
Organoids: Construction and Application in Gastric Cancer
 ,
,
Abstract
:1. Introduction
2. Organoids Definition and Characteristics
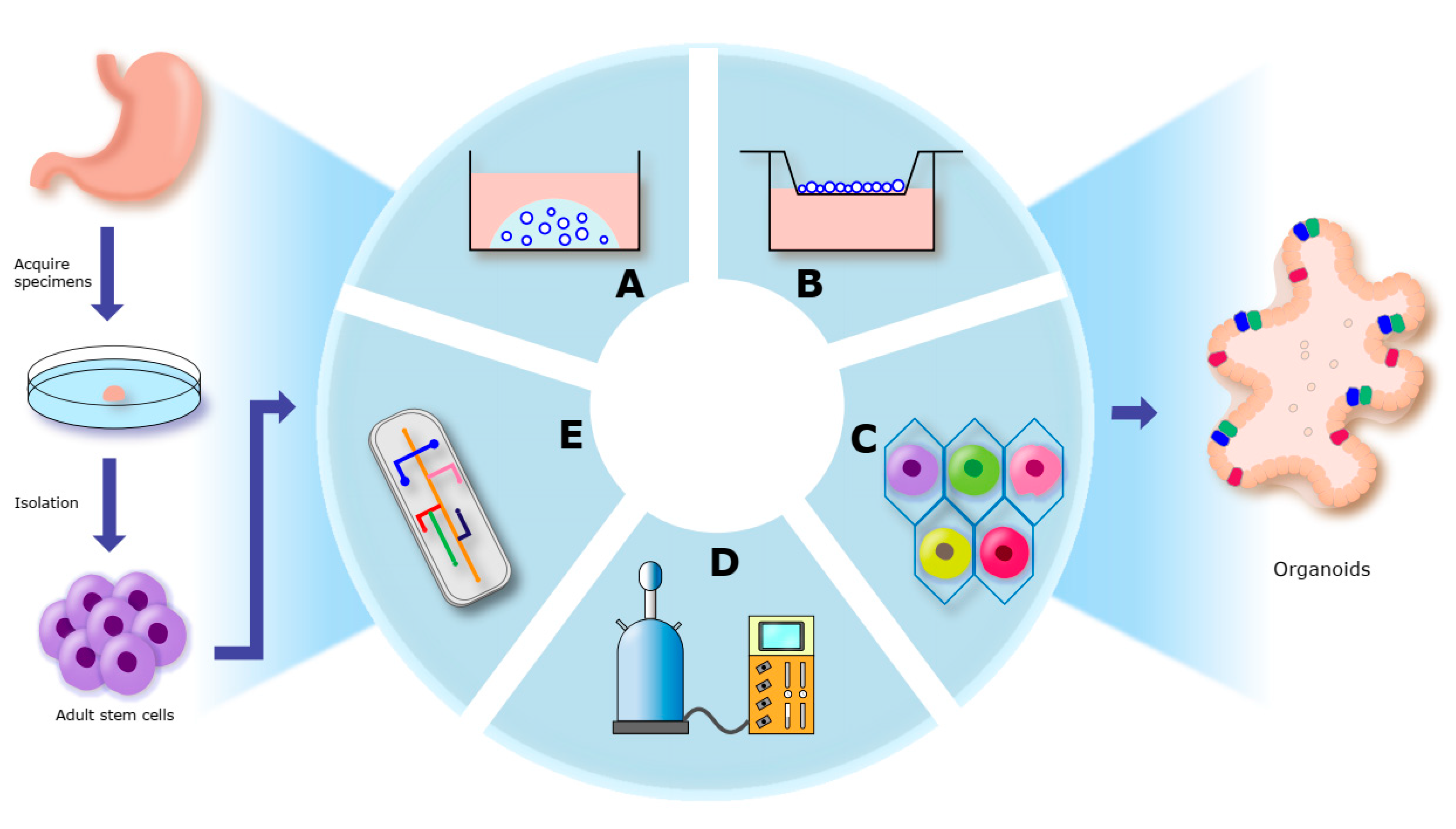
3. Construction of Organoids
3.1. Cells in Organoids Culture
3.2. Organoid Culture Systems
3.2.1. Culture Media
3.2.2. Extracellular Matrix Materials
3.2.3. Organoid Culture Modes
4. Application of Gastric Organoids
4.1. Tissue Engineering
4.2. Gastrointestinal Cancer–Microorganism Interaction
4.3. Drug Testing and Development
5. Limitations of Organoid Models
6. Concluding Remarks and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thrift, A.P.; El-Serag, H.B. Burden of Gastric Cancer. Clin. Gastroenterol. Hepatol. 2020, 18, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, K.; Clevers, H. Organoids: Modeling Development and the Stem Cell Niche in a Dish. Dev. Cell 2016, 38, 590–600. [Google Scholar] [CrossRef]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: Mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Hu, W.; Matulay, J.T.; Silva, M.V.; Owczarek, T.B.; Kim, K.; Chua, C.W.; Barlow, L.J.; Kandoth, C.; Williams, A.B.; et al. Tumor Evolution and Drug Response in Patient-Derived Organoid Models of Bladder Cancer. Cell 2018, 173, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Boj, S.F.; Hwang, C.I.; Baker, L.A.; Chio, I.I.C.; Engle, D.D.; Corbo, V.; Jager, M.; Ponz-Sarvise, M.; Tiriac, H.; Spector, M.S.; et al. Organoid models of human and mouse ductal pancreatic cancer. Cell 2015, 160, 324–338. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.H.N.; Siu, H.C.; Law, S.; Ho, S.L.; Yue, S.S.K.; Tsui, W.Y.; Chan, D.; Chan, A.S.; Ma, S.; Lam, K.O.; et al. A Comprehensive Human Gastric Cancer Organoid Biobank Captures Tumor Subtype Heterogeneity and Enables Therapeutic Screening. Cell Stem Cell 2018, 23, 882–897.e811. [Google Scholar] [CrossRef]
- Li, X.; Francies, H.E.; Secrier, M.; Perner, J.; Miremadi, A.; Galeano-Dalmau, N.; Barendt, W.J.; Letchford, L.; Leyden, G.M.; Goffin, E.K.; et al. Organoid cultures recapitulate esophageal adenocarcinoma heterogeneity providing a model for clonality studies and precision therapeutics. Nat. Commun. 2018, 9, 2983. [Google Scholar] [CrossRef] [PubMed]
- Girda, E.; Huang, E.C.; Leiserowitz, G.S.; Smith, L.H. The Use of Endometrial Cancer Patient-Derived Organoid Culture for Drug Sensitivity Testing Is Feasible. Int. J. Gynecol. Cancer 2017, 27, 1701–1707. [Google Scholar] [CrossRef]
- Pompaiah, M.; Bartfeld, S. Gastric Organoids: An Emerging Model System to Study Helicobacter pylori Pathogenesis. Curr. Top. Microbiol. Immunol. 2017, 400, 149–168. [Google Scholar] [CrossRef]
- Liu, L.; Yu, L.; Li, Z.; Li, W.; Huang, W. Patient-derived organoid (PDO) platforms to facilitate clinical decision making. J. Transl. Med. 2021, 19, 40. [Google Scholar] [CrossRef]
- Saito, Y.; Muramatsu, T.; Kanai, Y.; Ojima, H.; Sukeda, A.; Hiraoka, N.; Arai, E.; Sugiyama, Y.; Matsuzaki, J.; Uchida, R.; et al. Establishment of Patient-Derived Organoids and Drug Screening for Biliary Tract Carcinoma. Cell Rep. 2019, 27, 1265–1276.e1264. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, E.; Kolders, S.; Spelier, S.; Lõhmussaar, K.; Willems, S.M.; Devriese, L.A.; de Bree, R.; de Ruiter, E.J.; Korving, J.; Begthel, H.; et al. Oral Mucosal Organoids as a Potential Platform for Personalized Cancer Therapy. Cancer Discov. 2019, 9, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Fendler, A.; Bauer, D.; Busch, J.; Jung, K.; Wulf-Goldenberg, A.; Kunz, S.; Song, K.; Myszczyszyn, A.; Elezkurtaj, S.; Erguen, B.; et al. Inhibiting WNT and NOTCH in renal cancer stem cells and the implications for human patients. Nat. Commun. 2020, 11, 929. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.W.; Loisel-Duwattez, J.; Desterke, C.; Latsis, T.; Pagliaro, S.; Griscelli, F.; Bennaceur-Griscelli, A.; Turhan, A.G. A novel neuronal organoid model mimicking glioblastoma (GBM) features from induced pluripotent stem cells (iPSC). Biochim. Et Biophys. Acta. Gen. Subj. 2020, 1864, 129540. [Google Scholar] [CrossRef]
