.; Mirecki, R.M. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Tevini, M. UV-Effects on Plants. In Concepts in Photobiology; Springer: Dordrecht, The Netherlands, 1999; pp. 588–613. [Google Scholar] [CrossRef]
- Yao, X.; Chu, J.; Cai, K.; Liu, L.; Shi, J.; Geng, W. Silicon improves the tolerance of wheat seedlings to Ultraviolet-B stress. Biol. Trace Elem. Res. 2011, 143, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zhou, Y.; Duan, L.; Li, Z.; Eneji, A.E.; Li, J. Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and Ultraviolet-B radiation. J. Plant Physiol. 2010, 167, 1248–1252. [Google Scholar] [CrossRef]
- Mihaličová Malčovská, S.; Dučaiová, Z.; Bačkor, M. Impact of silicon on maize seedlings exposed to short-term UV-B irradiation. Biologia 2014, 69, 1349–1355. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hoseiny, H.; El-Sheery, N.I.; Rastogi, A.; Kalaji, H.M. Regulation and physiological role of silicon in alleviating drought stress of mango. Plant Physiol. Biochem. 2017, 118, 31–44. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Kapoor, D. Fascinating regulatory mechanism of silicon for alleviating drought stress in plants. Plant Physiol. Biochem. 2021, 166, 1044–1053. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Mur, L.A.J.; Ruan, J.; Shen, Q.; Guo, S. Functions of silicon in plant drought stress responses. Hortic. Res. 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Rehman, M.U. Application of silicon: A useful way to mitigate drought stress: An overview. Curr. Res. Agric. Farming 2021, 2, 9–17. [Google Scholar] [CrossRef]
- Hans Wedepohl, K. The composition of the continental crust. Geochim. Cosmochim. Acta 1995, 59, 1217–1232. [Google Scholar] [CrossRef]
- Matichenkov, V.V.; Bocharnikova, E.A. The relationship between silicon and soil physical and chemical properties. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 209–219. [Google Scholar] [CrossRef]
- Tubana, B.S.; Babu, T.; Datnoff, L.E. A review of silicon in soils and plants and its role in US Agriculture. Soil Sci. 2016, 181, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Gaur, S.; Kumar, J.; Kumar, D.; Chauhan, D.K.; Prasad, S.M.; Srivastava, P.K. Fascinating impact of silicon and silicon transporters in plants: A review. Ecotoxicol. Environ. Saf. 2020, 202, 110885. [Google Scholar] [CrossRef]
- Chen, C.; Lewin, J. Silicon as a nutrient element for Equisetum arvense. Can. J. Bot. 1969, 47, 125–131. [Google Scholar] [CrossRef]
- Ma, J.F.; Takahashi, E. Silicon-accumulating plants in the plant kingdom. In Soil, Fertilizer, and Plant Silicon Research in Japan; Ma, J.F., Takahashi, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 63–71. [Google Scholar] [CrossRef]
- Klančnik, K.; Vogel-Mikuš, K.; Kelemen, M.; Vavpetič, P.; Pelicon, P.; Kump, P.; Jezeršek, D.; Gianoncelli, A.; Gaberščik, A. Leaf optical properties are affected by the location and type of deposited biominerals. J. Photochem. Photobiol. B Biol. 2014, 140, 276–285. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T.; Vaculík, M. Silicon influences soil availability and accumulation of mineral nutrients in various plant species. Plants 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kump, P.; Vogel-Mikuš, K. Quantification of 2D elemental distribution maps of intermediate-thick biological sections by low energy synchrotron μ-X-Ray fluorescence spectrometry. J. Instrum. 2018, 13, C05014. [Google Scholar] [CrossRef]
- Vaculík, M.; Lukačová, Z.; Bokor, B.; Martinka, M.; Tripathi, D.K.; Lux, A. Alleviation mechanisms of metal(loid) stress in plants by silicon: A review. J. Exp. Bot. 2020, 71, 6744–6757. [Google Scholar] [CrossRef]
