
The Potential of Edible and Medicinal Resource Polysaccharides for Prevention and Treatment of Neurodegenerative Diseases
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Potential of EMPs for Regulating Neurodegenerative Diseases
2.1. Behavioural Movement Enhancement
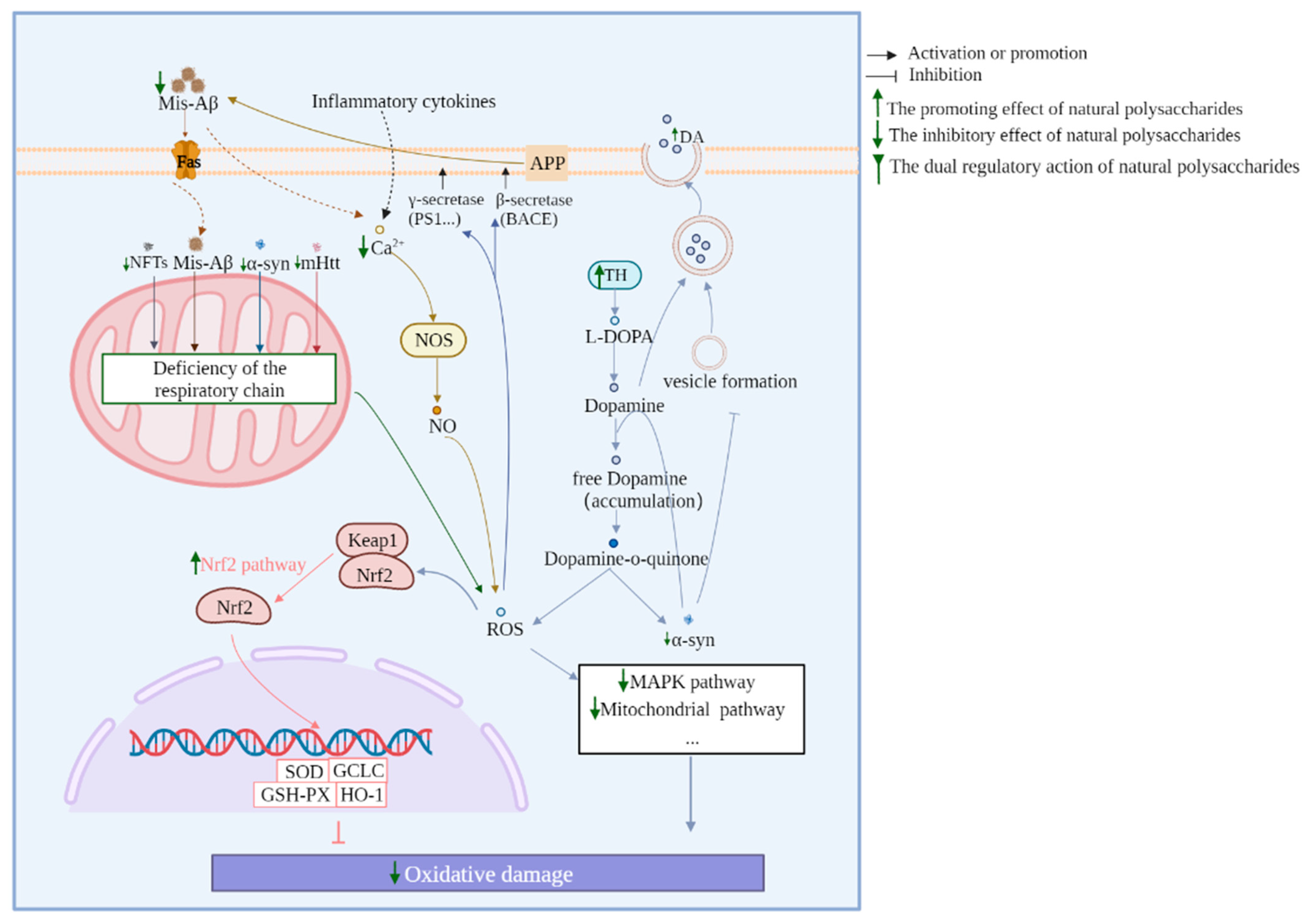
2.2. Reduction in Abnormal Protein Accumulation
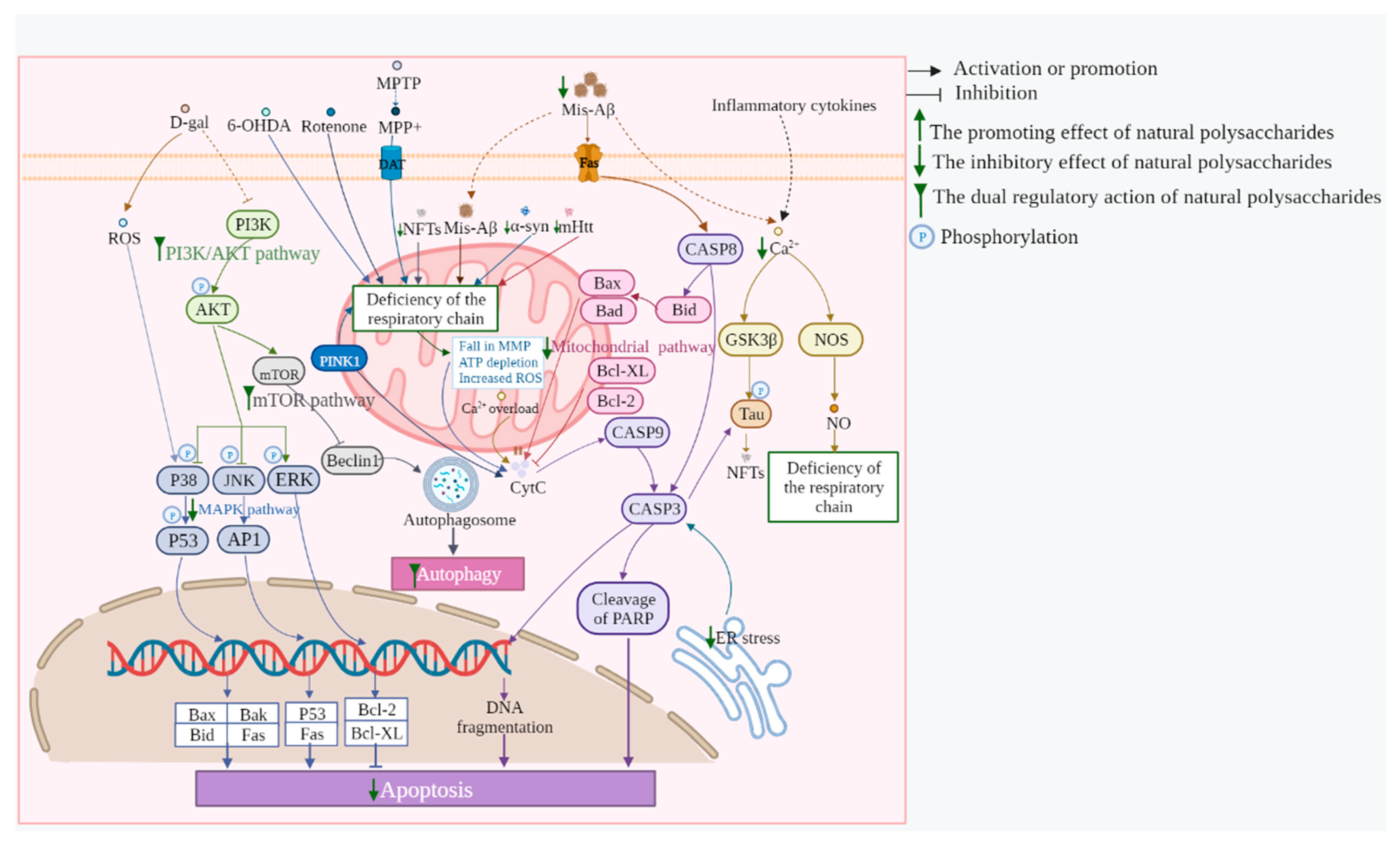
2.3. Neuronal Apoptosis Inhibition
2.4. Reduction in Neuronal Oxidative Stress
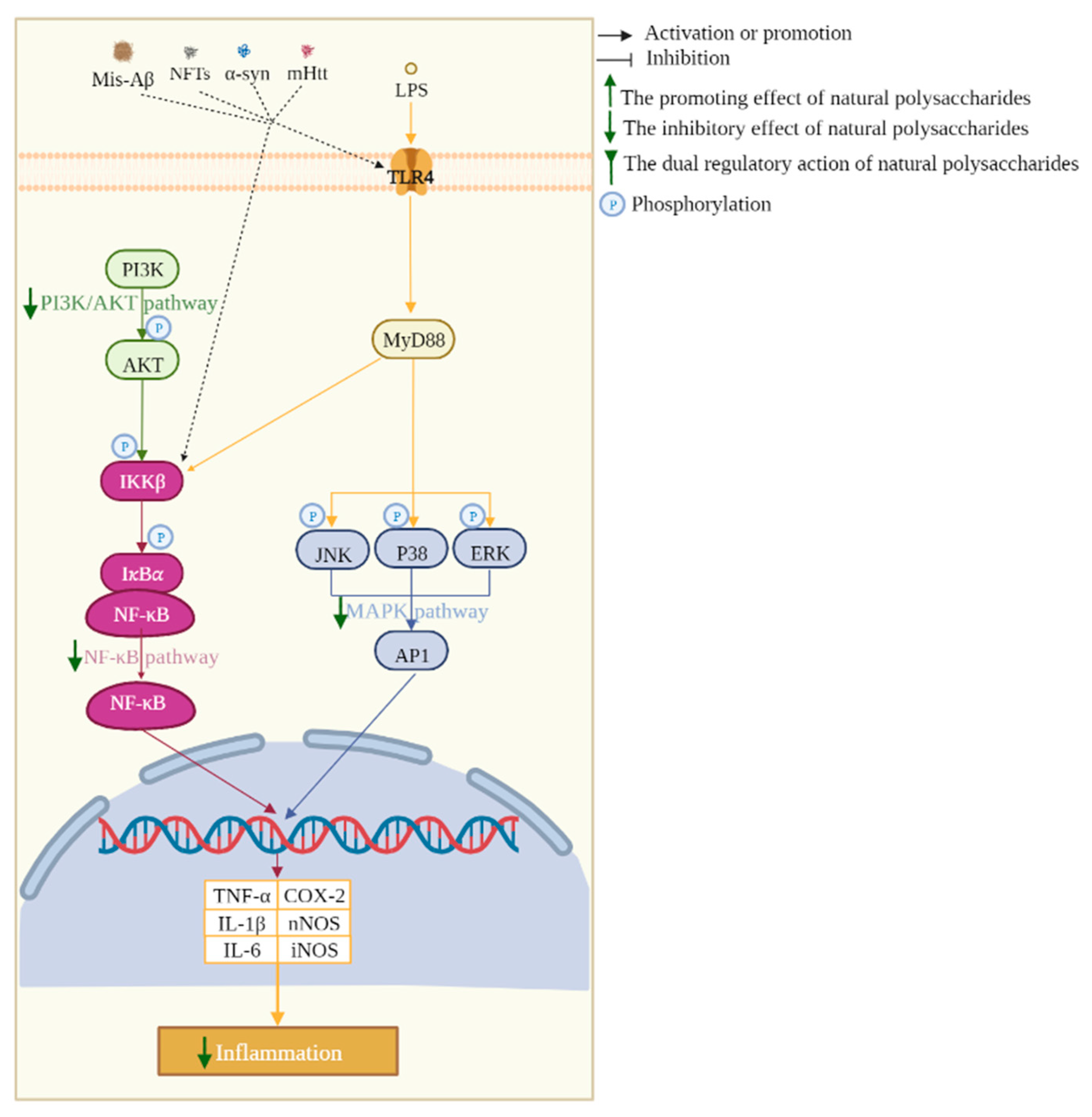
2.5. Neuroinflammation Inhibition
2.6. Gut Microbiota Regulation
2.7. Autophagy Regulation
2.8. Regulation of the Balance of Neurotransmitters
2.9. Restoration of Synaptic Plasticity
3. Multiple Pathways Support the Regulation of Neurodegenerative Diseases with MEPs
3.1. Mitochondrial Pathways
3.2. MAPK Pathway
3.3. NF-κB Pathway
3.4. Nrf2 Pathway
3.5. mTOR Pathway
3.6. PI3K/AKT Pathway
3.7. P53/p21 Pathway
3.8. BDNF/TrkB/CREB Pathway
4. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NFTs | neurofibrillary tangles |
| AKT | serine/threonine protein kinase B |
| MAPK | mitogen-activated protein kinase |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| BDNF | brain-derived neurotrophic factor |
| TrkB | neurotrophic tyrosine kinase receptor type 2 |
| CREB | cyclic AMP-responsive element-binding protein |
| AD | Alzheimer’s disease |
| Gal | galactose |
| PS1 | presenilin 1 |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| MPP+ | 1-methyl-4-phenylpyridine |
| 6-OHDA | 6-Hydroxydopamine hydrobromide |
| PD | Parkinson’s disease |
| HTT | Huntington protein |
| BBB | Blood–brain barrier |
| DA | dopamine |
| DAT | dopamine transporter |
| SN | substantia nigra |
| HD | Huntington’s disease |
| PolyQ | polyglutamic acid |
| LPS | lipopolysaccharide |
| MMW | morris water maze |
| OFT | open field test |
| NOR | novel object recognition test |
| α-syn | α-synuclein |
| TUNEL | TdT-mediated dUTP Nick-End Labeling |
| CAT | catalase |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| MDA | malondialdehyde |
| GSH-Px | glutathione peroxidase |
| AchE | acetylcholinesterase |
| Ach | acetylcholine |
| ChAT | choline acetyltransferase |
| 3-NT | 3-nitrotyrosine |
| 8-OHdG | 8-hydroxy-2-deoxyguanosine |
| IBA-1 | ionized calcium binding adaptor molecule-1 |
| IL | interleukin |
| TNF | tumor necrosis factor |
| iNOS | inducible nitric oxide synthase |
| COX-2 | cyclo-oxyganese-2 |
| NLRP3 | NOD-like receptor thermal protein domain associated protein 3 |
| 5-HT | 5-hydroxytryptamine |
| ERK | extracellular signal-regulated kinase |
| GSK-3 | glycogen synthase kinase-3 |
| PGE2 | prostaglandinE2 |
| TLR4 | Toll-like receptor 4 |
| MyD88 | myeloid differentiation primary response protein |
| JNK | c-Jun N-terminal kinase |
| PGC-1α | peroxisome proliferator-activated receptor-γ coactlvator-1α |
References
- Santos, A.C.D.; Nogueira, M.L.; de Oliveira, F.P.; Costa, E.V.; Bezerra, D.P. Essential Oils of Duguetia Species A. St. Hill (Annonaceae): Chemical Diversity and Pharmacological Potential. Biomolecules 2022, 12, 615. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Lao, F.; Pan, X.; Wu, J.H. Food Protein-Derived Antioxidant Peptides: Molecular Mechanism, Stability and Bioavailability. Biomolecules 2022, 12, 622. [Google Scholar] [CrossRef] [PubMed]
- Klose, J.; Griehl, C.; Rossner, S.; Schilling, S. Natural Products from Plants and Algae for Treatment of Alzheimer’s Disease: A Review. Biomolecules 2022, 12, 694. [Google Scholar] [CrossRef]
- Karagianni, K.; Pettas, S.; Christoforidou, G.; Kanata, E.; Bekas, N.; Xanthopoulos, K.; Dafou, D.; Sklaviadis, T. A Systematic Review of Common and Brain-Disease-Specific RNA Editing Alterations Providing Novel Insights into Neurological and Neurodegenerative Disease Manifestations. Biomolecules 2022, 12, 465. [Google Scholar] [CrossRef]
- Farmen, K.; Tofino-Vian, M.; Iovino, F. Neuronal Damage and Neuroinflammation, a Bridge Between Bacterial Meningitis and Neurodegenerative Diseases. Front. Cell. Neurosci. 2021, 15, 680858. [Google Scholar] [CrossRef] [PubMed]
- Doggrell, S. Still grasping at straws: Donanemab in Alzheimer’s disease. J Expert Opin. Investig. Drugs 2021, 30, 797–801. [Google Scholar] [CrossRef]
- Calzaferri, F.; Ruiz-Ruiz, C.; de Diego, A.; de Pascual, R.; Méndez-López, I.; Cano-Abad, M.; Maneu, V.; de Los Ríos, C.; Gandía, L.; García, A. The purinergic P2X7 receptor as a potential drug target to combat neuroinflammation in neurodegenerative diseases. J. Med. Res. Rev. 2020, 40, 2427–2465. [Google Scholar] [CrossRef]
- Sorrells, S.; Paredes, M.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.; James, D.; Mayer, S.; Chang, J.; Auguste, K.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Linsley, J.W.; Reisine, T.; Finkbeiner, S. Cell death assays for neurodegenerative disease drug discovery. Expert Opin. Drg Discov. 2019, 14, 901–913. [Google Scholar] [CrossRef]
- Buendia, I.; Michalska, P.; Navarro, E.; Gameiro, I.; Egea, J.; León, R. Nrf2-ARE pathway: An emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases. Pharmacol. Ther. 2016, 157, 84–104. [Google Scholar] [CrossRef]