- Gao, M.; Lin, M.; Rao, M.; Thompson, H.; Hirai, K.; Choi, M.; Georgakis, G.V.; Sasson, A.R.; Bucobo, J.C.; Tzimas, D.; et al. Development of Patient-Derived Gastric Cancer Organoids from Endoscopic Biopsies and Surgical Tissues. Ann. Surg. Oncol. 2018, 25, 2767–2775. [Google Scholar] [CrossRef]
- Vlachogiannis, G.; Hedayat, S.; Vatsiou, A.; Jamin, Y.; Fernández-Mateos, J.; Khan, K.; Lampis, A.; Eason, K.; Huntingford, I.; Burke, R.; et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers. Science 2018, 359, 920–926. [Google Scholar] [CrossRef]
- Nanki, K.; Toshimitsu, K.; Takano, A.; Fujii, M.; Shimokawa, M.; Ohta, Y.; Matano, M.; Seino, T.; Nishikori, S.; Ishikawa, K.; et al. Divergent Routes toward Wnt and R-spondin Niche Independency during Human Gastric Carcinogenesis. Cell 2018, 174, 856–869.e817. [Google Scholar] [CrossRef]
- Steele, N.G.; Chakrabarti, J.; Wang, J.; Biesiada, J.; Holokai, L.; Chang, J.; Nowacki, L.M.; Hawkins, J.; Mahe, M.; Sundaram, N.; et al. An Organoid-Based Preclinical Model of Human Gastric Cancer. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 161–184. [Google Scholar] [CrossRef]
- Li, J.; Xu, H.; Zhang, L.; Song, L.; Feng, D.; Peng, X.; Wu, M.; Zou, Y.; Wang, B.; Zhan, L.; et al. Malignant ascites-derived organoid (MADO) cultures for gastric cancer in vitro modelling and drug screening. J. Cancer Res. Clin. Oncol. 2019, 145, 2637–2647. [Google Scholar] [CrossRef]
- Seidlitz, T.; Merker, S.R.; Rothe, A.; Zakrzewski, F.; von Neubeck, C.; Grützmann, K.; Sommer, U.; Schweitzer, C.; Schölch, S.; Uhlemann, H.; et al. Human gastric cancer modelling using organoids. Gut 2019, 68, 207–217. [Google Scholar] [CrossRef]
- Sierra, J.C.; Piazuelo, M.B.; Luis, P.B.; Barry, D.P.; Allaman, M.M.; Asim, M.; Sebrell, T.A.; Finley, J.L.; Rose, K.L.; Hill, S.; et al. Spermine oxidase mediates Helicobacter pylori-induced gastric inflammation, DNA damage, and carcinogenic signaling. Oncogene 2020, 39, 4465–4474. [Google Scholar] [CrossRef]
- Giobbe, G.G.; Bonfante, F.; Jones, B.C.; Gagliano, O.; Luni, C.; Zambaiti, E.; Perin, S.; Laterza, C.; Busslinger, G.; Stuart, H.; et al. SARS-CoV-2 infection and replication in human gastric organoids. Nat. Commun. 2021, 12, 6610. [Google Scholar] [CrossRef]
- Gobert, A.P.; Boutaud, O.; Asim, M.; Zagol-Ikapitte, I.A.; Delgado, A.G.; Latour, Y.L.; Finley, J.L.; Singh, K.; Verriere, T.G.; Allaman, M.M.; et al. Dicarbonyl Electrophiles Mediate Inflammation-Induced Gastrointestinal Carcinogenesis. Gastroenterology 2021, 160, 1256–1268.e1259. [Google Scholar] [CrossRef] [PubMed]
- Koh, V.; Chakrabarti, J.; Torvund, M.; Steele, N.; Hawkins, J.A.; Ito, Y.; Wang, J.; Helmrath, M.A.; Merchant, J.L.; Ahmed, S.A.; et al. Hedgehog transcriptional effector GLI mediates mTOR-Induced PD-L1 expression in gastric cancer organoids. Cancer Lett. 2021, 518, 59–71. [Google Scholar] [CrossRef]
- Togasaki, K.; Sugimoto, S.; Ohta, Y.; Nanki, K.; Matano, M.; Takahashi, S.; Fujii, M.; Kanai, T.; Sato, T. Wnt Signaling Shapes the Histologic Variation in Diffuse Gastric Cancer. Gastroenterology 2021, 160, 823–830. [Google Scholar] [CrossRef]
- Chakrabarti, J.; Koh, V.; So, J.B.Y.; Yong, W.P.; Zavros, Y. A Preclinical Human-Derived Autologous Gastric Cancer Organoid/Immune Cell Co-Culture Model to Predict the Efficacy of Targeted Therapies. J. Vis. Exp. JoVE 2021, 173, e61443. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.H.; Kolahi, K.S.; Du, Y.; Chang, C.Y.; Krokhotin, A.; Nair, A.; Sobba, W.D.; Karlsson, K.; Jones, S.J.; Longacre, T.A.; et al. A CRISPR/Cas9-Engineered ARID1A-Deficient Human Gastric Cancer Organoid Model Reveals Essential and Nonessential Modes of Oncogenic Transformation. Cancer Discov. 2021, 11, 1562–1581. [Google Scholar] [CrossRef] [PubMed]
