
Physico-Chemical Changes Induced by Gamma Irradiation on Some Structural Protein Extracts

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction Procedures
2.3. Instrumentation and Sample Preparation
2.3.1. Infrared Spectroscopy
2.3.2. Circular Dichroism (CD) Spectroscopy
2.3.3. Differential Scanning Microcalorimetry (μDSC)
2.3.4. Electron Paramagnetic Resonance (EPR) Spectroscopy
2.3.5. UVA and Gamma Irradiation
3. Results and Discussions
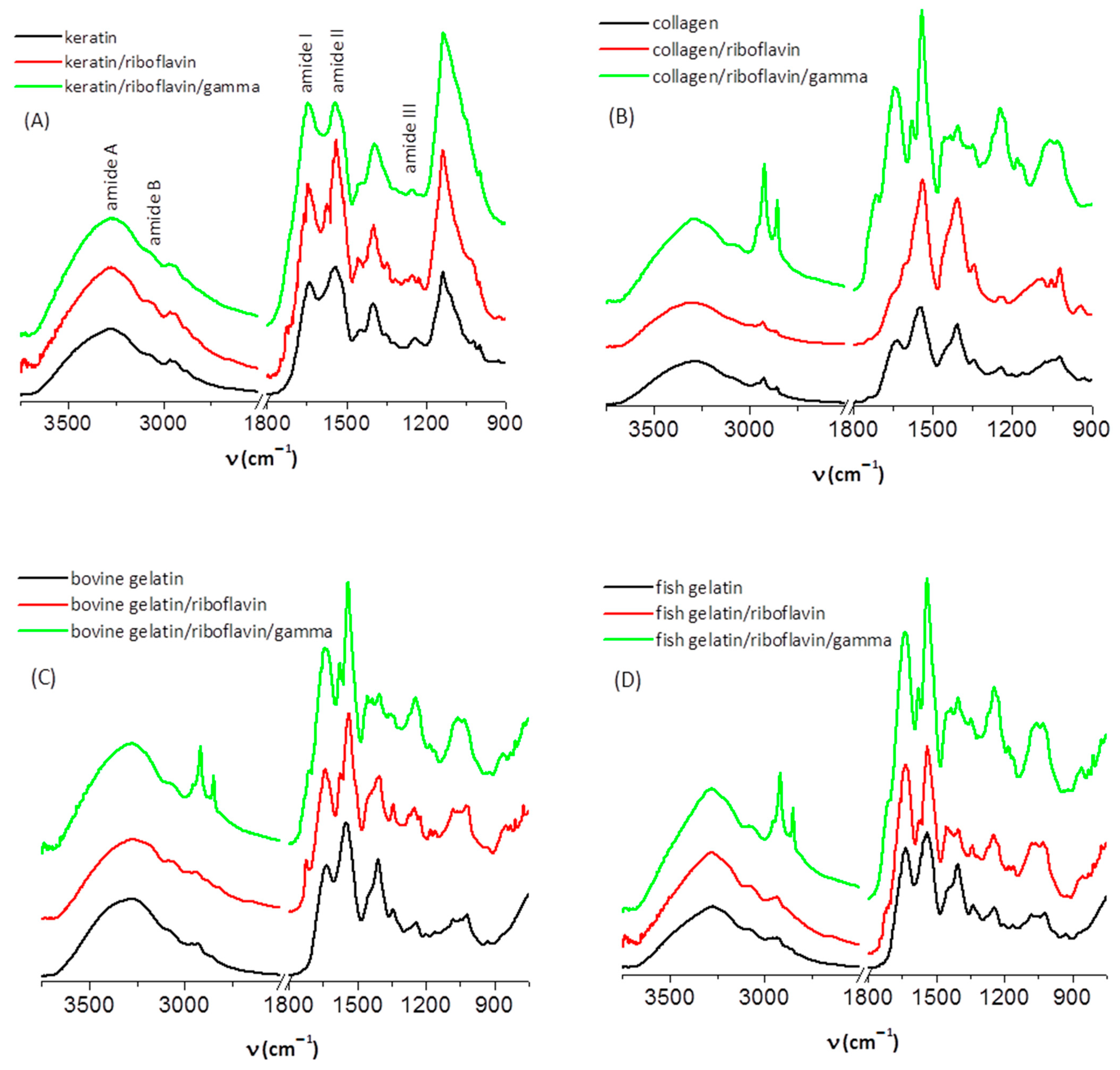
