
Unexpected Distribution of Chitin and Chitin Synthase across Soft-Bodied Cnidarians



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Collection and Sample Preparation
2.2. Description of Chitin-Binding Domain Peptide Probe
2.3. Tissue Embedding and Sectioning
2.4. Chitinase Treatment
2.5. Chitin Affinity Histochemistry
2.6. Semi-Quantitative PCR, Probe Synthesis, In Situ Hybridization
2.7. Bioinformatics Pipeline
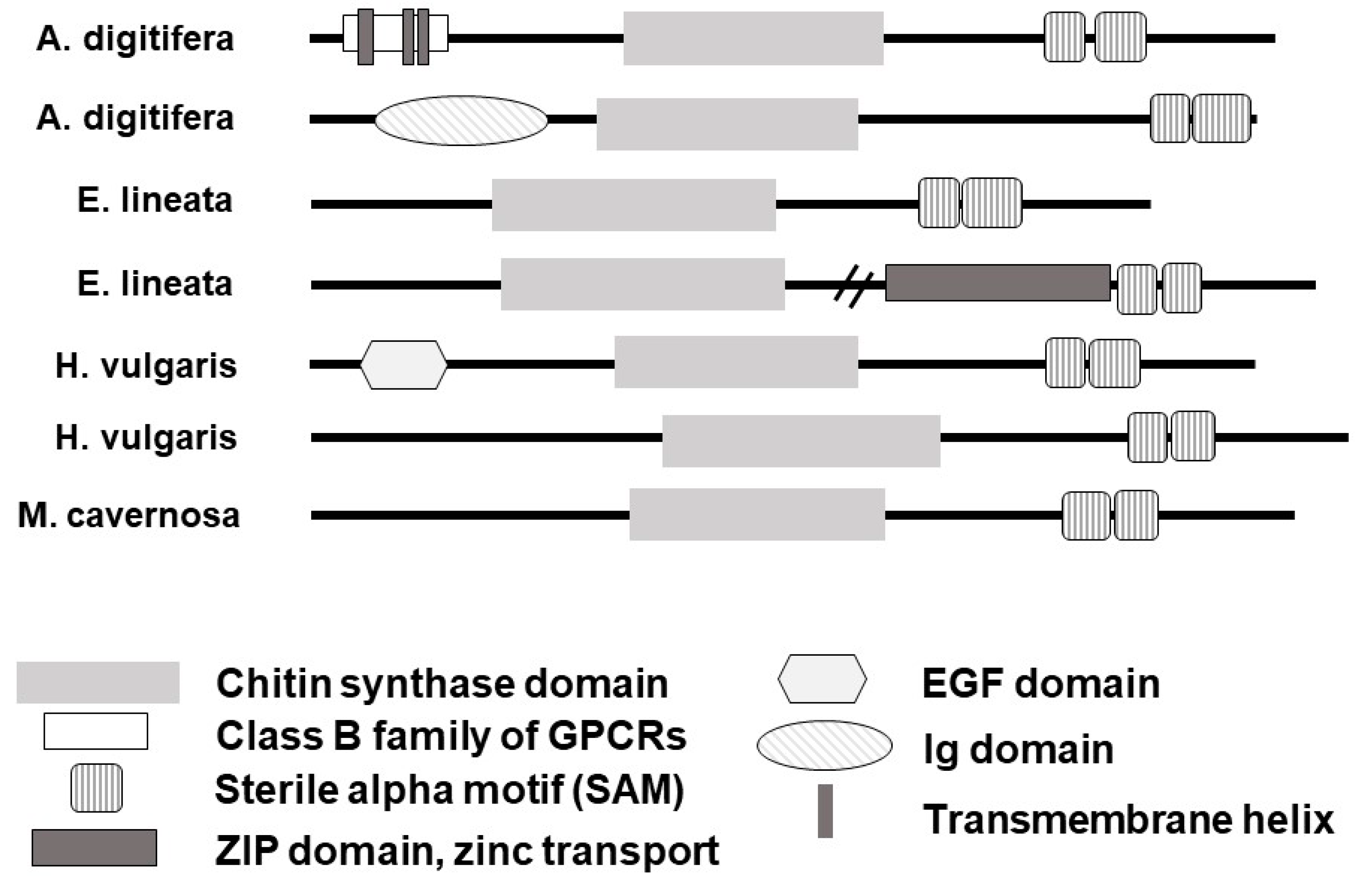
2.7.1. Identification of CHS Homologs
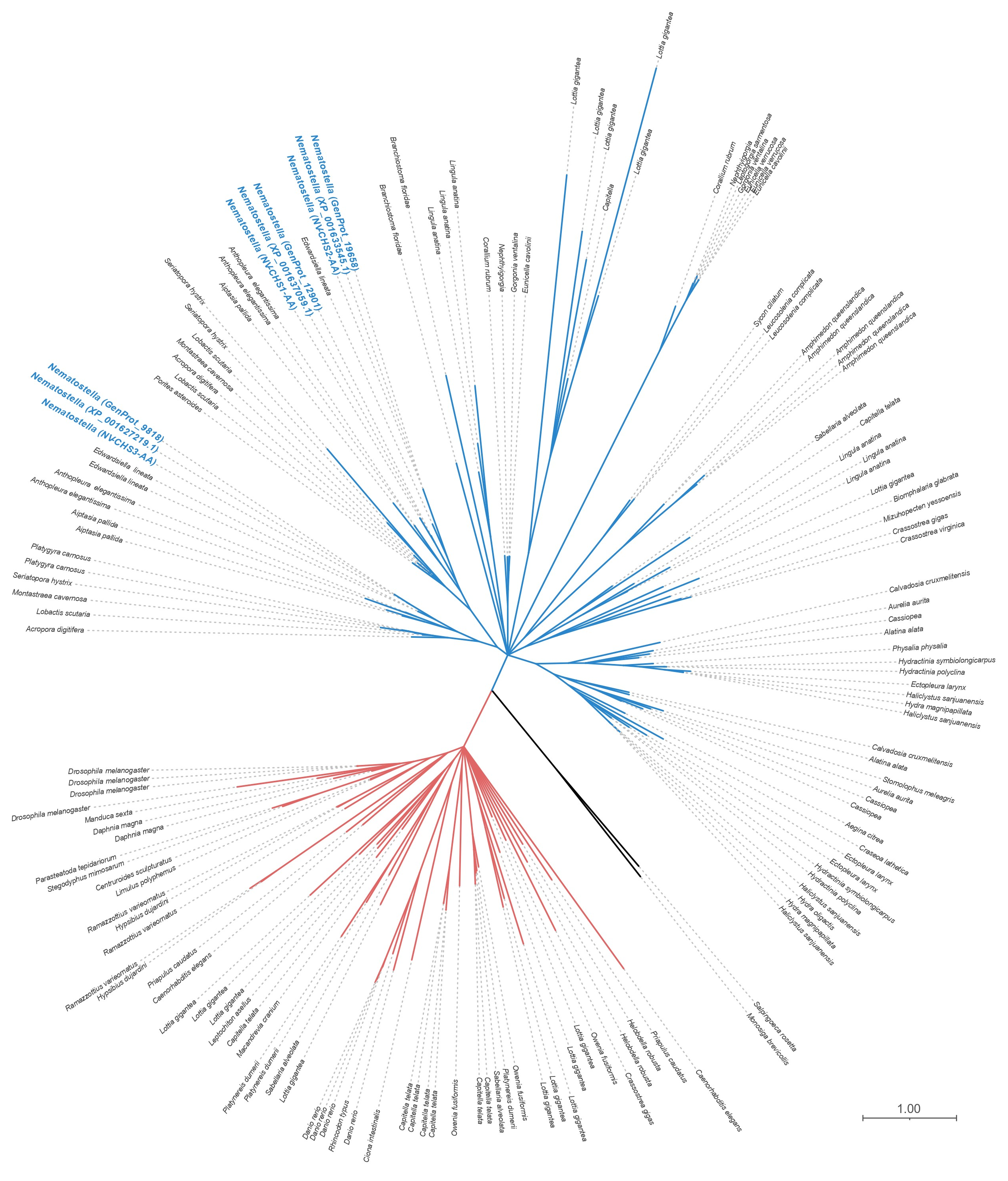
2.7.2. Phylogenetic Analysis
3. Results
3.1. Predicted Homologs for the Enzyme Chitin Synthase (CHS) Are Present in Most Recognized Cnidarian Clades and Expressed in Taxa or Life Stages with No Reported Rigid Structures
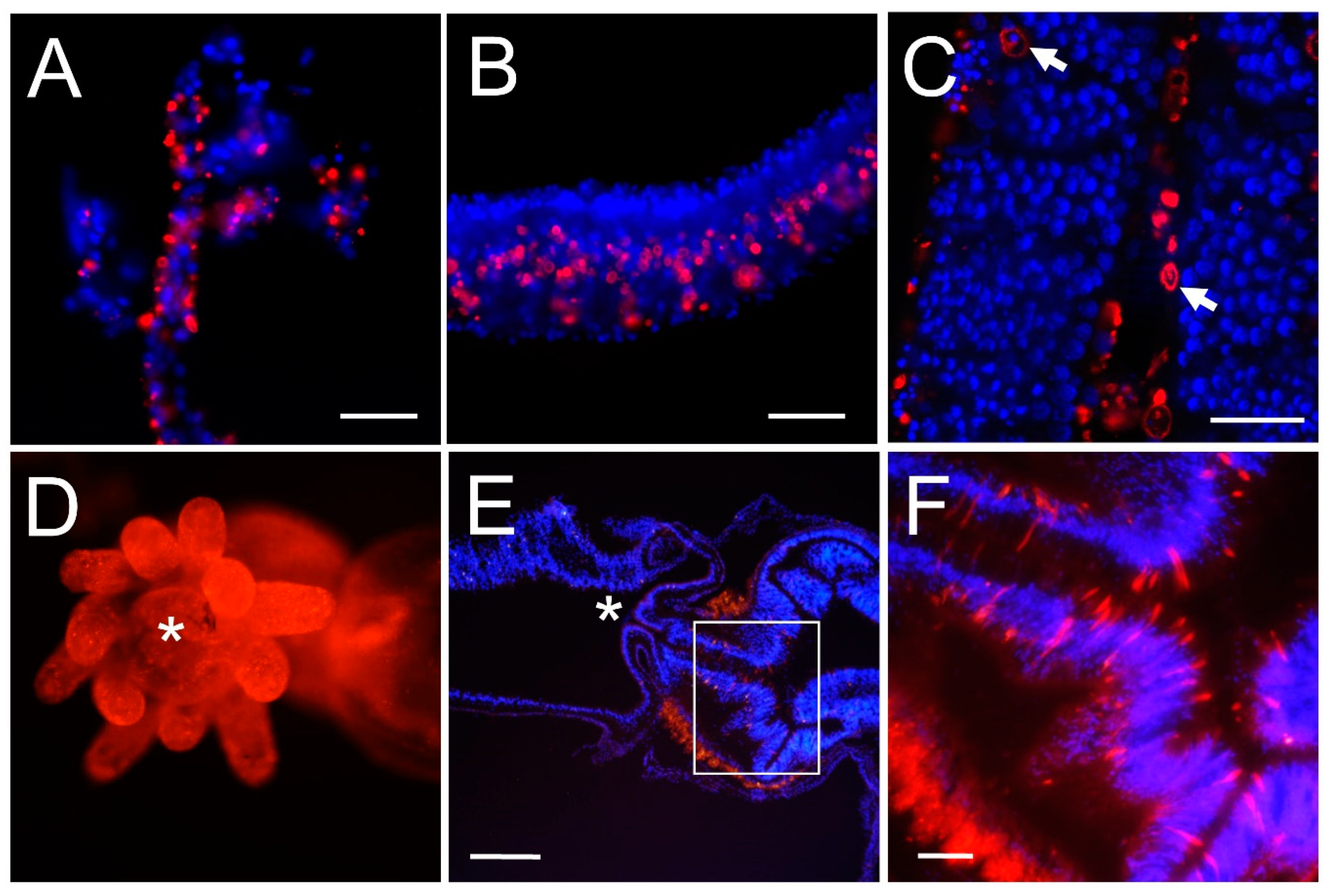
3.2. Chitin Is Present in Cnidarian Soft Tissues
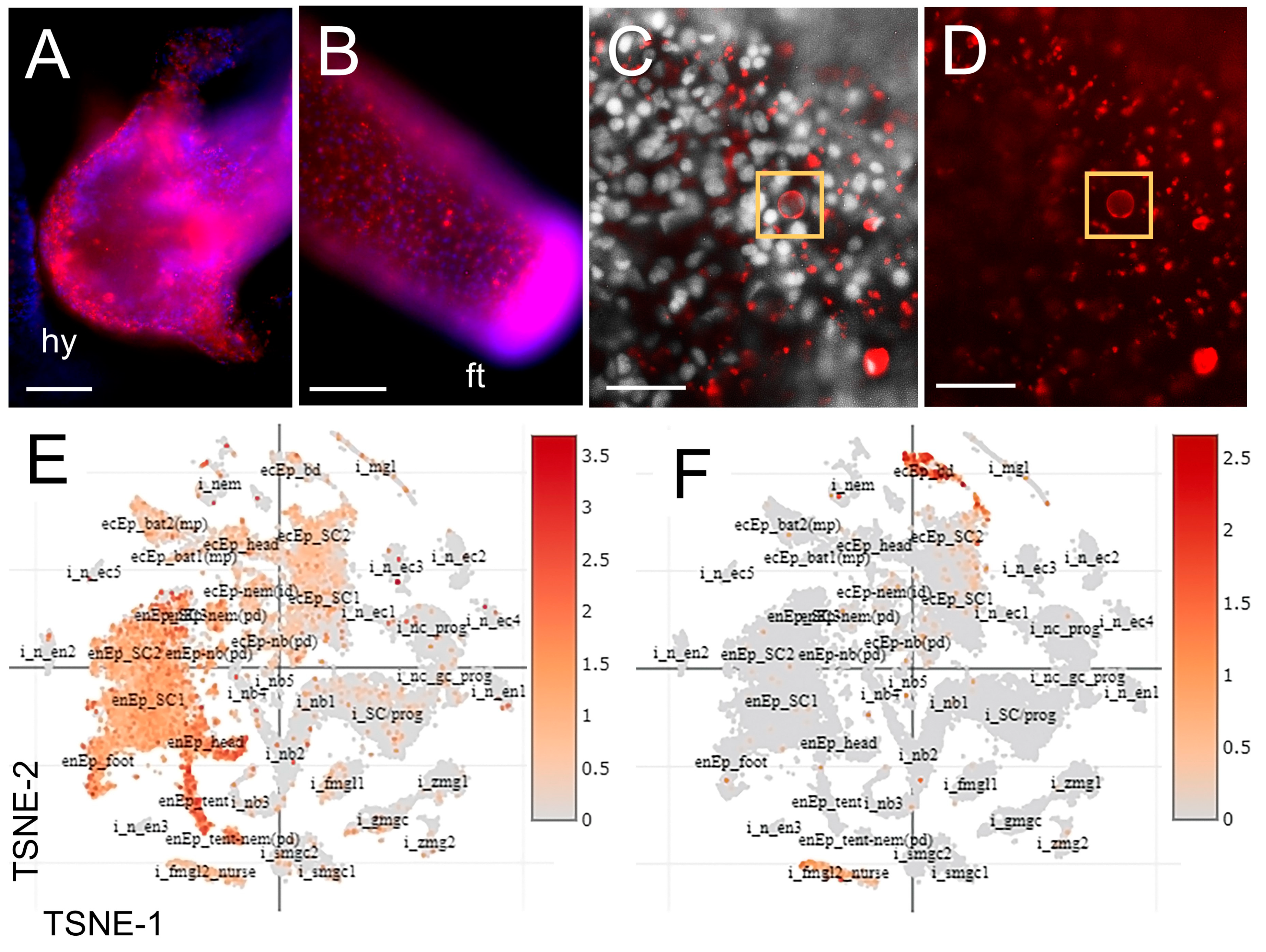
3.3. The Distribution of Chitin and Expression of Chitin Synthases in Hydra
3.4. Chitin Synthase Genes Are Differentially Expressed in the Model Sea Anemone Nematostella vectensis during Development
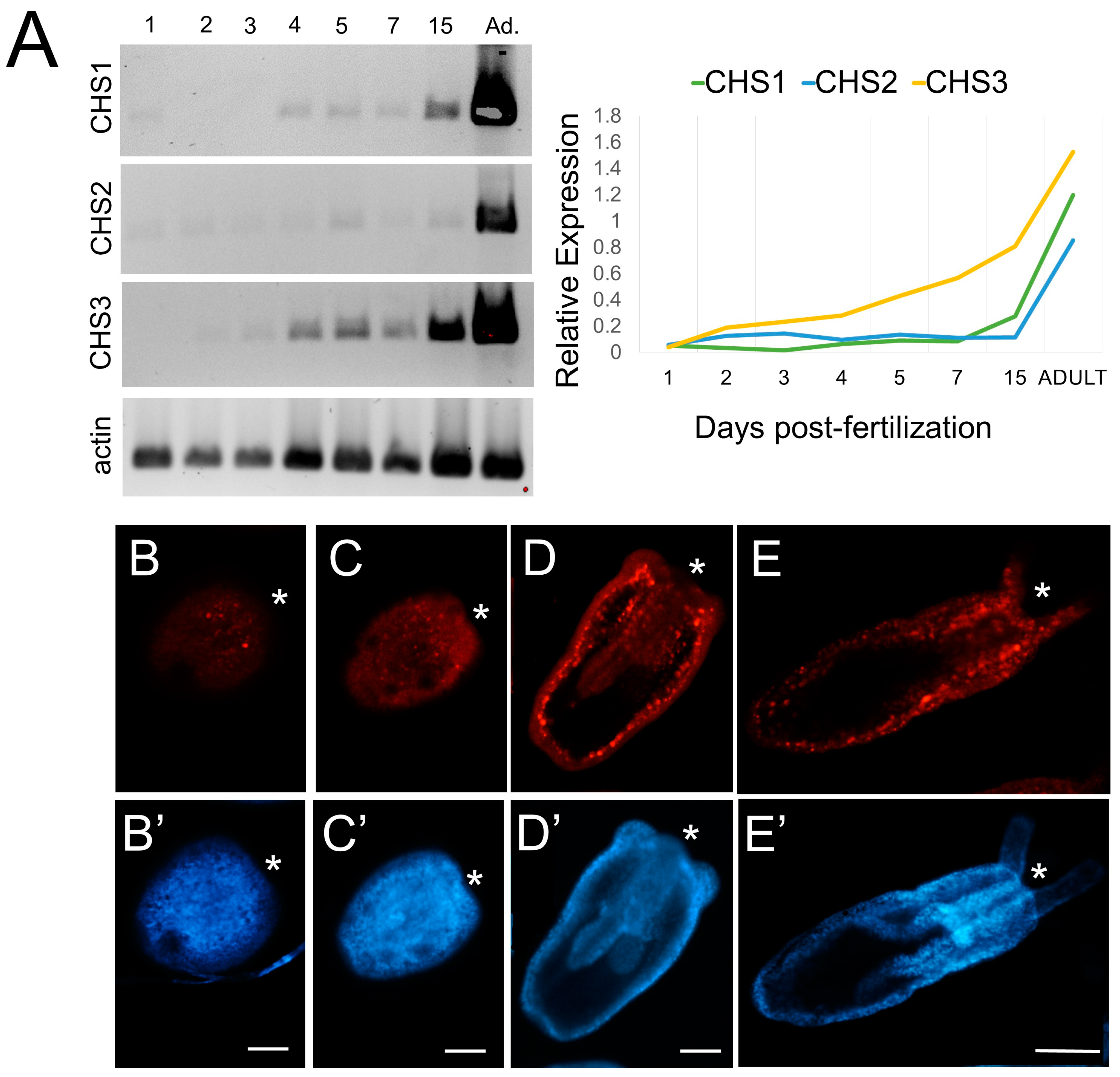
3.4.1. Three Nematostella CHS Paralogs Are Expressed during Development
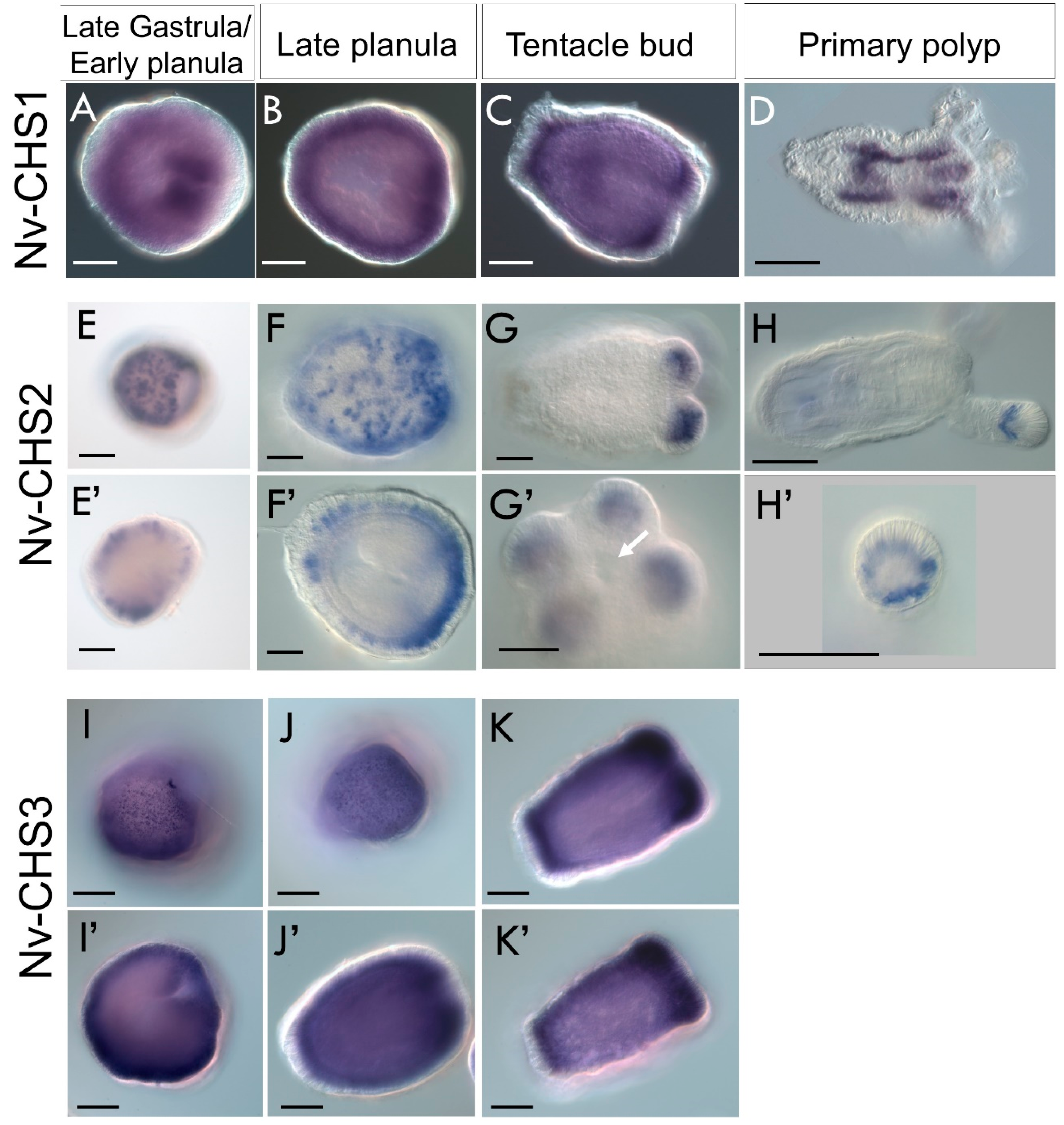
3.4.2. Chitin Is Distributed throughout the Developing Planula and Primary Polyp
3.4.3. Chitin Synthases Are Differentially Expressed in the Ectoderm during Nematostella Development
4. Discussion
4.1. The Molecular Toolkit for Chitin Synthesis Is Present in More Cnidarian Taxa Than Previously Reported, including in Soft-Bodied Species or Life-History Stages
4.2. Distribution of Chitin in Cnidarian Tissues
4.3. CHS Is Expressed during Nematostella vectensis Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, S.-K. Chitin, Chitosan, Oligosaccharides and Their Derivatives: Biological Activities and Applications, 1st ed.; Kim, S.-K., Ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 666. [Google Scholar]
- Zakrzewski, A.C.; Weigert, A.; Helm, C.; Adamski, M.; Adamska, M.; Bleidorn, C.; Raible, F.; Hausen, H. Early divergence, broad distribution, and high diversity of animal chitin synthases. Genome Biol. Evol. 2014, 6, 316–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, J.P.; Falomir, M.P.; Gozalbo, D. Chitin: A Structural Biopolysaccharide with Multiple Applications. eLS 2014. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.; Zhu, K.Y. Chitin in Arthropods: Biosynthesis, Modification, and Metabolism. Adv. Exp. Med. Biol. 2019, 1142, 169–207. [Google Scholar] [CrossRef]