- Eicher, A.K.; Kechele, D.O.; Sundaram, N.; Berns, H.M.; Poling, H.M.; Haines, L.E.; Sanchez, J.G.; Kishimoto, K.; Krishnamurthy, M.; Han, L.; et al. Functional human gastrointestinal organoids can be engineered from three primary germ layers derived separately from pluripotent stem cells. Cell Stem Cell 2022, 29, 36–51.e36. [Google Scholar] [CrossRef]
- Miao, X.; Wang, C.; Chai, C.; Tang, H.; Hu, J.; Zhao, Z.; Luo, W.; Zhang, H.; Zhu, K.; Zhou, W.; et al. Establishment of gastric cancer organoid and its application in individualized therapy. Oncol. Lett. 2022, 24, 447. [Google Scholar] [CrossRef]
- Li, G.; Ma, S.; Wu, Q.; Kong, D.; Yang, Z.; Gu, Z.; Feng, L.; Zhang, K.; Cheng, S.; Tian, Y.; et al. Establishment of gastric signet ring cell carcinoma organoid for the therapeutic drug testing. Cell Death Discov. 2022, 8, 6. [Google Scholar] [CrossRef]
- Yoon, C.; Lu, J.; Kim, B.J.; Cho, S.J.; Kim, J.H.; Moy, R.H.; Ryeom, S.W.; Yoon, S.S. Patient-Derived Organoids from Locally Advanced Gastric Adenocarcinomas Can Predict Resistance to Neoadjuvant Chemotherapy. J. Gastrointest. Surg. 2023, 27, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Qin, Y.; Jia, M.; Li, L.; Zhang, W.; Li, L.; Zhang, Z.; Liu, Y. A gastric cancer patient-derived three-dimensional cell spheroid culture model. Am. J. Cancer Res. 2023, 13, 964–975. [Google Scholar] [PubMed]
- Mazzocchi, A.; Dominijanni, A.; Soker, S. Pleural Effusion Aspirate for Use in 3D Lung Cancer Modeling and Chemotherapy Screening. Methods Mol. Biol. 2022, 2394, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.P.; Qiu, Y.; Zhang, B.; Chen, G.; Chen, Q.; Wang, M.; Mo, F.; Xu, J.; Wu, J.; Zhang, R.R.; et al. Zika virus infection induces RNAi-mediated antiviral immunity in human neural progenitors and brain organoids. Cell Res. 2019, 29, 265–273. [Google Scholar] [CrossRef]
- Li, J.; Ghazwani, M.; Liu, K.; Huang, Y.; Chang, N.; Fan, J.; He, F.; Li, L.; Bu, S.; Xie, W.; et al. Regulation of hepatic stellate cell proliferation and activation by glutamine metabolism. PLoS ONE 2017, 12, e0182679. [Google Scholar] [CrossRef]
- Sünwoldt, J.; Bosche, B.; Meisel, A.; Mergenthaler, P. Neuronal Culture Microenvironments Determine Preferences in Bioenergetic Pathway Use. Front. Mol. Neurosci. 2017, 10, 305. [Google Scholar] [CrossRef]
- Gao, K.; Zhang, T.; Wang, F.; Lv, C. Therapeutic Potential of Wnt-3a in Neurological Recovery after Spinal Cord Injury. Eur. Neurol. 2019, 81, 197–204. [Google Scholar] [CrossRef]
- Xiao, X.H.; Huang, Q.Y.; Qian, X.L.; Duan, J.; Jiao, X.Q.; Wu, L.Y.; Huang, Q.Y.; Li, J.; Lai, X.N.; Shi, Y.B.; et al. Cdc42 Promotes ADSC-Derived IPC Induction, Proliferation, And Insulin Secretion Via Wnt/β-Catenin Signaling. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2325–2339. [Google Scholar] [CrossRef]
- Lugli, N.; Kamileri, I.; Keogh, A.; Malinka, T.; Sarris, M.E.; Talianidis, I.; Schaad, O.; Candinas, D.; Stroka, D.; Halazonetis, T.D. R-spondin 1 and noggin facilitate expansion of resident stem cells from non-damaged gallbladders. EMBO Rep. 2016, 17, 769–779. [Google Scholar] [CrossRef]
- Dossena, M.; Piras, R.; Cherubini, A.; Barilani, M.; Dugnani, E.; Salanitro, F.; Moreth, T.; Pampaloni, F.; Piemonti, L.; Lazzari, L. Standardized GMP-compliant scalable production of human pancreas organoids. Stem Cell Res. Ther. 2020, 11, 94. [Google Scholar] [CrossRef]
- Nam, M.O.; Hahn, S.; Jee, J.H.; Hwang, T.S.; Yoon, H.; Lee, D.H.; Kwon, M.S.; Yoo, J. Effects of a small molecule R-spondin-1 substitute RS-246204 on a mouse intestinal organoid culture. Oncotarget 2018, 9, 6356–6368. [Google Scholar] [CrossRef]