- Amiri, A.; Barreto, G.; Sathyapalan, T.; Sahebkar, A. siRNA Therapeutics: Future Promise for Neurodegenerative Diseases. Curr. Neuropharmacol. 2021, 19, 1896–1911. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Wang, W.; Bian, J.; Gao, Y.; Hao, Z.; Tan, J. Recent advances in medicinal and edible homologous polysaccharides: Extraction, purification, structure, modification, and biological activities. Int. J. Biol. Macromol. 2022, 222, 1110–1126. [Google Scholar] [CrossRef] [PubMed]
- Benussi, A.; Cantoni, V.; Manes, M.; Libri, I.; Dell’Era, V.; Datta, A.; Thomas, C.; Ferrari, C.; Di Fonzo, A.; Fancellu, R.; et al. Motor and cognitive outcomes of cerebello-spinal stimulation in neurodegenerative ataxia. Brain J. Neurol. 2021, 144, 2310–2321. [Google Scholar] [CrossRef] [PubMed]
- Raha, S.; Ghosh, A.; Dutta, D.; Patel, D.; Pahan, K. Activation of PPARα enhances astroglial uptake and degradation of β-amyloid. Sci. Signal. 2021, 14, eabg4747. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, S.; Bennett, R.; Delorme, L.; Robbins, A.; Hu, M.; McKenzie, D.; Kirk, M.; Schiantarelli, J.; Tunio, N.; Amaral, A.; et al. Experimental evidence for the age dependence of tau protein spread in the brain. Sci. Adv. 2019, 5, eaaw6404. [Google Scholar] [CrossRef]
- Liu, Z.; Chan, R.; Cai, Z.; Liu, X.; Wu, Y.; Yu, Z.; Feng, T.; Yang, Y.; Zhang, J. α-Synuclein-containing erythrocytic extracellular vesicles: Essential contributors to hyperactivation of monocytes in Parkinson’s disease. J. Neuroinflamm. 2022, 19, 53. [Google Scholar] [CrossRef]
- Marchetti, B.; Tirolo, C.; L’Episcopo, F.; Caniglia, S.; Testa, N.; Smith, J.; Pluchino, S.; Serapide, M. Parkinson’s disease, aging and adult neurogenesis: Wnt/β-catenin signalling as the key to unlock the mystery of endogenous brain repair. Aging Cell 2020, 19, e13101. [Google Scholar] [CrossRef]
- Aladdin, A.; Király, R.; Boto, P.; Regdon, Z.; Tar, K. Juvenile Huntington’s Disease Skin Fibroblasts Respond with Elevated Parkin Level and Increased Proteasome Activity as a Potential Mechanism to Counterbalance the Pathological Consequences of Mutant Huntingtin Protein. Int. J. Mol. Sci. 2019, 20, 338. [Google Scholar] [CrossRef]
- Lu, A.; Narayan, P.; Grant, M.; Langfelder, P.; Wang, N.; Kwak, S.; Wilkinson, H.; Chen, R.; Chen, J.; Bawden, C.S.; et al. DNA methylation study of Huntington’s disease and motor progression in patients and in animal models. Nat. Commun. 2020, 11, 4529. [Google Scholar] [CrossRef]
- Yan, T.X.; Nian, T.T.; Wu, B.; Xiao, F.; He, B.S.; Bi, K.S.; Jia, Y. Okra polysaccharides can reverse the metabolic disorder induced by high-fat diet and cognitive function injury in A beta(1-42) mice. Exp. Gerontol. 2020, 130, 110802. [Google Scholar] [CrossRef]
- Lauer, A.; Grimm, H.; Apel, B.; Golobrodska, N.; Kruse, L.; Ratanski, E.; Schulten, N.; Schwarze, L.; Slawik, T.; Sperlich, S.; et al. Mechanistic Link between Vitamin B12 and Alzheimer’s Disease. Biomolecules 2022, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Nan, S.; Fan, J.; Chen, Q.; Zhang, Y. Inonotus obliquus polysaccharides protect against Alzheimer’s disease by regulating Nrf2 signaling and exerting antioxidative and antiapoptotic effects. Int. J. Biol. Macromol. 2019, 131, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, X.; Zhang, Y.; Liu, X.; Wang, C.; Teng, L.; Wang, D. Protective roles of Amanita caesarea polysaccharides against Alzheimer’s disease via Nrf2 pathway. Int. J. Biol. Macromol. 2019, 121, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Mao, J.; Ding, K.; Zhou, Y.; Zeng, X.; Yang, W.; Wang, P.; Zhao, C.; Yao, J.; Xia, P.; et al. Polysaccharides from Ganoderma lucidum Promote Cognitive Function and Neural Progenitor Proliferation in Mouse Model of Alzheimer’s Disease. Stem Cell Rep. 2017, 8, 84–94. [Google Scholar] [CrossRef]
- Guo, D.J.; Zhou, J.; Zhang, M.; Taximaimaiti, R.; Wang, X.P.; Wang, H. Momordica Charantia Polysaccharides Attenuates MPP+-Induced Injury in Parkinson’s Disease Mice and Cell Models by Regulating TLR4/MyD88/NF-kappa B Pathway. Int. J. Polym. Sci. 2021, 2021, 5575636. [Google Scholar] [CrossRef]
- Fang, F.; Peng, T.; Yang, S.; Wang, W.; Zhang, Y.; Li, H. Lycium barbarum polysaccharide attenuates the cytotoxicity of mutant huntingtin and increases the activity of AKT. Int. J. Dev. Neurosci. 2016, 52, 66–74. [Google Scholar] [CrossRef]
- Rolli, S.; Sontag, E. Spatial sequestration of misfolded proteins in neurodegenerative diseases. Biochem. Soc. Trans. 2022, 50, 759–771. [Google Scholar] [CrossRef]
- Mallucci, G.; Klenerman, D.; Rubinsztein, D.J. Developing Therapies for Neurodegenerative Disorders: Insights from Protein Aggregation and Cellular Stress Responses. Annu. Rev. Cell Dev. Biol. 2020, 36, 165–189. [Google Scholar] [CrossRef]
- Li, J.; Breker, M.; Graham, M.; Schuldiner, M.; Hochstrasser, M. AMPK regulates ESCRT-dependent microautophagy of proteasomes concomitant with proteasome storage granule assembly during glucose starvation. PLoS Genet. 2019, 15, e1008387. [Google Scholar] [CrossRef]
- Mao, K.; Chen, J.; Yu, H.; Li, H.; Ren, Y.; Wu, X.; Wen, Y.; Zou, F.; Li, W. Poly (ADP-ribose) polymerase 1 inhibition prevents neurodegeneration and promotes α-synuclein degradation via transcription factor EB-dependent autophagy in mutant α-synucleinA53T model of Parkinson’s disease. Aging Cell 2020, 19, e13163. [Google Scholar] [CrossRef]