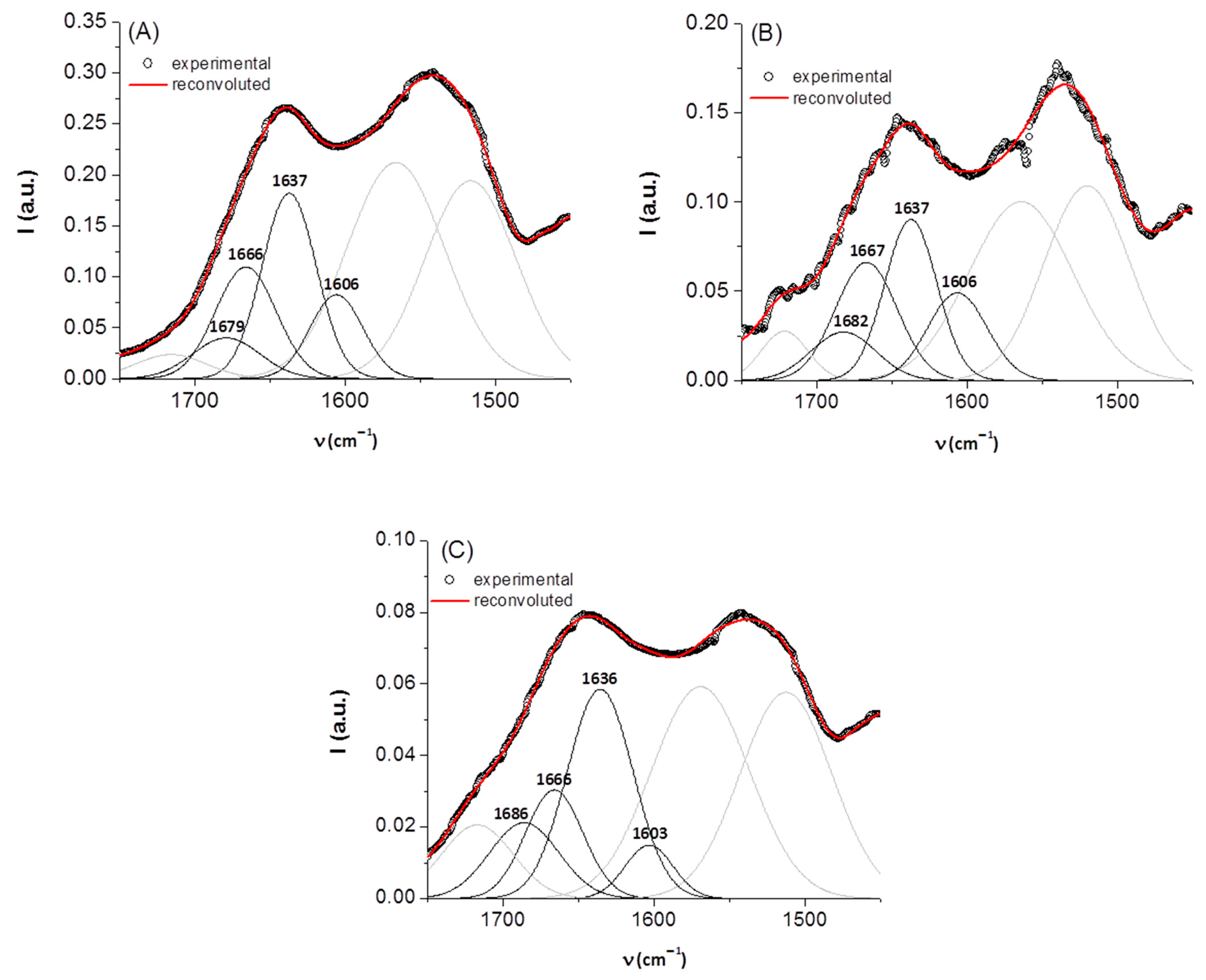
3.1. IR Spectroscopy
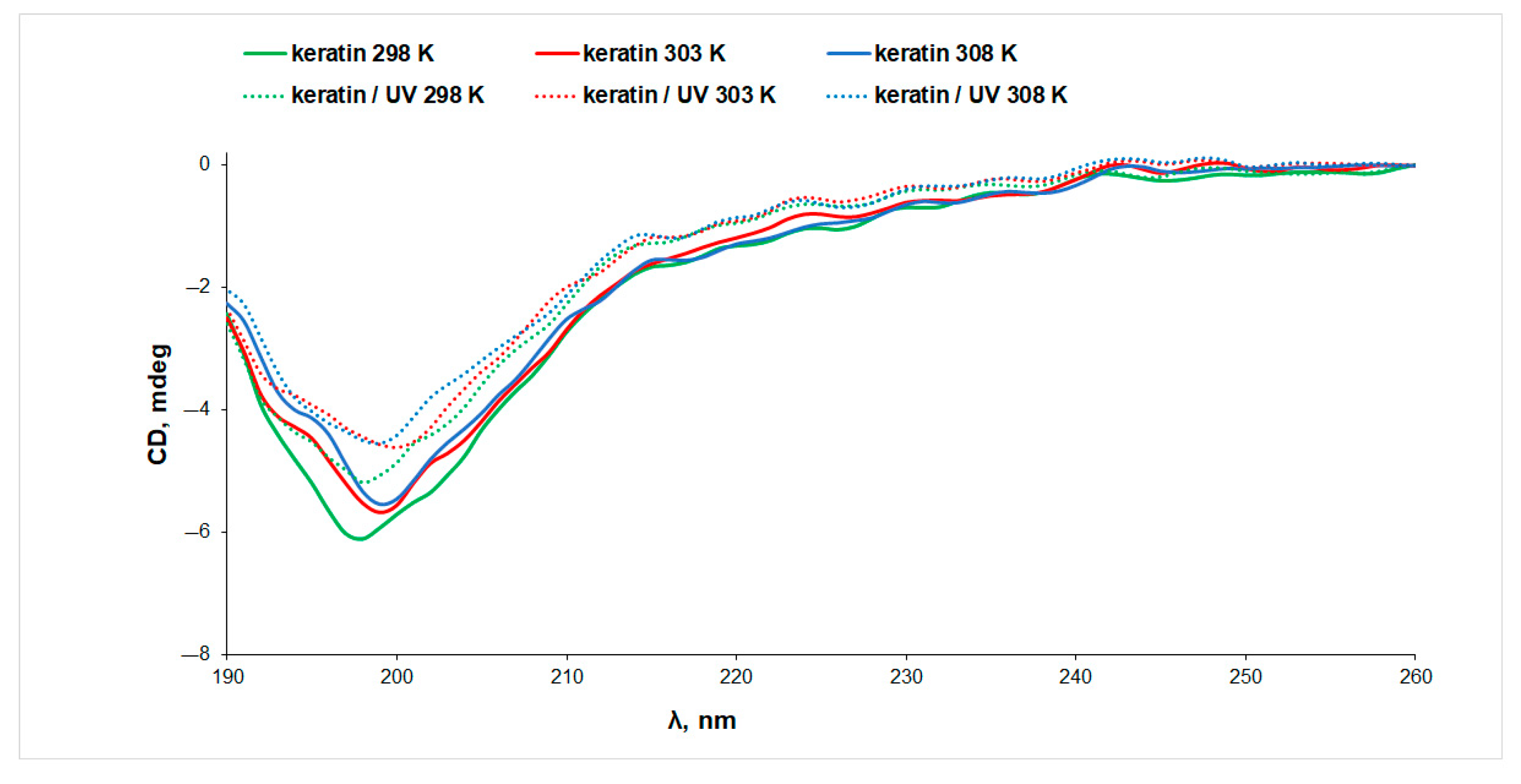