- Xie, Y.; Park, E.S.; Xiang, D.; Li, Z. Long-term organoid culture reveals enrichment of organoid-forming epithelial cells in the fimbrial portion of mouse fallopian tube. Stem Cell Res. 2018, 32, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.R.; Koido, M.; Tadokoro, T.; Ouchi, R.; Matsuno, T.; Ueno, Y.; Sekine, K.; Takebe, T.; Taniguchi, H. Human iPSC-Derived Posterior Gut Progenitors Are Expandable and Capable of Forming Gut and Liver Organoids. Stem Cell Rep. 2018, 10, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Yip, H.Y.K.; Tan, C.W.; Hirokawa, Y.; Burgess, A.W. Colon organoid formation and cryptogenesis are stimulated by growth factors secreted from myofibroblasts. PLoS ONE 2018, 13, e0199412. [Google Scholar] [CrossRef]
- Patel, M.; Velagapudi, C.; Burns, H.; Doss, R.; Lee, M.J.; Mariappan, M.M.; Wagner, B.; Arar, M.; Barnes, V.L.; Abboud, H.E.; et al. Mouse Metanephric Mesenchymal Cell-Derived Angioblasts Undergo Vasculogenesis in Three-Dimensional Culture. Am. J. Pathol. 2018, 188, 768–784. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.K.; Odedra, D.; Chiu, L.L.; Vunjak-Novakovic, G.; Radisic, M. Vascular endothelial growth factor secretion by nonmyocytes modulates Connexin-43 levels in cardiac organoids. Tissue Eng. Part A 2012, 18, 1771–1783. [Google Scholar] [CrossRef]
- Yoshida, T.; Singh, A.K.; Bishai, W.R.; McConkey, D.J.; Bivalacqua, T.J. Organoid culture of bladder cancer cells. Investig. Clin. Urol. 2018, 59, 149–151. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Liu, B.; Wang, J.; Wei, S.; Qi, Z.; Wang, S.; Fu, W.; Chen, Y.G. A growth factor-free culture system underscores the coordination between Wnt and BMP signaling in Lgr5(+) intestinal stem cell maintenance. Cell Discov. 2018, 4, 49. [Google Scholar] [CrossRef]
- Ye, P.; Chiang, Y.J.; Qi, Z.; Li, Y.; Wang, S.; Liu, Y.; Li, X.; Chen, Y.G. Tankyrases maintain homeostasis of intestinal epithelium by preventing cell death. PLoS Genet. 2018, 14, e1007697. [Google Scholar] [CrossRef]
- Zhang, C.; Guo, H.; Yang, C.; Chen, Q.; Huang, J.; Liu, L.; Zhang, Y.; Jin, S.; Song, A.; Yang, P. The biological behavior optimization of human periodontal ligament stem cells via preconditioning by the combined application of fibroblast growth factor-2 and A83-01 in in vitro culture expansion. J. Transl. Med. 2019, 17, 66. [Google Scholar] [CrossRef]
- Gargett, C.E.; Gurung, S.; Darzi, S.; Werkmeister, J.A.; Mukherjee, S. Tissue engineering approaches for treating pelvic organ prolapse using a novel source of stem/stromal cells and new materials. Curr. Opin. Urol. 2019, 29, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Yamashita, M.; Hashita, T.; Ogami, K.; Hoshino, S.; Iwao, T.; Matsunaga, T. Efficient differentiation and purification of human induced pluripotent stem cell-derived endothelial progenitor cells and expansion with the use of inhibitors of ROCK, TGF-β, and GSK3β. Heliyon 2020, 6, e03493. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Pu, Q.; Zhang, Y.; Ma, Q.; Li, G.; Li, X. Expansion and maintenance of primary corneal epithelial stem/progenitor cells by inhibition of TGFβ receptor I-mediated signaling. Exp. Eye Res. 2019, 182, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Urbanska, A.M.; Hayakawa, Y.; Wang, H.; Au, A.S.; Luna, A.M.; Chang, W.; Jin, G.; Bhagat, G.; Abrams, J.A.; et al. Gastrin stimulates a cholecystokinin-2-receptor-expressing cardia progenitor cell and promotes progression of Barrett’s-like esophagus. Oncotarget 2017, 8, 203–214. [Google Scholar] [CrossRef]
- Lewis, M.T.; Landua, J.D.; Adams, H.C., 3rd; Medina, D. A mystery wrapped in an enigma: Matrigel enhancement of mammary cell growth and morphogenesis. J. Mammary Gland Biol. Neoplasia 2012, 17, 99–101. [Google Scholar] [CrossRef]