- Krings, W.; Brütt, J.-O.; Gorb, S.; Glaubrecht, M. Tightening it Up: Diversity of the Chitin Anchorage of Radular-Teeth in Paludomid Freshwater-Gastropods. Malacologia 2020, 63, 77–94. [Google Scholar] [CrossRef]
- Tilic, E.; Bartolomaeus, T. Structure, function and cell dynamics during chaetogenesis of abdominal uncini in Sabellaria alveolata (Sabellariidae, Annelida). Zool. Lett. 2016, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.J.; Fernandez, J.G.; Sohn, J.J.; Amemiya, C.T. Chitin is endogenously produced in vertebrates. Curr. Biol. 2015, 25, 897–900. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.; Tang, W.J.; Robinson, M.; Daza, D.O.; Hassan, K.; Leppert, V.; Hirst, L.S.; Amemiya, C.T. Evidence of chitin in the ampullae of Lorenzini of chondrichthyan fishes. Curr. Biol. 2020, 30, R1254–R1255. [Google Scholar] [CrossRef]
- Phillips, M.; Wheeler, A.C.; Robinson, M.J.; Leppert, V.; Jia, M.; Rolandi, M.; Hirst, L.S.; Amemiya, C.T. Colloidal structure and proton conductivity of the gel within the electrosensory organs of cartilaginous fishes. iScience 2021, 24, 102947. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Hild, S.; Walther, P.; PLoSs, K.; Boland, W.; Tomaschko, K.H. Granular chitin in the epidermis of nudibranch molluscs. Biol. Bull. 2007, 213, 307–315. [Google Scholar] [CrossRef]
- Gonçalves, I.R.; Brouillet, S.; Soulié, M.-C.; Gribaldo, S.; Sirven, C.; Charron, N.; Boccara, M.; Choquer, M. Genome-wide analyses of chitin synthases identify horizontal gene transfers towards bacteria and allow a robust and unifying classification into fungi. BMC Evol. Biol. 2016, 16, 252. [Google Scholar] [CrossRef] [Green Version]
- Morozov, A.A.; Likhoshway, Y.V. Evolutionary history of the chitin synthases of eukaryotes. Glycobiology 2016, 26, 635–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Cao, P.; Liu, Y.; Yu, A.; Wang, D.; Chen, L.; Sundarraj, R.; Yuchi, Z.; Gong, Y.; Merzendorfer, H.; et al. Structural basis for directional chitin biosynthesis. Nature 2022, 610, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Klinter, S.; Bulone, V.; Arvestad, L. Diversity and evolution of chitin synthases in oomycetes (Straminipila: Oomycota). Mol. Phylogenet. Evol. 2019, 139, 106558. [Google Scholar] [CrossRef]