- Tateishi-Karimata, H.; Sugimoto, N. Roles of non-canonical structures of nucleic acids in cancer and neurodegenerative diseases. Nucleic Acids Res. 2021, 49, 7839–7855. [Google Scholar] [CrossRef] [PubMed]
- Vaillant-Beuchot, L.; Mary, A.; Pardossi-Piquard, R.; Bourgeois, A.; Lauritzen, I.; Eysert, F.; Kinoshita, P.; Cazareth, J.; Badot, C.; Fragaki, K.; et al. Accumulation of amyloid precursor protein C-terminal fragments triggers mitochondrial structure, function, and mitophagy defects in Alzheimer’s disease models and human brains. Acta Neuropathol. 2021, 141, 39–65. [Google Scholar] [CrossRef] [PubMed]
- Franzmeier, N.; Ossenkoppele, R.; Brendel, M.; Rubinski, A.; Smith, R.; Kumar, A.; Mattsson-Carlgren, N.; Strandberg, O.; Duering, M.; Buerger, K.; et al. The BIN1 rs744373 Alzheimer’s disease risk SNP is associated with faster Aβ-associated tau accumulation and cognitive decline. Alzheimer Dement. J. Alzheimer Assoc. 2022, 18, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Petit, D.; Fernández, S.; Zoltowska, K.; Enzlein, T.; Ryan, N.; O’Connor, A.; Szaruga, M.; Hill, E.; Vandenberghe, R.; Fox, N.; et al. Aβ profiles generated by Alzheimer’s disease causing PSEN1 variants determine the pathogenicity of the mutation and predict age at disease onset. Mol. Psychiatry 2022, 27, 2821–2832. [Google Scholar] [CrossRef] [PubMed]
- Galvagnion, C.; Marlet, F.; Cerri, S.; Schapira, A.; Blandini, F.; Di Monte, D. Sphingolipid changes in Parkinson L444P GBA mutation fibroblasts promote α-synuclein aggregation. Brain J. Neurol. 2022, 145, 1038–1051. [Google Scholar] [CrossRef] [PubMed]
- Thomzig, A.; Wagenführ, K.; Pinder, P.; Joncic, M.; Schulz-Schaeffer, W.; Beekes, M. Transmissible α-synuclein seeding activity in brain and stomach of patients with Parkinson’s disease. Acta Neuropathol. 2021, 141, 861–879. [Google Scholar] [CrossRef] [PubMed]
- Stuendl, A.; Kunadt, M.; Kruse, N.; Bartels, C.; Moebius, W.; Danzer, K.; Mollenhauer, B.; Schneider, A. Induction of α-synuclein aggregate formation by CSF exosomes from patients with Parkinson’s disease and dementia with Lewy bodies. Brain J. Neurol. 2016, 139, 481–494. [Google Scholar] [CrossRef]
- Ast, A.; Buntru, A.; Schindler, F.; Hasenkopf, R.; Schulz, A.; Brusendorf, L.; Klockmeier, K.; Grelle, G.; McMahon, B.; Niederlechner, H.; et al. mHTT Seeding Activity: A Marker of Disease Progression and Neurotoxicity in Models of Huntington’s Disease. Mol. Cell 2018, 71, 675.e676–688.e6. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, T.; Shan, Y.; Li, G.; Ni, X.; Chen, X.; Hu, X.; Lin, L.; Li, Y.; Guan, Y.; et al. Therapeutic reversal of Huntington’s disease by in vivo self-assembled siRNAs. Brain J. Neurol. 2021, 144, 3421–3435. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, Q.; Luo, H.; Xu, Z.; Huang, S.; Yang, F.; Liu, Y.; Mahaman, Y.; Ke, D.; Wang, Q.; et al. Codonopsis pilosula polysaccharide attenuates Aβ toxicity and cognitive defects in APP/PS1 mice. Aging 2020, 12, 13422–13436. [Google Scholar] [CrossRef]
- Hu, W.J.; Song, M.K.; Wang, C.Y.; Guo, Z.; Li, Y.; Wang, D. Structural characterization of polysaccharide purified from Hericium erinaceus fermented mycelium and its pharmacological basis for application in Alzheimer’s disease: Oxidative stress related calcium homeostasis. Int. J. Biol. Macromol. 2021, 193, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Z.; Wei, M.; Hu, M.; Yue, K.; Bi, R.; Zhai, S.; Pi, Z.; Song, F. Liu and function, Pharmacodynamic and urinary metabolomics studies on the mechanism of Schisandra polysaccharide in the treatment of Alzheimer’s disease. Food Funct. 2019, 10, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xia, Y.Y.; Luo, H.B.; Huang, S.; Wang, Y.J.; Shentu, Y.P.; Mahaman, Y.A.R.; Huang, F.; Ke, D.; Wang, Q.; et al. Codonopsis pilosula Polysaccharide Attenuates Tau Hyperphosphorylation and Cognitive Impairments in hTau Infected Mice. Front. Mol. Neurosci. 2018, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pang, L.; Zhang, Y.; Xu, J.; Ding, D.; Yang, T.; Zhao, Q.; Wu, F.; Li, F.; Meng, H.; et al. Lycium barbarum Polysaccharide Promotes Nigrostriatal Dopamine Function by Modulating PTEN/AKT/mTOR Pathway in a Methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) Murine Model of Parkinson’s Disease. Neurochem. Res. 2018, 43, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Zhu, X.; Si, J. Angelica polysaccharide ameliorates memory impairment in Alzheimer’s disease rat through activating BDNF/TrkB/CREB pathway. Exp. Biol. Med. 2020, 245, 1–10. [Google Scholar] [CrossRef]
- Zhang, H.; Pan, N.; Xiong, S.; Zou, S.; Li, H.; Xiao, L.; Cao, Z.; Tunnacliffe, A.; Huang, Z. Inhibition of polyglutamine-mediated proteotoxicity by Astragalus membranaceus polysaccharide through the DAF-16/FOXO transcription factor in Caenorhabditis elegans. Biochem. J. 2012, 441, 417–424. [Google Scholar] [CrossRef]
- Otero-Garcia, M.; Mahajani, S.; Wakhloo, D.; Tang, W.; Xue, Y.; Morabito, S.; Pan, J.; Oberhauser, J.; Madira, A.; Shakouri, T.; et al. Molecular signatures underlying neurofibrillary tangle susceptibility in Alzheimer’s disease. Neuron 2022, 18, 2929–2948. [Google Scholar] [CrossRef]