- Nayak, B.; Balachander, G.M.; Manjunath, S.; Rangarajan, A.; Chatterjee, K. Tissue mimetic 3D scaffold for breast tumor-derived organoid culture toward personalized chemotherapy. Colloids Surf. B Biointerfaces 2019, 180, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Balachander, G.M.; Balaji, S.A.; Rangarajan, A.; Chatterjee, K. Correction to “Enhanced Metastatic Potential in a 3D Tissue Scaffold toward a Comprehensive in Vitro Model for Breast Cancer Metastasis”. ACS Appl. Mater. Interfaces 2021, 13, 41361. [Google Scholar] [CrossRef]
- Murphy, A.R.; Laslett, A.; O’Brien, C.M.; Cameron, N.R. Scaffolds for 3D in vitro culture of neural lineage cells. Acta Biomater. 2017, 54, 1–20. [Google Scholar] [CrossRef]
- Knight, E.; Przyborski, S. Advances in 3D cell culture technologies enabling tissue-like structures to be created in vitro. J. Anat. 2015, 227, 746–756. [Google Scholar] [CrossRef]
- Saini, S.; Wick, T.M. Concentric cylinder bioreactor for production of tissue engineered cartilage: Effect of seeding density and hydrodynamic loading on construct development. Biotechnol. Prog. 2003, 19, 510–521. [Google Scholar] [CrossRef]
- DiStefano, T.J.; Chen, H.Y.; Panebianco, C.; Kaya, K.D.; Brooks, M.J.; Gieser, L.; Morgan, N.Y.; Pohida, T.; Swaroop, A. Accelerated and Improved Differentiation of Retinal Organoids from Pluripotent Stem Cells in Rotating-Wall Vessel Bioreactors. Stem Cell Rep. 2021, 16, 224. [Google Scholar] [CrossRef] [PubMed]
- Giandomenico, S.L.; Mierau, S.B.; Gibbons, G.M.; Wenger, L.M.D.; Masullo, L.; Sit, T.; Sutcliffe, M.; Boulanger, J.; Tripodi, M.; Derivery, E.; et al. Cerebral organoids at the air-liquid interface generate diverse nerve tracts with functional output. Nat. Neurosci. 2019, 22, 669–679. [Google Scholar] [CrossRef]
- Chandrasekaran, A.; Kouthouridis, S.; Lee, W.; Lin, N.; Ma, Z.; Turner, M.J.; Hanrahan, J.W.; Moraes, C. Magnetic microboats for floating, stiffness tunable, air-liquid interface epithelial cultures. Lab A Chip 2019, 19, 2786–2798. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Uroz, M.; Bays, J.L.; Chen, C.S. Harnessing Mechanobiology for Tissue Engineering. Dev. Cell 2021, 56, 180–191. [Google Scholar] [CrossRef]
- Park, S.E.; Georgescu, A.; Huh, D. Organoids-on-a-chip. Science 2019, 364, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Shirure, V.S.; Hughes, C.C.W.; George, S.C. Engineering Vascularized Organoid-on-a-Chip Models. Annu. Rev. Biomed. Eng. 2021, 23, 141–167. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, X.; Xia, X.; Han, M.; Li, F.; Li, C.; Li, Y.; Gao, D. Organoid technology for tissue engineering. J. Mol. Cell Biol. 2020, 12, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Richards, Z.; McCray, T.; Marsili, J.; Zenner, M.L.; Manlucu, J.T.; Garcia, J.; Kajdacsy-Balla, A.; Murray, M.; Voisine, C.; Murphy, A.B.; et al. Prostate Stroma Increases the Viability and Maintains the Branching Phenotype of Human Prostate Organoids. IScience 2019, 12, 304–317. [Google Scholar] [CrossRef]
- Bredenoord, A.L.; Clevers, H.; Knoblich, J.A. Human tissues in a dish: The research and ethical implications of organoid technology. Science 2017, 355, eaaf9414. [Google Scholar] [CrossRef]
- Mansour, A.A.; Gonçalves, J.T.; Bloyd, C.W.; Li, H.; Fernandes, S.; Quang, D.; Johnston, S.; Parylak, S.L.; Jin, X.; Gage, F.H. An in vivo model of functional and vascularized human brain organoids. Nat. Biotechnol. 2018, 36, 432–441. [Google Scholar] [CrossRef]
- Low, J.H.; Li, P.; Chew, E.G.Y.; Zhou, B.; Suzuki, K.; Zhang, T.; Lian, M.M.; Liu, M.; Aizawa, E.; Rodriguez Esteban, C.; et al. Generation of Human PSC-Derived Kidney Organoids with Patterned Nephron Segments and a De Novo Vascular Network. Cell Stem Cell 2019, 25, 373–387.e379. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Tan, Z.; Su, Y.; Liu, J.; Chang, M.; Yan, F.; Chen, J.; Chen, T.; Li, C.; et al. Human ESC-derived expandable hepatic organoids enable therapeutic liver repopulation and pathophysiological modeling of alcoholic liver injury. Cell Res. 2019, 29, 1009–1026. [Google Scholar] [CrossRef] [PubMed]