- Stapleton, D.; Balan, I.; Pawson, T.; Sicheri, F. The crystal structure of an Eph receptor SAM domain reveals a mechanism for modular dimerization. Nat. Struct. Biol. 1999, 6, 44–49. [Google Scholar] [CrossRef]
- Ray, S.; Chee, L.; Matson, D.R.; Palermo, N.Y.; Bresnick, E.H.; Hewitt, K.J. Sterile α-motif domain requirement for cellular signaling and survival. J. Biol. Chem. 2020, 295, 7113–7125. [Google Scholar] [CrossRef] [Green Version]
- Guerriero, G. Putative chitin synthases from Branchiostoma floridae Show Extracellular Matrix-related Domains and Mosaic Structures. Genom. Proteom. Bioinform. 2012, 10, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, C.W.; Giribet, G.; Edgecombe, G.D.; Hejnol, A. Animal Phylogeny and Its Evolutionary Implications. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 371–395. [Google Scholar] [CrossRef] [Green Version]
- Whelan, N.V.; Kocot, K.M.; Moroz, T.P.; Mukherjee, K.; Williams, P.; Paulay, G.; Moroz, L.L.; Halanych, K.M. Ctenophore relationships and their placement as the sister group to all other animals. Nat. Ecol. Evol. 2017, 1, 1737–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laumer, C.E.; Fernandez, R.; Lemer, S.; Combosch, D.; Kocot, K.M.; Riesgo, A.; Andrade, S.C.S.; Sterrer, W.; Sorensen, M.V.; Giribet, G. Revisiting metazoan phylogeny with genomic sampling of all phyla. Proc. Biol. Sci./R. Soc. 2019, 286, 20190831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
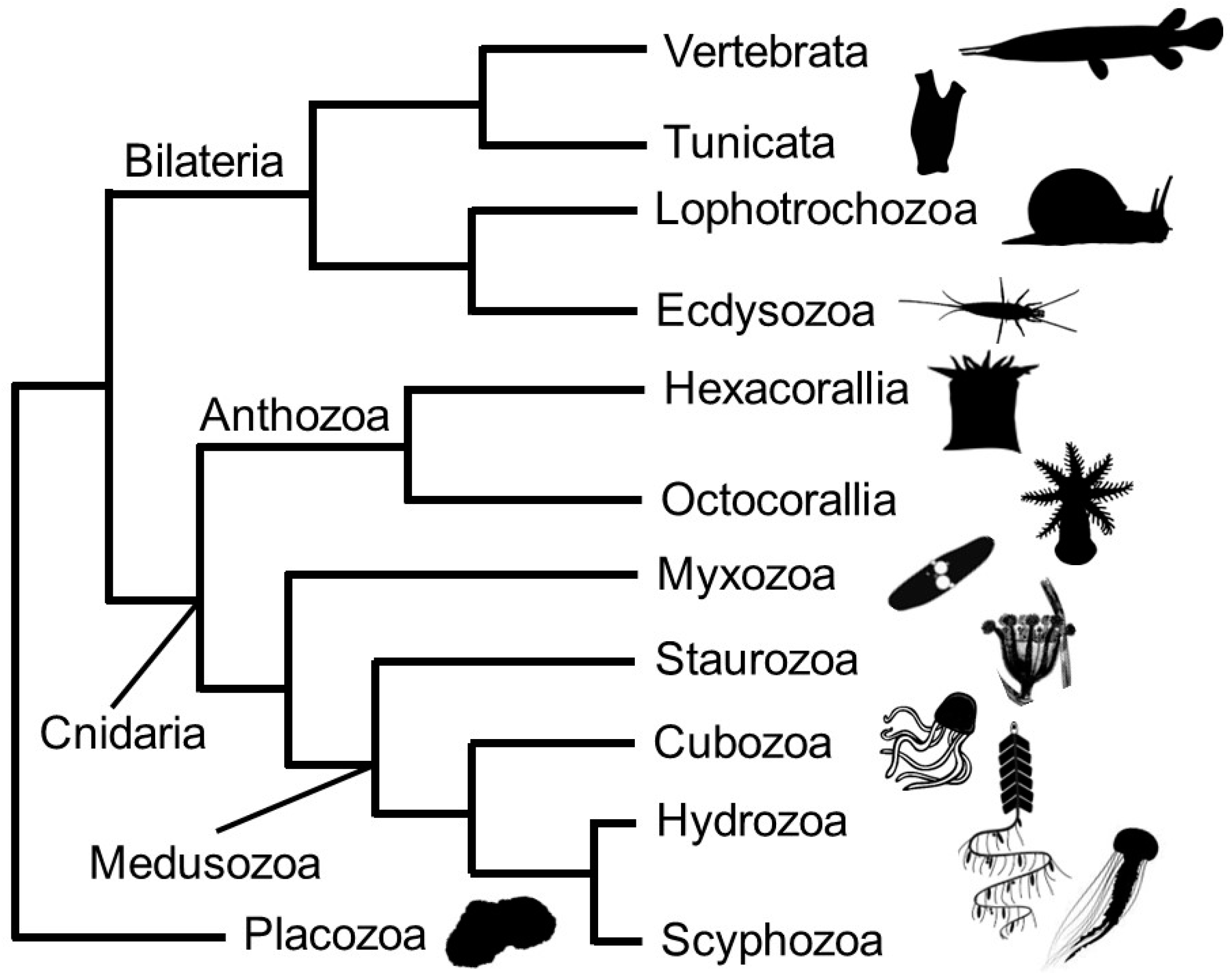
- Daly, M.; Brugler, M.R.; Cartwright, P.; Collins, A.G.; Dawson, M.N.; Fautin, D.G.; McFadden, C.S.; Opresko, D.M.; Rodríguez, E.; Romano, S.L.; et al. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 2007, 1668, 127–182. [Google Scholar] [CrossRef] [Green Version]
- Babonis, L.S.; Martindale, M.Q.; Ryan, J.F. Do novel genes drive morphological novelty? An investigation of the nematosomes in the sea anemone Nematostella vectensis. BMC Evol. Biol. 2016, 16, 114. [Google Scholar] [CrossRef] [Green Version]