- Stepanov, Y.; Golovynska, I.; Zhang, R.; Golovynskyi, S.; Stepanova, L.; Gorbach, O.; Dovbynchuk, T.; Garmanchuk, L.; Ohulchanskyy, T.; Qu, J. Near-infrared light reduces β-amyloid-stimulated microglial toxicity and enhances survival of neurons: Mechanisms of light therapy for Alzheimer’s disease. Alzheimer Res. Ther. 2022, 14, 84. [Google Scholar] [CrossRef]
- Almeida, L.; Pinho, B.; Duchen, M.; Oliveira, J. The PERKs of mitochondria protection during stress: Insights for PERK modulation in neurodegenerative and metabolic diseases. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1737–1748. [Google Scholar] [CrossRef]
- Dionísio, P.A.; Amaral, J.D.; Rodrigues, C.M.P. Oxidative stress and regulated cell death in Parkinson’s disease. Ageing Res. Rev. 2021, 67, 101263. [Google Scholar] [CrossRef]
- Fitzgerland, J.C.; Zimprich, A.; Carvajal, D.A.; Schindler, K.M.; Maurer, B.; Schulte, C.; Bus, C.; Hauser, A.K.; Kubler, M.; Lewin, R.; et al. Metformin reverses TRAP1 mutation-associated alterations in mitochondrial function in Parkinson’s disease. Brain 2017, 140, 2444–2459. [Google Scholar] [CrossRef] [PubMed]
- Lessard-Beaudoin, M.; Yu-Taeger, L.; Laroche, M.; Singer, E.; Riess, O.; Nguyen, H.; Graham, R. Olfactory bulb atrophy and caspase activation observed in the BACHD rat models of Huntington disease. Neurobiol. Dis. 2019, 125, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, H.; Zhang, X.; Li, X.; Geng, L.; Zhang, H.; Zhang, Q. Sulfated Hetero-Polysaccharides Protect SH-SY5Y Cells from H2O2-Induced Apoptosis by Affecting the PI3K/Akt Signaling Pathway. Mar. Drugs 2017, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; He, C.; Shen, Y.; Zeng, G.; Tian, D.; Cheng, Y.; Xu, M.; Fan, D.; Tan, C.; Shi, A.; et al. Polysaccharide Krestin Prevents Alzheimer’s Disease-type Pathology and Cognitive Deficits by Enhancing Monocyte Amyloid-β Processing. Neurosci. Bull. 2022, 38, 290–302. [Google Scholar] [CrossRef]
- Cenini, G.; Lloret, A.; Cascella, R. Oxidative Stress in Neurodegenerative Diseases: From a Mitochondrial Point of View. Oxidative Med. Cell. Longev. 2019, 2019, 2105607. [Google Scholar] [CrossRef]
- Delp, J.; Cediel-Ulloa, A.; Suciu, I.; Kranaster, P.; van Vugt-Lussenburg, B.; Kos, V.M.; van der Stel, W.; Carta, G.; Bennekou, S.; Jennings, P.; et al. Neurotoxicity and underlying cellular changes of 21 mitochondrial respiratory chain inhibitors. Arch. Toxicol. 2021, 95, 591–615. [Google Scholar] [CrossRef]
- Kocabas, F.; Zheng, J.K.; Thet, S.; Copeland, N.G.; Jenkins, N.A.; DeBerardinis, R.J.; Zhang, C.C.; Sadek, H.A. Meis1 regulates the metabolic phenotype and oxidant defense of hematopoietic stem cells. Blood 2012, 120, 4963–4972. [Google Scholar] [CrossRef]
- Hassan, A.H.A.; Alkhalifah, D.H.M.; Al Yousef, S.A.; Beemster, G.T.S.; Mousa, A.S.M.; Hozzein, W.N.; AbdElgawad, H. Salinity Stress Enhances the Antioxidant Capacity ofBacillusandPlanococcusSpecies Isolated from Saline Lake Environment. Front. Microbiol. 2020, 11, 581816. [Google Scholar] [CrossRef]
- Yang, S.S.; Lian, G.J. ROS and diseases: Role in metabolism and energy supply. Mol. Cell. Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef]
- Shi, L.; Hu, F.; Duan, Y.; Wu, W.; Dong, J.; Meng, X.; Zhu, X.; Liu, B. Hybrid Nanospheres to Overcome Hypoxia and Intrinsic Oxidative Resistance for Enhanced Photodynamic Therapy. ACS Nano 2020, 14, 2183–2190. [Google Scholar] [CrossRef]
- Wang, X.Y.; Zhou, Y.X.; Gao, Q.N.; Ping, D.N.; Wang, Y.L.; Wu, W.; Lin, X.; Fang, Y.J.; Zhang, J.M.; Shao, A.W. The Role of Exosomal microRNAs and Oxidative Stress in Neurodegenerative Diseases. Oxidative Med. Cell. Longev. 2020, 2020, 3232869. [Google Scholar] [CrossRef] [PubMed]
- Höhn, A.; Tramutola, A.; Cascella, R. Proteostasis Failure in Neurodegenerative Diseases: Focus on Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 5497046. [Google Scholar] [CrossRef]
- Trist, B.; Hare, D.; Double, K. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, M.; Terzioglu, M.; Galter, D.; Zhu, S.; Hofstetter, C.; Lindqvist, E.; Thams, S.; Bergstrand, A.; Hansson, F.; Trifunovic, A.; et al. Progressive parkinsonism in mice with respiratory-chain-deficient dopamine neurons. Proc. Natl. Acad. Sci. USA 2007, 104, 1325–1330. [Google Scholar] [CrossRef]
- He, J.Y.; Zhu, G.F.; Wang, G.Q.; Zhang, F. Oxidative Stress and Neuroinflammation Potentiate Each Other to Promote Progression of Dopamine Neurodegeneration. Oxidative Med. Cell. Longev. 2020, 2020, 6137521. [Google Scholar] [CrossRef] [PubMed]
- Song, W.J.; Chen, J.; Petrilli, A.; Liot, G.; Klinglmayr, E.; Zhou, Y.; Poquiz, P.; Tjong, J.; Pouladi, M.A.; Hayden, M.R.; et al. Mutant huntingtin binds the mitochondrial fission GTPase dynamin-related protein-1 and increases its enzymatic activity. Nat. Med. 2011, 17, 377–382. [Google Scholar] [CrossRef]