- Baldan, J.; Houbracken, I.; Rooman, I.; Bouwens, L. Adult human pancreatic acinar cells dedifferentiate into an embryonic progenitor-like state in 3D suspension culture. Sci. Rep. 2019, 9, 4040. [Google Scholar] [CrossRef] [PubMed]
- Bartfeld, S.; Bayram, T.; van de Wetering, M.; Huch, M.; Begthel, H.; Kujala, P.; Vries, R.; Peters, P.J.; Clevers, H. In vitro expansion of human gastric epithelial stem cells and their responses to bacterial infection. Gastroenterology 2015, 148, 126–136.e126. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.K.; Kurisaki, A. Formation of Stomach Tissue by Organoid Culture Using Mouse Embryonic Stem Cells. Methods Mol. Biol. 2017, 1597, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Broda, T.R.; McCracken, K.W.; Wells, J.M. Generation of human antral and fundic gastric organoids from pluripotent stem cells. Nat. Protoc. 2019, 14, 28–50. [Google Scholar] [CrossRef]
- Jee, J.; Park, J.H.; Im, J.H.; Kim, M.S.; Park, E.; Lim, T.; Choi, W.H.; Kim, J.H.; Kim, W.R.; Ko, J.S.; et al. Functional recovery by colon organoid transplantation in a mouse model of radiation proctitis. Biomaterials 2021, 275, 120925. [Google Scholar] [CrossRef]
- Sampaziotis, F.; Muraro, D.; Tysoe, O.C.; Sawiak, S.; Beach, T.E.; Godfrey, E.M.; Upponi, S.S.; Brevini, T.; Wesley, B.T.; Garcia-Bernardo, J.; et al. Cholangiocyte organoids can repair bile ducts after transplantation in the human liver. Science 2021, 371, 839–846. [Google Scholar] [CrossRef]
- Workman, M.J.; Mahe, M.M.; Trisno, S.; Poling, H.M.; Watson, C.L.; Sundaram, N.; Chang, C.F.; Schiesser, J.; Aubert, P.; Stanley, E.G.; et al. Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system. Nat. Med. 2017, 23, 49–59. [Google Scholar] [CrossRef]
- Múnera, J.O.; Wells, J.M. Generation of Gastrointestinal Organoids from Human Pluripotent Stem Cells. Methods Mol. Biol. 2017, 1597, 167–177. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Zeng, X.L.; Utama, B.; Atmar, R.L.; Shroyer, N.F.; Estes, M.K. Stem cell-derived human intestinal organoids as an infection model for rotaviruses. mBio 2012, 3, e00159-12. [Google Scholar] [CrossRef] [PubMed]
- Forbester, J.L.; Goulding, D.; Vallier, L.; Hannan, N.; Hale, C.; Pickard, D.; Mukhopadhyay, S.; Dougan, G. Interaction of Salmonella enterica Serovar Typhimurium with Intestinal Organoids Derived from Human Induced Pluripotent Stem Cells. Infect. Immun. 2015, 83, 2926–2934. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.L.; Huang, S.; Opp, J.S.; Nagy, M.S.; Kobayashi, M.; Young, V.B.; Spence, J.R. Persistence and toxin production by Clostridium difficile within human intestinal organoids result in disruption of epithelial paracellular barrier function. Infect. Immun. 2015, 83, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Salama, N.R.; Hartung, M.L.; Müller, A. Life in the human stomach: Persistence strategies of the bacterial pathogen Helicobacter pylori. Nat. Rev. Microbiol. 2013, 11, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Wuputra, K.; Ku, C.C.; Kato, K.; Wu, D.C.; Saito, S.; Yokoyama, K.K. Translational models of 3-D organoids and cancer stem cells in gastric cancer research. Stem Cell Res. Ther. 2021, 12, 492. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Smolka, A.J. Gastric Parietal Cell Physiology and Helicobacter pylori-Induced Disease. Gastroenterology 2019, 156, 2158–2173. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, J.L.; Lee, A. Animal models of Helicobacter pylori infection and disease. Microbes Infect. 2003, 5, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Schlaermann, P.; Toelle, B.; Berger, H.; Schmidt, S.C.; Glanemann, M.; Ordemann, J.; Bartfeld, S.; Mollenkopf, H.J.; Meyer, T.F. A novel human gastric primary cell culture system for modelling Helicobacter pylori infection in vitro. Gut 2016, 65, 202–213. [Google Scholar] [CrossRef] [PubMed]