- Zargar, S.M.; Mahajan, R.; Bhat, J.A.; Nazir, M.; Deshmukh, R. Role of silicon in plant stress tolerance: Opportunities to achieve a sustainable cropping system. 3 Biotech. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S.; Maatougui, M.; Michael, M.; Slash, M.; Haghparast, R.; Rahmanian, M.; Taheri, A.; Al-Yassin, A.; Benbelkacem, A.; et al. Plant breeding and climate changes. J. Agric. Sci. 2010, 148, 627–637. [Google Scholar] [CrossRef]
- Karl, T.R.; Melillo, J.M.; Peterson, T.C. Global Climate Change Impacts in the United States; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global synthesis of drought effects on food legume production. PLoS ONE 2015, 10, e0127401. [Google Scholar] [CrossRef] [Green Version]
- Sorkheh, K.; Shiran, B.; Rouhi, V.; Khodambashi, M.; Sofo, A. Regulation of the ascorbate-glutathione cycle in wild almond during drought stress. Russ. J. Plant Physiol. 2011, 58, 76–84. [Google Scholar] [CrossRef]
- Colom, M.R.; Vazzana, C. Photosynthesis and PSII functionality of drought-resistant and drought-sensitive weeping lovegrass plants. Environ. Exp. Bot. 2003, 49, 135–144. [Google Scholar] [CrossRef]
- Parry, M.A.J.; Andralojc, P.J.; Khan, S.; Lea, P.J.; Keys, A.J. Rubisco activity: Effects of drought stress. Ann. Bot. 2002, 89, 833–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Terzi, R.; Saǧlam, A.; Kutlu, N.; Nar, H.; Kadioǧlu, A. Impact of soil drought stress on photochemical efficiency of photosystem II and antioxidant enzyme activities of Phaseolus vulgaris cultivars. Turk. J. Botany 2010, 34, 1–10. [Google Scholar] [CrossRef]
- Nelissen, H.; Sun, X.H.; Rymen, B.; Jikumaru, Y.; Kojima, M.; Takebayashi, Y.; Abbeloos, R.; Demuynck, K.; Storme, V.; Vuylsteke, M.; et al. The reduction in maize leaf growth under mild drought affects the transition between cell division and cell expansion and cannot be restored by elevated gibberellic acid levels. Plant Biotechnol. J. 2018, 16, 615–627. [Google Scholar] [CrossRef] [Green Version]
- Alkahtani, M.D.F.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Al Husnain, L.; Algwaiz, H.I.M.; Abdelaal, K.A.A. Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef]
- Mulroy, T.W.; Rundel, P.W. Annual plants: Adaptations to desert environments. Bioscience 1977, 27, 109–114. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Saed-Moucheshi, A.; Pakniyat, H.; Pessarakli, M. Stomatal responses to drought stress. In Water Stress and Crop Plants; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 24–40. [Google Scholar] [CrossRef]
- Shepherd, T.; Griffiths, D.W. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, F.J.; Manzanares, M.; De Andres, E.F.; Tenorio, J.L.; Ayerbe, L. Turgor maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in response to water stress. Field Crop. Res. 1998, 59, 225–235. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.S.; Zou, Y.N. Arbuscular mycorrhizal fungi and tolerance of drought stress in plants. In Arbuscular Mycorrhizas Stress Tolerance of Plants; Springer Nature: Singapore, 2017; pp. 25–41. [Google Scholar] [CrossRef]
- Sasidharan, R.; Voesenek, L.A.C.J.; Pierik, R. Cell wall modifying proteins mediate plant acclimatization to biotic and abiotic stresses. CRC. Crit. Rev. Plant Sci. 2011, 30, 548–562. [Google Scholar] [CrossRef]
- Lü, P.; Kang, M.; Jiang, X.; Dai, F.; Gao, J.; Zhang, C. RhEXPA4, a rose expansin gene, modulates leaf growth and confers drought and salt tolerance to Arabidopsis. Planta 2013, 237, 1547–1559. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Cruz De Carvalho, M.H.; Torres-Jerez, I.; Kang, Y.; Allen, S.N.; Huhman, D.V.; Tang, Y.; Murray, J.; Sumner, L.W.; Udvardi, M.K. Global reprogramming of transcription and metabolism in Medicago truncatula during progressive drought and after rewatering. Plant Cell Environ. 2014, 37, 2553–2576. [Google Scholar] [CrossRef] [Green Version]