- Habaike, A.; Yakufu, M.; Cong, Y.Y.; Gahafu, Y.; Li, Z.; Abulizi, P. Neuroprotective effects of Fomes officinalis Ames polysaccharides on A beta(25–35)-induced cytotoxicity in PC12 cells through suppression of mitochondria-mediated apoptotic pathway. Cytotechnology 2020, 72, 539–549. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhang, J.; Li, H.; Wang, Q.; Xiao, L.; Weng, H.; Zhou, X.; Ma, C.; Ma, F.; Hu, M.; et al. Epimedium Polysaccharide Alleviates Polyglutamine-Induced Neurotoxicity in Caenorhabditis elegans by Reducing Oxidative Stress. Rejuvenation Res. 2017, 20, 32–41. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, J.W.; Zheng, Y.; Su, R.J.; Liao, Y.J.; Gong, X.L.; Liu, L.M.; Wang, X.M. Fucoidan Protects Dopaminergic Neurons by Enhancing the Mitochondrial Function in a Rotenone-induced Rat Model of Parkinson’s Disease. Aging Dis. 2018, 9, 590–604. [Google Scholar] [CrossRef]
- Harms, A.; Ferreira, S.; Romero-Ramos, M. Periphery and brain, innate and adaptive immunity in Parkinson’s disease. Acta Neuropathol. 2021, 141, 527–545. [Google Scholar] [CrossRef]
- Saimaiti, A.; Zhou, D.D.; Li, J.H.; Xiong, R.G.; Gan, R.Y.; Huang, S.Y.; Shang, A.; Zhao, C.N.; Li, H.Y.; Li, H.B. Dietary sources, health benefits, and risks of caffeine. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hammond, T.; Robinton, D.; Stevens, B.J. Microglia and the Brain: Complementary Partners in Development and Disease. Annu. Rev. Cell Dev. Biol. 2018, 34, 523–544. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, C.; Friedman, B.; Dejanovic, B.; Sheng, M. Microglia in Brain Development, Homeostasis, and Neurodegeneration. Annu. Rev. Genet. 2019, 53, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, M.; Kim, D.; Shin, M.; Son, O.; Jo, H.; Yeo, H.; Cho, S.; Park, J.; Lee, C.; et al. Transduced PEP-1-PON1 proteins regulate microglial activation and dopaminergic neuronal death in a Parkinson’s disease model. Biomaterials 2015, 64, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.; Hansen, D.; Sheng, M. TREM2, Microglia, and Neurodegenerative Diseases. Trends Mol. Med. 2017, 23, 512–533. [Google Scholar] [CrossRef]
- Yang, H.; Li, X.; Zhu, L.; Wu, X.; Zhang, S.; Huang, F.; Feng, X.; Shi, L. Heat Shock Protein Inspired Nanochaperones Restore Amyloid-β Homeostasis for Preventative Therapy of Alzheimer’s Disease. Adv. Sci. 2019, 6, 1901844. [Google Scholar] [CrossRef]
- Olanow, C.; Savolainen, M.; Chu, Y.; Halliday, G.; Kordower, J. Temporal evolution of microglia and α-synuclein accumulation following foetal grafting in Parkinson’s disease. Brain J. Neurol. 2019, 142, 1690–1700. [Google Scholar] [CrossRef]
- Guo, M.; Wang, J.; Zhao, Y.; Feng, Y.; Han, S.; Dong, Q.; Cui, M.; Tieu, K. Microglial exosomes facilitate α-synuclein transmission in Parkinson’s disease. Brain J. Neurol. 2020, 143, 1476–1497. [Google Scholar] [CrossRef]
- Xu, M.; Yan, T.; Fan, K.; Wang, M.; Qi, Y.; Xiao, F.; Bi, K.; Jia, Y. Polysaccharide of Schisandra Chinensis Fructus ameliorates cognitive decline in a mouse model of Alzheimer’s disease. J. Ethnopharmacol. 2019, 237, 354–365. [Google Scholar] [CrossRef]
- Shin, S.J.; Nam, Y.; Park, Y.H.; Kim, M.J.; Lee, E.; Jeon, S.G.; Bae, B.S.; Seo, J.; Shim, S.L.; Kim, J.S.; et al. Therapeutic effects of non-saponin fraction with rich polysaccharide from Korean red ginseng on aging and Alzheimer’s disease. Free. Radic. Biol. Med. 2021, 164, 233–248. [Google Scholar] [CrossRef]
- Chen, P.B.; Wang, H.C.; Liu, Y.W.; Lin, S.H.; Chou, H.N.; Sheen, L.Y. Immunomodulatory activities of polysaccharides from Chlorella pyrenoidosa in a mouse model of Parkinson’s disease. J. Funct. Foods 2014, 11, 103–113. [Google Scholar] [CrossRef]
- Lin, Y.W.; Fang, C.H.; Liang, Y.J.; Liao, H.H.; Lin, F.H. Modified Low-Temperature Extraction Method for Isolation of Bletilla striata Polysaccharide as Antioxidant for the Prevention of Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 760. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Qin, J.; Liu, M.; Luo, J.; Ding, F.; Wang, M.; Zheng, L. Astragalus polysaccharide attenuates lipopolysaccharide-induced inflammatory responses in microglial cells: Regulation of protein kinase B and nuclear factor-κB signaling. Inflamm. Res. 2015, 64, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Guo, L.; Yang, Y.; Li, W.; Sheng, Y.; Wang, J.; Guan, Q.; Zhang, X. Study on antrodia camphorata polysaccharide in alleviating the neuroethology of PD mice by decreasing the expression of NLRP3 inflammasome. Phytother. Res. PTR 2019, 33, 2288–2297. [Google Scholar] [CrossRef]
- Han, C.Y.; Shen, H.P.; Yang, Y.; Sheng, Y.J.; Wang, J.; Li, W.Y.; Zhou, X.H.; Guo, L.; Zhai, L.P.; Guan, Q.B. Antrodia camphorata polysaccharide resists 6-OHDA-induced dopaminergic neuronal damage by inhibiting ROS-NLRP3 activation. Brain Behav. 2020, 10, 1824. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Guo, Y.H.; Cheng, Y.L.; Yao, W.R.; Qian, H. Neuroprotective effects of polysaccharide from Sparassis crispa on Alzheimer’s disease-like mice: Involvement of microbiota-gut-brain axis. Int. J. Biol. Macromol. 2023, 225, 974–986. [Google Scholar] [CrossRef]
- Luo, S.L.; Zhang, X.; Huang, S.; Feng, X.P.; Zhang, X.J.; Xiang, D.X. A monomeric polysaccharide from Polygonatum sibiricum improves cognitive functions in a model of Alzheimer’s disease by reshaping the gut microbiota. Int. J. Biol. Macromol. 2022, 213, 404–415. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Ho, Y.S.; Chang, R.C.C. Linking circadian rhythms to microbiome-gut-brain axis in aging-associated neurodegenerative diseases. Ageing Res. Rev. 2022, 78, 101620. [Google Scholar] [CrossRef]