- Muller, W.A.; Plickert, G. Quantitative analysis of an inhibitory gradient field in the hydrozoan stolon. Wilehm Roux Arch. Dev. Biol. 1982, 191, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Lange, R.G.; Muller, W.A. SIF, a novel morphogenetic inducer in hydrozoa. Dev. Biol. 1991, 147, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Bo, M.; Bavestrello, G.; Kurek, D.; Paasch, S.; Brunner, E.; Born, R.; Galli, R.; Stelling, A.L.; Sivkov, V.N.; Petrova, O.V.; et al. Isolation and identification of chitin in the black coral Parantipathes larix (Anthozoa: Cnidaria). Int. J. Biol. Macromol. 2012, 51, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zapata, F.; Goetz, F.E.; Smith, S.A.; Howison, M.; Siebert, S.; Church, S.H.; Sanders, S.M.; Ames, C.L.; McFadden, C.S.; France, S.C.; et al. Phylogenomic Analyses Support Traditional Relationships within Cnidaria. PLoS ONE 2015, 10, e0139068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moneer, J.; Siebert, S.; Krebs, S.; Cazet, J.; Prexl, A.; Pan, Q.; Juliano, C.; Bottger, A. Differential gene regulation in DAPT-treated Hydra reveals candidate direct Notch signalling targets. J. Cell Sci. 2021, 134, jcs258768. [Google Scholar] [CrossRef]
- Stefanik, D.J.; Friedman, L.E.; Finnerty, J.R. Collecting, rearing, spawning and inducing regeneration of the starlet sea anemone, Nematostella vectensis. Nat. Protoc. 2013, 8, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Foster, J.M.; Nelson, L.S.; Ma, D.; Carlow, C.K.S. The chitin synthase genes chs-1 and chs-2 are essential for C. elegans development and responsible for chitin deposition in the eggshell and pharynx, respectively. Dev. Biol. 2005, 285, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Mandel Mark, J.; Schaefer Amy, L.; Brennan Caitlin, A.; Heath-Heckman Elizabeth, A.C.; DeLoney-Marino Cindy, R.; McFall-Ngai Margaret, J.; Ruby Edward, G. Squid-Derived Chitin Oligosaccharides Are a Chemotactic Signal during Colonization by Vibrio fischeri. Appl. Environ. Microbiol. 2012, 78, 4620–4626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- JayaNandanan, N.; Mathew, R.; Leptin, M. Guidance of subcellular tubulogenesis by actin under the control of a synaptotagmin-like protein and Moesin. Nat. Commun. 2014, 5, 3036. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.; Matus, D.Q.; Martindale, M.Q. The ancestral role of COE genes may have been in chemoreception: Evidence from the development of the sea anemone, Nematostella vectensis (Phylum Cnidaria; Class Anthozoa). Dev. Genes Evol. 2004, 214, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Servetnick, M.D.; Steinworth, B.; Babonis, L.S.; Simmons, D.; Salinas-Saavedra, M.; Martindale, M.Q. Cas9-mediated excision of Nematostella brachyury disrupts endoderm development, pharynx formation and oral-aboral patterning. Development 2017, 144, 2951–2960. [Google Scholar] [CrossRef] [Green Version]
- Kayal, E.; Bentlage, B.; Sabrina Pankey, M.; Ohdera, A.H.; Medina, M.; Plachetzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 68. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. A new generation of homology search tools based on probabilistic inference. Genome Inf. 2009, 23, 205–211. [Google Scholar]
- Specht, C.A.; Liu, Y.; Robbins, P.W.; Bulawa, C.E.; Iartchouk, N.; Winter, K.R.; Riggle, P.J.; Rhodes, J.C.; Dodge, C.L.; Culp, D.W.; et al. The chsD and chsE genes of Aspergillus nidulans and their roles in chitin synthesis. Fungal Genet. Biol. 1996, 20, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2015, 44, D279–D285. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Siebert, S.; Farrell, J.A.; Cazet, J.F.; Abeykoon, Y.; Primack, A.S.; Schnitzler, C.E.; Juliano, C.E. Stem cell differentiation trajectories in Hydra resolved at single-cell resolution. Science 2019, 365, eaav9314. [Google Scholar] [CrossRef]
- Rathore, A.S.; Gupta, R.D. Chitinases from Bacteria to Human: Properties, Applications, and Future Perspectives. Enzym. Res. 2015, 2015, 791907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mali, B.; Möhrlen, F.; Frohme, M.; Frank, U. A putative double role of a chitinase in a cnidarian: Pattern formation and immunity. Dev. Comp. Immunol. 2004, 28, 973–981. [Google Scholar] [CrossRef]
- Darling, J.A.; Reitzel, A.R.; Burton, P.M.; Mazza, M.E.; Ryan, J.F.; Sullivan, J.C.; Finnerty, J.R. Rising starlet: The starlet sea anemone, Nematostella vectensis. BioEssays 2005, 27, 211–221. [Google Scholar] [CrossRef]
- Ormestad, M.; Martindale, M.Q.; Röttinger, E. A comparative gene expression database for invertebrates. EvoDevo 2011, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Layden, M.J.; Rentzsch, F.; Röttinger, E. The rise of the starlet sea anemone Nematostella vectensis as a model system to investigate development and regeneration. WIREs Dev. Biol. 2016, 5, 408–428. [Google Scholar] [CrossRef] [Green Version]