- Verma, G.; Srivastava, D.; Tiwari, P.; Chakrabarty, D. ROS modulation in crop plants under drought stress ROS generation: An overview. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; John Wiley & Sons: Hoboken, NJ, USA, 2019; Volume 1, pp. 311–336. [Google Scholar]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, B.K.L.; Lima, G.P.P.; dos Reis, A.R.; de Almeida Silva, M.; de Camargo, M.S. Physiological and biochemical impacts of silicon against water deficit in sugarcane. Acta Physiol. Plant. 2019, 41, 1–12. [Google Scholar] [CrossRef]
- Cao, B.L.; Ma, Q.; Xu, K. Silicon restrains drought-induced ROS accumulation by promoting energy dissipation in leaves of tomato. Protoplasma 2020, 257, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, Y.; Kuşvuran, A.; Alharby, H.F.; Kuşvuran, S.; Rady, M.M. The defensive role of silicon in wheat against stress conditions induced by drought, salinity or cadmium. Ecotoxicol. Environ. Saf. 2018, 154, 187–196. [Google Scholar] [CrossRef]
- Mauad, M.; Costa Crusciol, C.A.; Nascente, A.S.; Filho, H.G.; Pereira Lima, G.P. Effects of silicon and drought stress on biochemical characteristics of leaves of upland rice cultivars1. Rev. Cienc. Agron. 2016, 47, 532–539. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shi, Y.; Gong, H.; Zhao, H.; Li, H.; Hu, Y.; Wang, Y. Beneficial effects of silicon on photosynthesis of tomato seedlings under water stress. J. Integr. Agric. 2018, 17, 2151–2159. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A. Silicon application increases drought tolerance of kentucky bluegrass by improving plant water relations and morphophysiological functions. Sci. World J. 2014, 2014, 368694. [Google Scholar] [CrossRef]
- Cao, B.; Ma, Q.; Zhao, Q.; Wang, L.; Xu, K. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Sci. Hortic. 2015, 194, 53–62. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Shaaban, A.; Abd El-Mageed, S.A.; Semida, W.M.; Rady, M.O.A. Silicon defensive role in maize (Zea mays L.) against drought stress and metals-contaminated irrigation water. Silicon 2020, 13, 2165–2176. [Google Scholar] [CrossRef]
- Alam, A.; Hariyanto, B.; Ullah, H.; Salin, K.R.; Datta, A. Effects of silicon on growth, yield and fruit quality of cantaloupe under drought stress. Silicon 2020, 13, 3153–3162. [Google Scholar] [CrossRef]
- Thorne, S.J.; Hartley, S.E.; Maathuis, F.J.M. The effect of silicon on osmotic and drought stress tolerance in wheat landraces. Plants 2021, 10, 814. [Google Scholar] [CrossRef] [PubMed]
- Saja-Garbarz, D.; Ostrowska, A.; Kaczanowska, K.; Janowiak, F. Accumulation of silicon and changes in water balance under drought stress in Brassica napus var. napus L. Plants 2021, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Carruthers, T.J.B.; Longstaff, B.J.; Dennison, W.C.; Abal, E.G.; Aioi, K. Measurement of light penetration in relation to seagrass. In Global Seagrass Research Methods; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 369–392. [Google Scholar] [CrossRef]
- McCree, K.J. Test of current definitions of photosynthetically active radiation against leaf photosynthesis data. Agric. Meteorol. 1972, 10, 443–453. [Google Scholar] [CrossRef]
- International Organization for Standardization. ISO 21348 Definitions of solar irradiance spectral categories. Environment 2007, 5, 6–7. [Google Scholar]