- Tao, J.; An, Y.; Xu, L.; Wang, Y.; Wang, C.; Li, P.; Li, M.; Yan, D.; Wang, M.; Zhong, G.; et al. The protective role of microbiota in the prevention of MPTP/P-induced Parkinson’s disease by resveratrol. Food Funct. 2023, 2042–6496. [Google Scholar] [CrossRef]
- Van Den Berge, N.; Ferreira, N.; Mikkelsen, T.; Alstrup, A.; Tamgüney, G.; Karlsson, P.; Terkelsen, A.; Nyengaard, J.; Jensen, P.; Borghammer, P. Ageing promotes pathological alpha-synuclein propagation and autonomic dysfunction in wild-type rats. Brain J. Neurol. 2021, 144, 1853–1868. [Google Scholar] [CrossRef]
- Ahn, E.; Liu, X.; Alam, A.; Kang, S.; Ye, K. Helicobacter hepaticus augmentation triggers Dopaminergic degeneration and motor disorders in mice with Parkinson’s disease. Mol. Psychiatry 2023, 28, 1337–1350. [Google Scholar] [CrossRef]
- Smith, M.; Wade, J.; Wolstenholme, J.; Bajaj, J. Gut microbiome-brain-cirrhosis axis. Hepatology 2023, 10, 1097. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, M.; Quan, W.; Huang, W.; Liu, X.; Zhang, C.; Lu, B.; Xiao, X.; Liu, Z. Acteoside palliates d-galactose induced cognitive impairment by regulating intestinal homeostasis. Food Chem. 2023, 421, 135978. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, S.; Wang, Y.; Fan, M.; Dai, J.; Zhu, C.; Xu, K.; Cui, M.; Suo, C.; Jin, L.; et al. Metagenomic association analysis of cognitive impairment in community-dwelling older adults. Neurobiol. Dis. 2023, 180, 106081. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Chen, Y.; Song, X.; Dou, X.; Xu, C. Lactobacillus caseiSelenium Nanoparticles-Enriched ATCC 393 Prevents Cognitive Dysfunction in Mice Through Modulating Microbiota-Gut-Brain Axis. Int. J. Nanomed. 2022, 17, 4807–4827. [Google Scholar] [CrossRef]
- Zhu, G.; Zhao, J.; Wang, G.; Chen, W. Bifidobacterium breve HNXY26M4 Attenuates Cognitive Deficits and Neuroinflammation by Regulating the Gut-Brain Axis in APP/PS1 Mice. J. Agric. Food Chem. 2023, 71, 4646–4655. [Google Scholar] [CrossRef]
- Sánchez-Tapia, M.; Mimenza-Alvarado, A.; Granados-Domínguez, L.; Flores-López, A.; López-Barradas, A.; Ortiz, V.; Pérez-Cruz, C.; Sánchez-Vidal, H.; Hernández-Acosta, J.; Ávila-Funes, J.; et al. The Gut Microbiota-Brain Axis during Aging, Mild Cognitive Impairment and Dementia: Role of Tau Protein, β-Amyloid and LPS in Serum and Curli Protein in Stool. Nutrients 2023, 15, 932. [Google Scholar] [CrossRef]
- Vidal, R.; Matus, S.; Bargsted, L.; Hetz, C. Targeting autophagy in neurodegenerative diseases. Trends Pharmacol. Sci. 2014, 35, 583–591. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, X.; Song, Y.; Tu, J. Autophagy in Alzheimer’s disease pathogenesis: Therapeutic potential and future perspectives. Ageing Res. Rev. 2021, 72, 101464. [Google Scholar] [CrossRef]
- Tan, Y.; Yin, L.; Sun, Z.; Shao, S.; Chen, W.; Man, X.; Du, Y.; Chen, Y. Astragalus polysaccharide exerts anti-Parkinson via activating the PI3K/AKT/mTOR pathway to increase cellular autophagy level in vitro. Int. J. Biol. Macromol. 2020, 153, 349–356. [Google Scholar] [CrossRef]
- Li, Y.Q.; Peng, Y.; Shen, Y.B.; Zhang, Y.Z.; Liu, L.L.; Yang, X.Q. Dietary polyphenols: Regulate the advanced glycation end products-RAGE axis and the microbiota-gut-brain axis to prevent neurodegenerative diseases. Crit. Rev. Food Sci. Nutr. 2022, 5, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lu, J.; Zhang, J.G.; Xie, J.X. Protective effects of a polysaccharide from Spirulina platensis on dopaminergic neurons in an MPTP-induced Parkinson’s disease model in C57BL/6J mice. Neural Regen. Res. 2015, 10, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wei, Y.; Xu, J.; Wei, X.L. A comprehensive review on the prevention and regulation of Alzheimer’s disease by tea and its active ingredients. Crit. Rev. Food Sci. Nutr. 2022, 6, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Duan, Y.; Huang, S.; Zhou, X.; Zhou, L.; Hu, T.; Yang, Y.; Lu, J.; Ding, K.; Guo, D.; et al. Polysaccharides from Lycium barbarum ameliorate amyloid pathology and cognitive functions in APP/PS1 transgenic mice. Int. J. Biol. Macromol. 2020, 144, 1004–1012. [Google Scholar] [CrossRef]
- Mollazadeh, H.; Tavana, E.; Fanni, G.; Bo, S.; Banach, M.; Pirro, M.; von Haehling, S.; Jamialahmadi, T.; Sahebkar, A. Effects of statins on mitochondrial pathways. J. Cachexia Sarcopenia Muscle 2021, 12, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.X.; Wang, J.; Hu, J.; Lou, G.H.; Xiong, H.J.; Peng, C.Y.; Huang, Q.W. Modulation of Apoptosis by Plant Polysaccharides for Exerting Anti-Cancer Effects: A Review. Front. Pharmacol. 2020, 11, 792. [Google Scholar] [CrossRef]
- Yu, W.; Lin, C.; Zhao, Q.; Lin, X.; Dong, X. Neuroprotection against hydrogen peroxide-induced toxicity by Dictyophora echinovolvata polysaccharide via inhibiting the mitochondria-dependent apoptotic pathway. Biomed. Pharmacother. 2017, 88, 569–573. [Google Scholar] [CrossRef]
- Jin, Y.; Hu, X.; Zhang, Y.; Liu, T. Studies on the purification of polysaccharides separated from Tremella fuciformis and their neuroprotective effect. Mol. Med. Rep. 2016, 13, 3985–3992. [Google Scholar] [CrossRef]