- Al-Shaer, L.; Havrilak, J.A.; Layden, M.J. Nematostella vectensis as a Model System. In Handbook of Marine Model Organisms in Experimental Biology; Boutet, A., Schierwater, B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 107–127. [Google Scholar]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Speir, M.L.; Bhaduri, A.; Markov, N.S.; Moreno, P.; Nowakowski, T.J.; Papatheodorou, I.; Pollen, A.A.; Raney, B.J.; Seninge, L.; Kent, W.J.; et al. UCSC Cell Browser: Visualize your single-cell data. Bioinformatics 2021, 37, 4578–4580. [Google Scholar] [CrossRef]
- Steger, J.; Cole, A.G.; Denner, A.; Lebedeva, T.; Genikhovich, G.; Ries, A.; Reischl, R.; Taudes, E.; Lassnig, M.; Technau, U. Single-cell transcriptomics identifies conserved regulators of neuroglandular lineages. Cell Rep. 2022, 40, 111370. [Google Scholar] [CrossRef] [PubMed]
- Dunn, D.F.; Liberman, M.H. Chitin in Sea Anemone Shells. Science 1983, 221, 157–159. [Google Scholar] [CrossRef]
- Wainwright, S.A. Skeletal Organization in the Coral, Pocillopora Damicornis. J. Cell Sci. 1963, 3, 169–183. [Google Scholar] [CrossRef]
- Chapman, D.M. Cnidarian Histology; Academic Press: New York, NY, USA, 1974; p. 92. [Google Scholar]
- Campbell, R.D. The Hydra of Madagascar (Cnidaria: Hydrozoa). Ann. De Limnol.-Int. J. Limnol. 1999, 35, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Arai, M.N. The potential importance of podocysts to the formation of scyphozoan blooms: A review. Hydrobiologia 2009, 616, 241–246. [Google Scholar] [CrossRef]
- Ikeda, H.; Ohtsu, K.; Uye, S.-I. Fine structure, Histochemistry, and Morphogenesis During Excystment of the Podocysts of the Giant Jellyfish Nemopilema nomurai (Scyphozoa, Rhizostomeae). Biol. Bull. 2011, 221, 248–260. [Google Scholar] [CrossRef]
- Babonis, L.S.; Martindale, M.Q. PaxA, but not PaxC, is required for cnidocyte development in the sea anemone Nematostella vectensis. EvoDevo 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of venom composition across a complex life cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef]