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- Kataria, S.; Guruprasad, K.N.; Ahuja, S.; Singh, B. Enhancement of growth, photosynthetic performance and yield by exclusion of ambient UV components in C3 and C4 plants. J. Photochem. Photobiol. B Biol. 2013, 127, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Taylor, J.S. In vivo evidence that UV-induced C → T mutations at dipyrimidine sites could result from the replicative bypass of cis-syn cyclobutane dimers or their deamination products. Biochemistry 1993, 32, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Hoerner, R.; Weissenböck, G. Contribution of phenolic compounds to the UV-B screening capacity of developing barley primary leaves in relation to DNA damage and repair under elevated UV-B levels. Phytochemistry 2003, 64, 243–255. [Google Scholar] [CrossRef]
- Staxén, I.; Bergounioux, C.; Bornman, J.F. Effect of ultraviolet radiation on cell division and microtubule organization in Petunia hybrida protoplasts. Protoplasma 1993, 173, 70–76. [Google Scholar] [CrossRef]
- Grossweiner, L.I. Photochemistry of proteins: A review. Curr. Eye Res. 1984, 3, 137–144. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Marwood, C.A.; Greenberg, B.M. Effect of supplementary UVB Radiation on chlorophyll synthesis and accumulation of photosystems during chloroplast development in Spirodela oligorrhiza. Photochem. Photobiol. 1996, 64, 664–670. [Google Scholar] [CrossRef]
- Correia, C.M.; Pereira, J.M.M.; Coutinho, J.F.; Björn, L.O.; Torres-Pereira, J.M.G. Ultraviolet-B radiation and nitrogen affect the photosynthesis of maize: A mediterranean field study. Eur. J. Agron. 2005, 22, 337–347. [Google Scholar] [CrossRef]
- Kataria, S.; Guruprasad, K.N. Exclusion of solar UV components improves growth and performance of Amaranthus tricolor varieties. Sci. Hortic. 2014, 174, 36–45. [Google Scholar] [CrossRef]
- Pontin, M.A.; Piccoli, P.N.; Francisco, R.; Bottini, R.; Martinez-Zapater, J.M.; Lijavetzky, D. Transcriptome Changes in Grapevine (Vitis vinifera L.) Cv. malbec leaves induced by Ultraviolet-B radiation. BMC Plant Biol. 2010, 10, 224. [Google Scholar] [CrossRef] [Green Version]
- Ros, J.; Tevini, M. Interaction of UV-radiation and IAA during growth of seedlings and hypocotyl segments of sunflower. J. Plant Physiol. 1995, 146, 295–302. [Google Scholar] [CrossRef]
- Klem, K.; Holub, P.; Štroch, M.; Nezval, J.; Špunda, V.; Tříska, J.; Jansen, M.A.K.; Robson, T.M.; Urban, O. Ultraviolet and photosynthetically active radiation can both induce photoprotective capacity allowing barley to overcome high radiation stress. Plant Physiol. Biochem. 2015, 93, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Yang, Y.; Li, Y.; Lutts, S. Intraspecific responses of fagopyrum esculentum to enhanced ultraviolet B radiation. Plant Growth Regul. 2008, 56, 297–306. [Google Scholar] [CrossRef]
- Bornman, J.F.; Evert, R.F.; Mierzwa, R.J. The effect of UV-B and UV-C radiation on sugar beet leaves. Protoplasma 1983, 117, 7–16. [Google Scholar] [CrossRef]
- Bornman, J.F.; Vogelmann, T.C. Effect of UV-B radiation on leaf optical properties measured with fibre optics. J. Exp. Bot. 1991, 42, 547–554. [Google Scholar] [CrossRef]
- Skaltsa, H.; Verykokidou, E.; Harvala, C.; Karabourniotis, G.; Manetasi, Y. UV-B protective potential and flavonoid content of leaf hairs of Quercus ilex. Phytochemistry 1994, 37, 987–990. [Google Scholar] [CrossRef]
- Day, T.A. Relating UV-B radiation screening effectiveness of foliage to absorbing-compound concentration and anatomical characteristics in a diverse group of plants. Oecologia 1993, 95, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Manetas, Y. The importance of being hairy: The adverse effects of hair removal on stem photosynthesis of Verbascum speciosum are due to solar UV-B radiation. New Phytol. 2003, 158, 503–508. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. Northeast Forestry University February 2020. J. For. Res. 2020, 31, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Lelandais, M.; Kunert, K.J. Photooxidative stress in plants. Physiol. Plant. 1994, 92, 696–717. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Scandalios, J.G. Oxygen stress and superoxide dismutases. Plant Physiol. 1983, 101, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Schaller, J.; Brackhage, C.; Dudel, E.G. Silicon availability changes structural carbon ratio and phenol content of grasses. Environ. Exp. Bot. 2012, 77, 283–287. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef]