- Hu, S.; Wang, D.; Zhang, J.; Du, M.; Cheng, Y.; Liu, Y.; Zhang, N.; Wang, D.; Wu, Y. Mitochondria Related Pathway Is Essential for Polysaccharides Purified from Sparassis crispa Mediated Neuro-Protection against Glutamate-Induced Toxicity in Differentiated PC12 Cells. Int. J. Mol. Sci. 2016, 17, 133. [Google Scholar] [CrossRef]
- An, S.; Lu, W.; Zhang, Y.; Yuan, Q.; Wang, D. Armillaria melleaPharmacological Basis for Use of Polysaccharides in Alzheimer’s Disease: Antiapoptosis and Antioxidation. Oxidative Med. Cell. Longev. 2017, 2017, 4184562. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, L.; Zhang, T.; Luo, D.; Jia, Y.; Guo, Z.; Zhang, Q.; Wang, X.; Wang, X. Inhibitory effect of fucoidan on nitric oxide production in lipopolysaccharide-activated primary microglia. Clin. Exp. Pharmacol. Physiol. 2010, 37, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Li, Q.; Chen, C.; Chen, Z.; Huang, W. Neuroprotective effect of crude polysaccharide isolated from the fruiting bodies of Morchella importuna against HO-induced PC12 cell cytotoxicity by reducing oxidative stress. Biomed. Pharm. Ther. 2016, 83, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, T.G. Role of Nuclear Factor Kappa B (NF-kappa B) Signalling in Neurodegenerative Diseases: An Mechanistic Approach. Curr. Neuropharmacol. 2020, 18, 918–935. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Qiu, X.; Yu, Q.; Chen, H.Y.; Yan, C.Y. A novel polysaccharide from Acorus tatarinowii protects against LPS-induced neuroinflammation and neurotoxicity by inhibiting TLR4-mediated MyD88/NF-kappa B and PI3K/Akt signaling pathways. Int. J. Biol. Macromol. 2020, 163, 464–475. [Google Scholar] [CrossRef]
- Li, Q.; Xing, S.S.; Chen, Y.; Liao, Q.H.; Li, Q.H.; Liu, Y.; He, S.Y.; Feng, F.; Chen, Y.; Zhang, J.; et al. Reasonably activating Nrf2: A long-term, effective and controllable strategy for neurodegenerative diseases. Eur. J. Med. Chem. 2020, 185, 111862. [Google Scholar] [CrossRef]
- Qin, X.; Hua, J.; Lin, S.; Zheng, H.; Wang, J.; Li, W.; Ke, J.; Cai, H. Astragalus polysaccharide alleviates cognitive impairment and β-amyloid accumulation in APP/PS1 mice via Nrf2 pathway. Biochem. Biophys. Res. Commun. 2020, 531, 431–437. [Google Scholar] [CrossRef]
- Huang, S.; Yuan, H.Y.; Li, W.Q.; Liu, X.Y.; Zhang, X.J.; Xiang, D.X.; Luo, S.L. Polygonatum sibiricum Polysaccharides Protect against MPP-Induced Neurotoxicity via the Akt/mTOR and Nrf2 Pathways. Oxidative Med. Cell. Longev. 2021, 2021, 8843899. [Google Scholar] [CrossRef]
- Szwed, A.; Kim, E.; Jacinto, E. Regulation and metabolic functions of mtorc1 and mtorc2. Physiol. Rev. 2021, 101, 1371–1426. [Google Scholar] [CrossRef]
- Rai, S.N.; Dilnashin, H.; Birla, H.; Singh, S.S.; Zahra, W.; Rathore, A.S.; Singh, B.K.; Singh, S.P. The Role of PI3K/Akt and ERK in Neurodegenerative Disorders. Neurotox. Res. 2019, 35, 775–795. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y. Lycium barbarum polysaccharides alleviate hydrogen peroxide-induced injury by up-regulation of miR-4295 in human trabecular meshwork cells. Exp. Mol. Pathol. 2019, 106, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Pan, X.; Li, T.; Zhang, C.; Shi, N. Lycium barbarum Polysaccharides Protect against Trimethyltin Chloride-Induced Apoptosis via Sonic Hedgehog and PI3K/Akt Signaling Pathways in Mouse Neuro-2a Cells. Oxidative Med. Cell. Longev. 2016, 2016, 9826726. [Google Scholar] [CrossRef] [PubMed]
- Kou, L.; Du, M.; Zhang, C.; Dai, Z.; Li, X.; Zhang, B.; Hu, X. Polysaccharide purified from Lycium barbarum protects differentiated PC12 cells against L-Glu-induced toxicity via the mitochondria-associated pathway. Mol. Med. Rep. 2017, 16, 5533–5540. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cao, Y.; Chen, L.; Wang, J.; Tian, Q.; Wang, N.; Liu, Z.; Li, J.; Wang, N.; Wang, X.; et al. A polysaccharide from Polygonatum sibiricum attenuates amyloid-β-induced neurotoxicity in PC12 cells. Carbohydr. Polym. 2015, 117, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Du, J.; Günther, S.; Guo, X.; Wang, S.; Schneider, A.; Zhu, L.; Braun, T. Mechano-signaling via Piezo1 prevents activation and p53-mediated senescence of muscle stem cells. Redox Biol. 2022, 52, 102309. [Google Scholar] [CrossRef] [PubMed]
- Wyss-Coray, T. Ageing, neurodegeneration and brain rejuvenation. Nature 2016, 539, 180–186. [Google Scholar] [CrossRef]
- Cheng, X.; Yao, H.; Xiang, Y.; Chen, L.; Xiao, M.; Wang, Z.; Xiao, H.; Wang, L.; Wang, S.; Wang, Y. Effect of Angelica polysaccharide on brain senescence of Nestin-GFP mice induced by D-galactose. Neurochem. Int. 2019, 122, 149–156. [Google Scholar] [CrossRef]
- Wang, R.; Holsinger, R. Exercise-induced brain-derived neurotrophic factor expression: Therapeutic implications for Alzheimer’s dementia. Ageing Res. Rev. 2018, 48, 109–121. [Google Scholar] [CrossRef]
- Qin, X.; Zhao, Y.; Zhang, T.; Yin, C.; Qiao, J.; Guo, W.; Lu, B. TrkB agonist antibody ameliorates fertility deficits in aged and cyclophosphamide-induced premature ovarian failure model mice. Nat. Commun. 2022, 13, 914. [Google Scholar] [CrossRef]