- Tournière, O.; Dolan, D.; Richards, G.S.; Sunagar, K.; Columbus-Shenkar, Y.Y.; Moran, Y.; Rentzsch, F. NvPOU4/Brain3 Functions as a Terminal Selector Gene in the Nervous System of the Cnidarian Nematostella vectensis. Cell Rep. 2020, 30, 4473–4489. [Google Scholar] [CrossRef]
- Tournière, O.; Gahan, J.M.; Busengdal, H.; Bartsch, N.; Rentzsch, F. Insm1-expressing neurons and secretory cells develop from a common pool of progenitors in the sea anemone Nematostella Vectensis. Sci. Adv. 2022, 8, eabi7109. [Google Scholar] [CrossRef]
- Shinzato, C.; Shoguchi, E.; Kawashima, T.; Hamada, M.; Hisata, K.; Tanaka, M.; Fujie, M.; Fujiwara, M.; Koyanagi, R.; Ikuta, T.; et al. Using the Acropora digitifera genome to understand coral responses to environmental change. Nature 2011, 476, 320–323. [Google Scholar] [CrossRef] [Green Version]
- Kenkel, C.D.; Meyer, E.; Matz, M.V. Gene expression under chronic heat stress in populations of the mustard hill coral (P orites astreoides) from different thermal environments. Mol. Ecol. 2013, 22, 4322–4334. [Google Scholar] [CrossRef] [PubMed]
- Brekhman, V.; Malik, A.; Haas, B.; Sher, N.; Lotan, T. Transcriptome profiling of the dynamic life cycle of the scypohozoan jellyfish Aurelia aurita. BMC Genom. 2015, 16, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simakov, O.; Marletaz, F.; Cho, S.J.; Edsinger-Gonzales, E.; Havlak, P.; Hellsten, U.; Kuo, D.H.; Larsson, T.; Lv, J.; Arendt, D.; et al. Insights into bilaterian evolution from three spiralian genomes. Nature 2013, 493, 526–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colbourne, J.K.; Pfrender, M.E.; Gilbert, D.; Thomas, W.K.; Tucker, A.; Oakley, T.H.; Tokishita, S.; Aerts, A.; Arnold, G.J.; Basu, M.K.; et al. The ecoresponsive genome of Daphnia pulex. Science 2011, 331, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millburn, G.H.; Crosby, M.A.; Gramates, L.S.; Tweedie, S.; FlyBase Consortium. FlyBase portals to human disease research using Drosophila models. Dis. Model. Mech. 2016, 9, 245–252. [Google Scholar] [PubMed] [Green Version]