- Shen, X.; Li, X.; Li, Z.; Li, J.; Duan, L.; Eneji, A.E. Growth, physiological attributes and antioxidant enzyme activities in soybean seedlings treated with or without silicon under UV-B radiation stress. J. Agron. Crop Sci. 2010, 196, 431–439. [Google Scholar] [CrossRef]
- Shen, X.; Li, J.; Duan, L.; Li, Z.; Eneji, A.E. Nutrient acquisition by soybean treated with and without silicon under Ultraviolet-B radiation. J. Plant Nutr. 2009, 32, 1731–1743. [Google Scholar] [CrossRef]
- Li, W.B.; Shi, X.H.; Wang, H.; Zhang, F.S. Effects of silicon on rice leaves resistance to Ultraviolet-B. Acta Bot. Sin. 2004, 46, 691–697. [Google Scholar]
- Grašič, M.; Malovrh, U.; Golob, A.; Vogel-Mikuš, K.; Gaberščik, A. Effects of water availability and UV radiation on silicon accumulation in the C4 crop proso millet. Photochem. Photobiol. Sci. 2019, 18, 375–386. [Google Scholar] [CrossRef]


| Drought | UV-B | UV-B × Drought | Reference | |||
|---|---|---|---|---|---|---|
| Elemental composition | Nutrient uptake |  | [43] | |||
| P, Mg uptake | | [86] | ||||
| Leaf Si concentration | | [3] | ||||
| Leaf C:N ratio |  | [49] | ||||
| Antioxidant capacity | | [3] | ||||
| Antioxidants | Non-enzymatic system | Flavonoids | | [3](↑), [5](↓) | ||
| Anthocyanin | | [3] | ||||
| Ascorbic acid | | [46] | ||||
| Glutathione | | [46] | ||||
| Proline | | | | [44,46,47] 1[4] [4] | ||
| Phenol substances | | [5] | ||||
| Carotenoids | | [6,48] | ||||
| Soluble sugars | | [3] | ||||
| Enzymatic system | Catalase | | | | [6] [85] [4] | |
| Superoxide dismutase | | | | [6] [85] [4] | ||
| Peroxidase | | | [47] [85] | |||
| Ascorbate peroxidase | | [44] | ||||
| Membrane damage | Malondialdehyde | | | | [44,46] [3,4] [4] | |
| Osmolyte leakage | | | [4] [4] | |||
| Hormones | Indole-3-acetic acid | | [6] | |||
| Gibberellic acid | | [6] | ||||
| Cytokinins | | [6] | ||||
| Abscisic acid | | [6] | ||||
| Oxidative stress | Reactive oxygen species | | | [45] [5] | ||
| H2O2 | | [4] | ||||
| Photosynthesis | Photosynthesis rate | | | [45,46,49,50,51] [4] | ||
| Chlorophyll content | | | [6,48] [3] | |||
| Photosynthesis-related genes expression | | [48] | ||||
| Morphology | Growth rate | | [6,46,48,49,52] | |||
| Number of leaves | | [46,52] | ||||
| Total biomass | | [3,84] | ||||
| Root:shoot ratio | | [49] | ||||
| Turf quality | | [49] | ||||
| Water content | Relative water content | | [6,46] | |||
| Leaf water content | | [49] | ||||
| Cell structure | Lignification, suberinisation | | [84] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavrič Čermelj, A.; Golob, A.; Vogel-Mikuš, K.; Germ, M. Silicon Mitigates Negative Impacts of Drought and UV-B Radiation in Plants. Plants 2022, 11, 91. https://doi.org/10.3390/plants11010091
Mavrič Čermelj A, Golob A, Vogel-Mikuš K, Germ M. Silicon Mitigates Negative Impacts of Drought and UV-B Radiation in Plants. Plants. 2022; 11(1):91. https://doi.org/10.3390/plants11010091
Chicago/Turabian StyleMavrič Čermelj, Anja, Aleksandra Golob, Katarina Vogel-Mikuš, and Mateja Germ. 2022. "Silicon Mitigates Negative Impacts of Drought and UV-B Radiation in Plants" Plants 11, no. 1: 91. https://doi.org/10.3390/plants11010091