- Rabie, M.; Ibrahim, H.; Nassar, N.; Atef, R. Adenosine A receptor agonist, N6-cyclohexyladenosine, attenuates Huntington’s disease via stimulation of TrKB/PI3K/Akt/CREB/BDNF pathway in 3-nitropropionic acid rat model. Chem.-Biol. Interact. 2023, 369, 110288. [Google Scholar] [CrossRef]
- Lin, T.; Chang, K.; Chiu, Y.; Weng, Z.; Sun, Y.; Lin, W.; Lee-Chen, G.; Chen, C. Neuroprotective Action of Coumarin Derivatives through Activation of TRKB-CREB-BDNF Pathway and Reduction of Caspase Activity in Neuronal Cells Expressing Pro-Aggregated Tau Protein. Int. J. Mol. Sci. 2022, 23, 2734. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.; Lin, T.; Chang, K.; Lin, W.; Hsieh-Li, H.; Su, M.; Chen, C.; Sun, Y.; Lee-Chen, G. Novel TRKB agonists activate TRKB and downstream ERK and AKT signaling to protect Aβ-GFP SH-SY5Y cells against Aβ toxicity. Aging 2022, 14, 7568–7586. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Du, J.; Hei, Q. Lycium barbarum polysaccharide protects against neurotoxicity via the Nrf2-HO-1 pathway. Exp. Ther. Med. 2017, 14, 4919–4927. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.S.; Liao, W.F.; Chen, X.; Yue, H.; Li, S.J.; Ding, K. An arabinogalactan from fruits of Lycium barbarum L. inhibits production and aggregation of A beta(42). Carbohydr. Polym. 2018, 195, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Liu, M.; Cao, J.; Yao, M.; Lu, Y.; Li, J.; Zhu, X.; Yang, Z.; Wen, A. Protective effects of Lycium barbarum polysaccharide on 6-OHDA-induced apoptosis in PC12 cells through the ROS-NO pathway. Molecules 2014, 20, 293–308. [Google Scholar] [CrossRef]
- Hu, Y.R.; Xing, S.; Chen, C.A.; Shen, D.; Chen, J. Codonopsis pilosula Polysaccharides Alleviate ABETA(1–40)-Induced PC12 Cells Energy Dysmetabolism via CD38/NAD+ Signaling Pathway. Curr. Alzheimer Res. 2021, 18, 208–221. [Google Scholar] [CrossRef]
- Huang, Y.C.; Tsay, H.J.; Lu, M.K.; Lin, C.H.; Yeh, C.W.; Liu, H.K.; Shiao, Y.J. Astragalus membranaceus-Polysaccharides Ameliorates Obesity, Hepatic Steatosis, Neuroinflammation and Cognition Impairment without Affecting Amyloid Deposition in Metabolically Stressed APPswe/PS1dE9 Mice. Int. J. Mol. Sci. 2017, 18, 2746. [Google Scholar] [CrossRef]
- Li, H.F.; Shi, R.N.; Ding, F.; Wang, H.Y.; Han, W.J.; Ma, F.L.; Hu, M.H.; Ma, C.W.; Huang, Z.B. Astragalus Polysaccharide Suppresses 6-Hydroxydopamine-Induced Neurotoxicity in Caenorhabditis elegans. Oxidative Med. Cell. Longev. 2016, 2016, 4856761. [Google Scholar] [CrossRef]
- Wang, X.L.; Yi, K.X.; Zhao, Y. Fucoidan inhibits amyloid-beta-induced toxicity in transgenic Caenorhabditis elegans by reducing the accumulation of amyloid-beta and decreasing the production of reactive oxygen species. Food Funct. 2018, 9, 552–560. [Google Scholar] [CrossRef]
- Xing, M.M.; Li, G.Y.; Liu, Y.; Yang, L.Y.; Zhang, Y.J.; Zhang, Y.R.; Ding, J.H.; Lu, M.; Yu, G.L.; Hu, G. Fucoidan from Fucus vesiculosus prevents the loss of dopaminergic neurons by alleviating mitochondrial dysfunction through targeting ATP5F1a. Carbohydr. Polym. 2023, 303, 120470. [Google Scholar] [CrossRef]
- Ma, L.J.; Wang, X.M.; Li, Y.; Xiao, H.; Yuan, F. Effect of polysaccharides from Vitis vinifera L. on NF-kappa B/I kappa B-alpha signal pathway and inflammatory factors in Alzheimer’s model rats. Biotechnol. Biotechnol. Equip. 2018, 32, 1012–1020. [Google Scholar] [CrossRef]
- Zhang, S.; Li, L.; Hu, J.; Ma, P.; Zhu, H. Taxus chinensis Polysaccharide of var. Cheng et L.K.Fu attenuates neurotoxicity and cognitive dysfunction in mice with Alzheimer’s disease. Pharm. Biol. 2020, 58, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, X.M.; Jin, G.; Yang, X.D.; Zhang, Y. Polysaccharides from Pleurotus ostreatus alleviate cognitive impairment in a rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 92, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.T.; Zhong, J.; Zhang, Q.; Yan, C.Y. Structural characterization and antineuroinflammatory activity of a novel heteropolysaccharide obtained from the fruits of Alpinia oxyphylla. Carbohydr. Polym. 2020, 229, 115405. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Yang, X. Protective effects of Gynostemma pentaphyllum polysaccharides on PC12 cells impaired by MPP(+). Int. J. Biol. Macromol. 2014, 69, 171–175. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, R.; Li, H.; Xiang, Y.; Xiao, L.; Hu, M.; Ma, F.; Ma, C.; Huang, Z. Antioxidant and neuroprotective effects of Dictyophora indusiata polysaccharide in Caenorhabditis elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, Q.; Ding, Y.; Peng, M.; Chen, L.; Dong, J.; Hu, J.; Ma, Y. The Potential of Edible and Medicinal Resource Polysaccharides for Prevention and Treatment of Neurodegenerative Diseases. Biomolecules 2023, 13, 873. https://doi.org/10.3390/biom13050873
Gan Q, Ding Y, Peng M, Chen L, Dong J, Hu J, Ma Y. The Potential of Edible and Medicinal Resource Polysaccharides for Prevention and Treatment of Neurodegenerative Diseases. Biomolecules. 2023; 13(5):873. https://doi.org/10.3390/biom13050873
Chicago/Turabian StyleGan, Qingxia, Yugang Ding, Maoyao Peng, Linlin Chen, Jijing Dong, Jiaxi Hu, and Yuntong Ma. 2023. "The Potential of Edible and Medicinal Resource Polysaccharides for Prevention and Treatment of Neurodegenerative Diseases" Biomolecules 13, no. 5: 873. https://doi.org/10.3390/biom13050873