- McCracken, K.W.; Catá, E.M.; Crawford, C.M.; Sinagoga, K.L.; Schumacher, M.; Rockich, B.E.; Tsai, Y.H.; Mayhew, C.N.; Spence, J.R.; Zavros, Y.; et al. Modelling human development and disease in pluripotent stem-cell-derived gastric organoids. Nature 2014, 516, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.C.; Evans, T.; Chen, S. Modeling endodermal organ development and diseases using human pluripotent stem cell-derived organoids. J. Mol. Cell Biol. 2020, 12, 580–592. [Google Scholar] [CrossRef]
- Bertaux-Skeirik, N.; Feng, R.; Schumacher, M.A.; Li, J.; Mahe, M.M.; Engevik, A.C.; Javier, J.E.; Peek, R.M., Jr.; Ottemann, K.; Orian-Rousseau, V.; et al. CD44 plays a functional role in Helicobacter pylori-induced epithelial cell proliferation. PLoS Pathog. 2015, 11, e1004663. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Sweeney, E.G.; Sigal, M.; Zhang, H.C.; Remington, S.J.; Cantrell, M.A.; Kuo, C.J.; Guillemin, K.; Amieva, M.R. Chemodetection and Destruction of Host Urea Allows Helicobacter pylori to Locate the Epithelium. Cell Host Microbe 2015, 18, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Holokai, L.; Chakrabarti, J.; Broda, T.; Chang, J.; Hawkins, J.A.; Sundaram, N.; Wroblewski, L.E.; Peek, R.M., Jr.; Wang, J.; Helmrath, M.; et al. Increased Programmed Death-Ligand 1 is an Early Epithelial Cell Response to Helicobacter pylori Infection. PLoS Pathog. 2019, 15, e1007468. [Google Scholar] [CrossRef] [PubMed]
- Sebrell, T.A.; Hashimi, M.; Sidar, B.; Wilkinson, R.A.; Kirpotina, L.; Quinn, M.T.; Malkoç, Z.; Taylor, P.J.; Wilking, J.N.; Bimczok, D. A Novel Gastric Spheroid Co-culture Model Reveals Chemokine-Dependent Recruitment of Human Dendritic Cells to the Gastric Epithelium. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 157–171.e153. [Google Scholar] [CrossRef] [PubMed]
- Seidlitz, T.; Koo, B.K.; Stange, D.E. Gastric organoids-an in vitro model system for the study of gastric development and road to personalized medicine. Cell Death Differ. 2021, 28, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, K. Cancer research using organoid technology. J. Mol. Med. 2021, 99, 501–515. [Google Scholar] [CrossRef]
- Pang, M.J.; Burclaff, J.R.; Jin, R.; Adkins-Threats, M.; Osaki, L.H.; Han, Y.; Mills, J.C.; Miao, Z.F.; Wang, Z.N. Gastric Organoids: Progress and Remaining Challenges. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 19–33. [Google Scholar] [CrossRef]
- Wallaschek, N.; Niklas, C.; Pompaiah, M.; Wiegering, A.; Germer, C.T.; Kircher, S.; Brändlein, S.; Maurus, K.; Rosenwald, A.; Yan, H.H.N.; et al. Establishing Pure Cancer Organoid Cultures: Identification, Selection and Verification of Cancer Phenotypes and Genotypes. J. Mol. Biol. 2019, 431, 2884–2893. [Google Scholar] [CrossRef]
- Garreta, E.; Kamm, R.D.; Chuva de Sousa Lopes, S.M.; Lancaster, M.A.; Weiss, R.; Trepat, X.; Hyun, I.; Montserrat, N. Rethinking organoid technology through bioengineering. Nat. Mater. 2021, 20, 145–155. [Google Scholar] [CrossRef]
- Velasco, S.; Kedaigle, A.J.; Simmons, S.K.; Nash, A.; Rocha, M.; Quadrato, G.; Paulsen, B.; Nguyen, L.; Adiconis, X.; Regev, A.; et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 2019, 570, 523–527. [Google Scholar] [CrossRef]
- Rossi, G.; Manfrin, A.; Lutolf, M.P. Progress and potential in organoid research. Nat. Rev. Genet. 2018, 19, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Seong, H.; Li, X.; Yu, X.; Xu, S.; Li, Y. Human Brain Organoid: A Versatile Tool for Modeling Neurodegeneration Diseases and for Drug Screening. Stem Cells Int. 2022, 2022, 2150680. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Yeleswarapu, S.; Heinrich, M.A.; Prakash, J.; Pati, F. Mimicking tumor microenvironment by 3D bioprinting: 3D cancer modeling. Biofabrication 2022, 14, 032002. [Google Scholar] [CrossRef] [PubMed]