- Zhu, Y.C.; Specht, C.A.; Dittmer, N.T.; Muthukrishnan, S.; Kanost, M.R.; Kramer, K.J. Sequence of a cDNA and expression of the gene encoding a putative epidermal chitin synthase of Manduca sexta. Insect Biochem. Mol. Biol. 2002, 32, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.J.; Takeuchi, T.; Koyanagi, R.; Yamada, L.; Kanda, M.; Khalturina, M.; Fujie, M.; Yamasaki, S.I.; Endo, K.; Satoh, N. The Lingula genome provides insights into brachiopod evolution and the origin of phosphate biomineralization. Nat. Commun. 2015, 6, 8301. [Google Scholar] [CrossRef] [Green Version]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Kimura, S.; Ogawa, Y.; Watanabe, S.; Soma, S.; Kaneko, T.; Yamada, L.; Sawada, H.; Tung, C.H.; Lu, T.M.; et al. Chitin-based barrier immunity and its loss predated mucus-colonization by indigenous gut microbiota. Nat. Commun. 2018, 9, 3402. [Google Scholar]
- Read, T.D.; Petit, R.A.; Joseph, S.J.; Alam, M.T.; Weil, M.R.; Ahmad, M.; Bhimani, R.; Vuong, J.S.; Haase, C.P.; Webb, D.H.; et al. Draft sequencing and assembly of the genome of the world’s largest fish, the whale shark: Rhincodon typus Smith 1828. BMC Genom. 2017, 18, 1–10. [Google Scholar]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.I.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Koutsovoulos, G.; Laetsch, D.R.; Stevens, L.; Kumar, S.; Horikawa, D.D.; Ishino, K.; Komine, S.; Kunieda, T.; Tomita, M.; et al. Comparative genomics of the tardigrades Hypsibius dujardini and Ramazzottius varieornatus. PLoS Biol. 2017, 15, e2002266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Horikawa, D.D.; Saito, Y.; Kuwahara, H.; Kozuka-Hata, H.; Shin-i, T.; Minakuchi, Y.; Ohishi, K.; Motoyama, A.; Aizu, T.; et al. Extremotolerant tardigrade genome and improved radiotolerance of human cultured cells by tardigrade-unique protein. Nat. Commun. 2016, 7, 12808. [Google Scholar] [CrossRef]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vandepas, L.E.; Tassia, M.G.; Halanych, K.M.; Amemiya, C.T. Unexpected Distribution of Chitin and Chitin Synthase across Soft-Bodied Cnidarians. Biomolecules 2023, 13, 777. https://doi.org/10.3390/biom13050777
Vandepas LE, Tassia MG, Halanych KM, Amemiya CT. Unexpected Distribution of Chitin and Chitin Synthase across Soft-Bodied Cnidarians. Biomolecules. 2023; 13(5):777. https://doi.org/10.3390/biom13050777
Chicago/Turabian StyleVandepas, Lauren E., Michael G. Tassia, Kenneth M. Halanych, and Chris T. Amemiya. 2023. "Unexpected Distribution of Chitin and Chitin Synthase across Soft-Bodied Cnidarians" Biomolecules 13, no. 5: 777. https://doi.org/10.3390/biom13050777