- O’Hara-Wright, M.; Mobini, S.; Gonzalez-Cordero, A. Bioelectric Potential in Next-Generation Organoids: Electrical Stimulation to Enhance 3D Structures of the Central Nervous System. Front. Cell Dev. Biol. 2022, 10, 901652. [Google Scholar] [CrossRef] [PubMed]
- Tuveson, D.; Clevers, H. Cancer modeling meets human organoid technology. Science 2019, 364, 952–955. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.C.H.; Kranenburg, O.; Xiao, H.; Yu, J. Organoid models of gastrointestinal cancers in basic and translational research. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 203–222. [Google Scholar] [CrossRef]


{kind=link}
| iPSC-Derived | ASC-Derived | |
|---|---|---|
| Cell types | Epithelial and mesenchymal cells | Epithelial cells only |
| Proliferative capacity in vitro | Multiple passages | Unlimited multiplication |
| Modelling time | 4–8 weeks | 1–2 weeks |
| Histomorphological features | Resembling embryonic tissue | Resembling adult tissue |
| Year | Authors | Tissue Acquisition | Stem Cells | Methods of Isolating Glands | Applications |
|---|---|---|---|---|---|
| 2018 | Gao M et al. [15] | Endoscope Surgery | ASC | Physical pressure | Next-generation sequencing Drug testing |
| 2018 | Vlachogiannis G et al. [16] | Biopsy | ASC | TrypLE | Whole genome sequencing Drug testing Targeted therapy testing Biobank establishment |
| 2018 | Nanki K et al. [17] | Surgery Endoscope Ascites puncture | ASC | Libase TH TrypLE | CRISPR/Cas9 Transcriptomic analysis Xenotransplantation of organoids Biobank establishment |
| 2019 | Steele NG et al. [18] | Surgery | ASC | Collagenase Hyaluronidase | RNA sequencing Drug testing Targeted therapy testing Organoids tumorigenicity analysis |
| 2019 | Li J et al. [19] | Ascites puncture | ASC | - | Malignant ascites-derived organoid Drug testing |
| 2019 | Seidlitz T et al. [20] | Surgery | ASC | Collagenase XI Dispase II | Whole genome sequencing RNA sequencing Drug testing Targeted therapy testing Biobank establishment |
| 2020 | Sierra JC et al. [21] | Surgery | ASC | Collagenase IV | H. pylori |
| 2021 | Giobbe GG. et al. [22] | Biopsy | ASC | Physical pressure | SARS-CoV-2 infection |
| 2021 | Gobert AP et al. [23] | Endoscope | ASC | - | H. pylori |
| 2021 | Koh V et al. [24] | Surgery | ASC | Collagenase | Immunoregulation in EMT |
| 2021 | Togasaki K et al. [25] | Surgery Ascites puncture | ASC | Libase TH | Whole exome sequence RNA sequence Xenotransplantation of organoids |
| 2021 | Chakrabarti J et al. [26] | Biopsy | ASC | Collagenase I Hyaluronidase IV-S | Organoid/immune cells co-culture model Targeted therapy testing |
| 2021 | Lo YH et al. [27] | Surgery | ASC | Collagenase I | CRISPR/Cas9 |
| 2022 | Eicher AK et al. [28] | - | hPSC | - | Organoid assembly approach Tissue engineering |
| 2022 | Miao X et al. [29] | Surgery | ASC | Collagenase II Dispase II | Drug testing |
| 2022 | Li G et al. [30] | Surgery | ASC | Collagenase | Organoids tumorigenicity analysis Drug testing |
| 2023 | Yoon C et al. [31] | Endoscope | ASC | Collagenase III | Drug testing |
| 2023 | Zhang H et al. [32] | Surgery | ASC | Gentle MACS™ Dissociator tumor dissociation kit | Transcriptome sequencing Drug testing |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, C.; Zhang, X.; Gu, Y.; Wang, D.; Zhang, S.; Liu, T.; Li, Y.; He, W. Organoids: Construction and Application in Gastric Cancer. Biomolecules 2023, 13, 875. https://doi.org/10.3390/biom13050875
Huo C, Zhang X, Gu Y, Wang D, Zhang S, Liu T, Li Y, He W. Organoids: Construction and Application in Gastric Cancer. Biomolecules. 2023; 13(5):875. https://doi.org/10.3390/biom13050875
Chicago/Turabian StyleHuo, Chengdong, Xiaoxia Zhang, Yanmei Gu, Daijun Wang, Shining Zhang, Tao Liu, Yumin Li, and Wenting He. 2023. "Organoids: Construction and Application in Gastric Cancer" Biomolecules 13, no. 5: 875. https://doi.org/10.3390/biom13050875