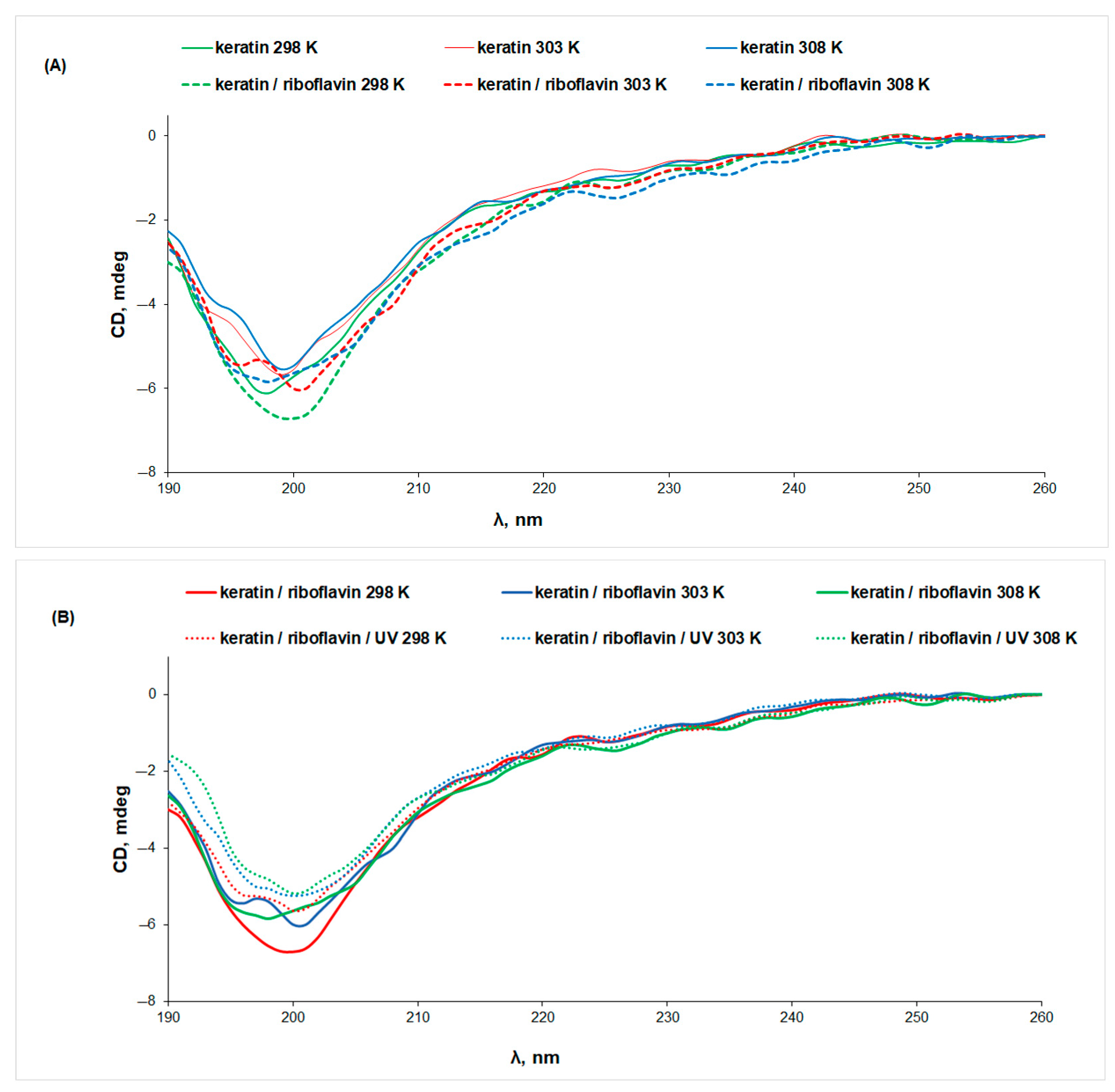
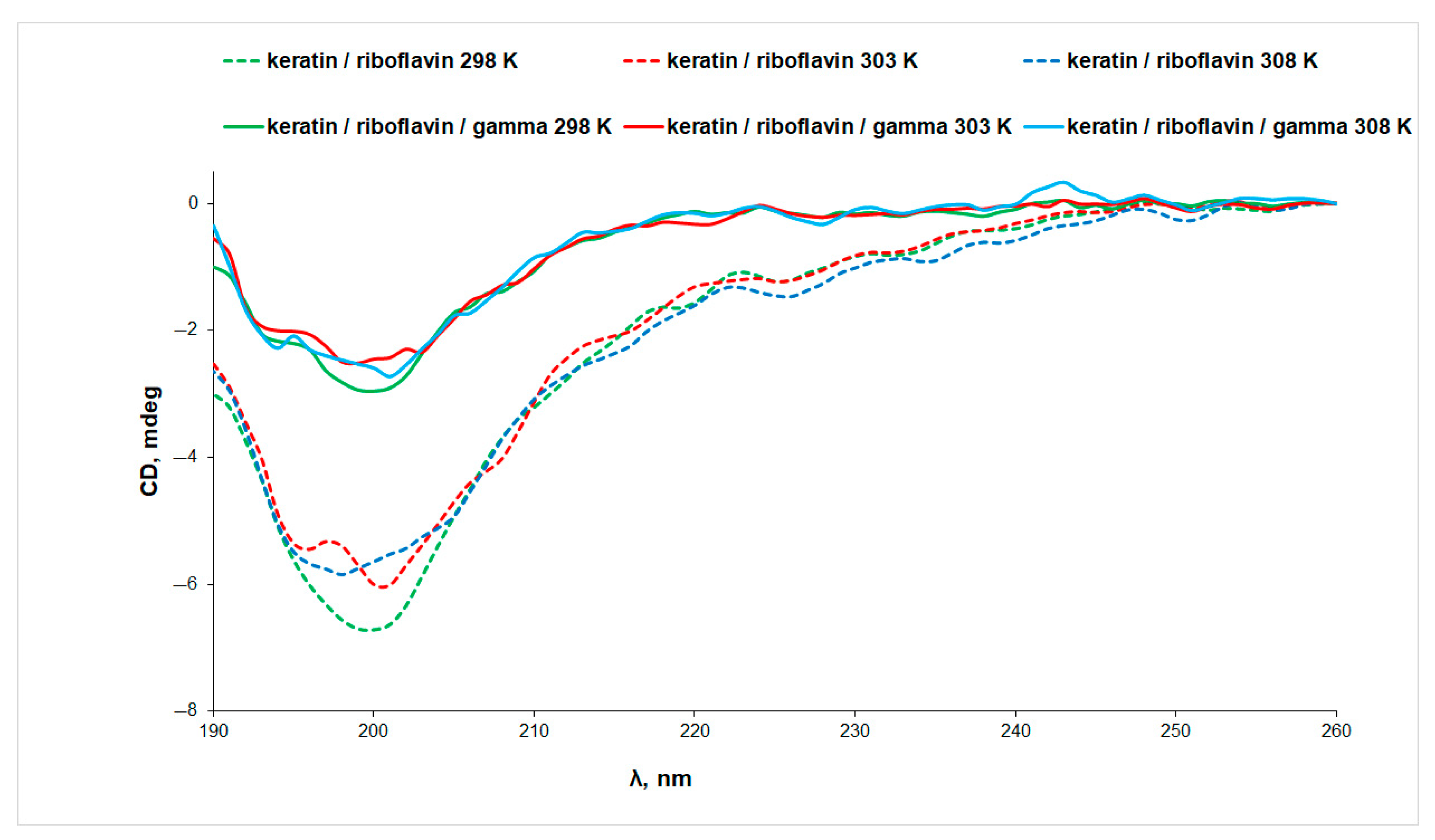
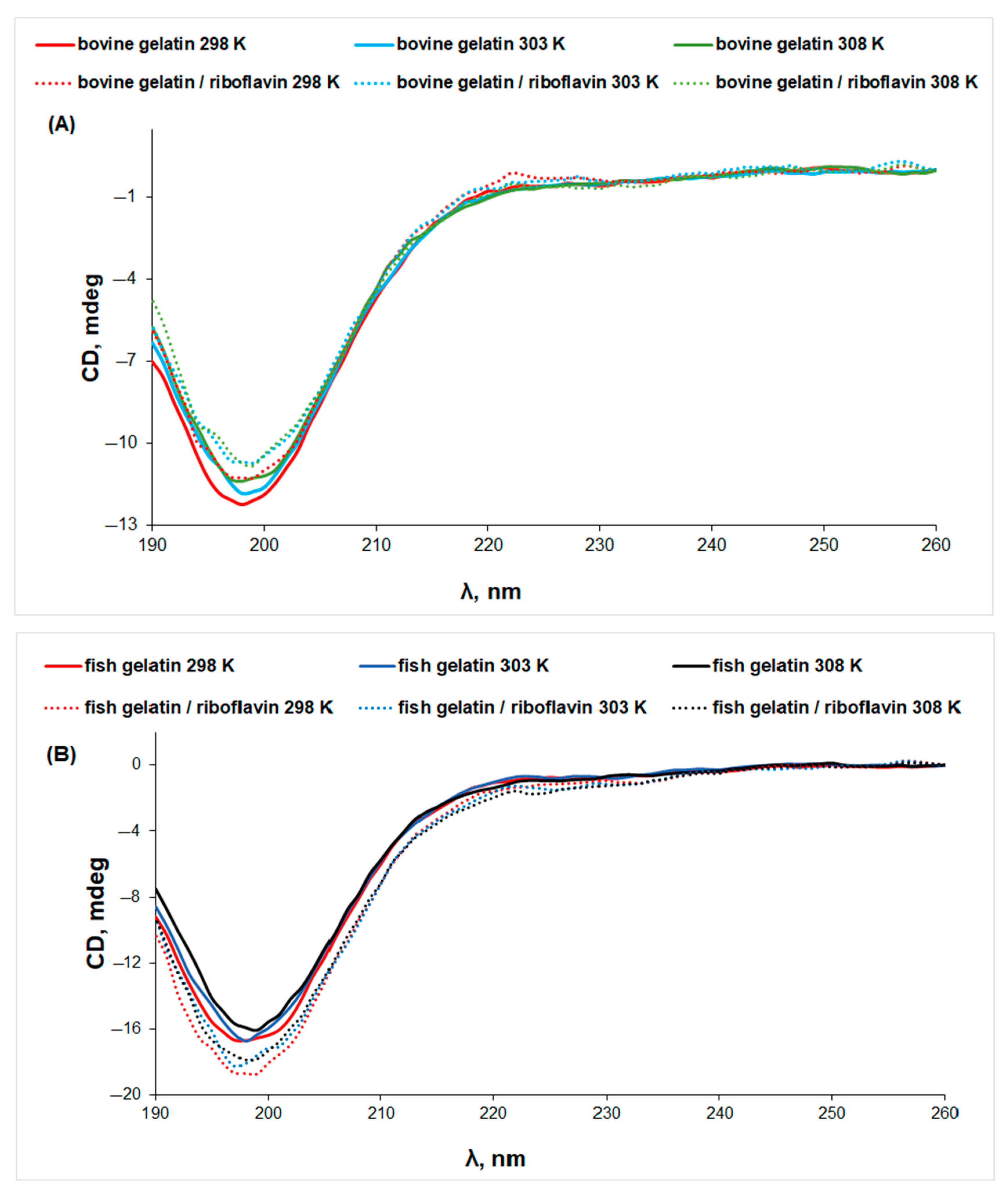
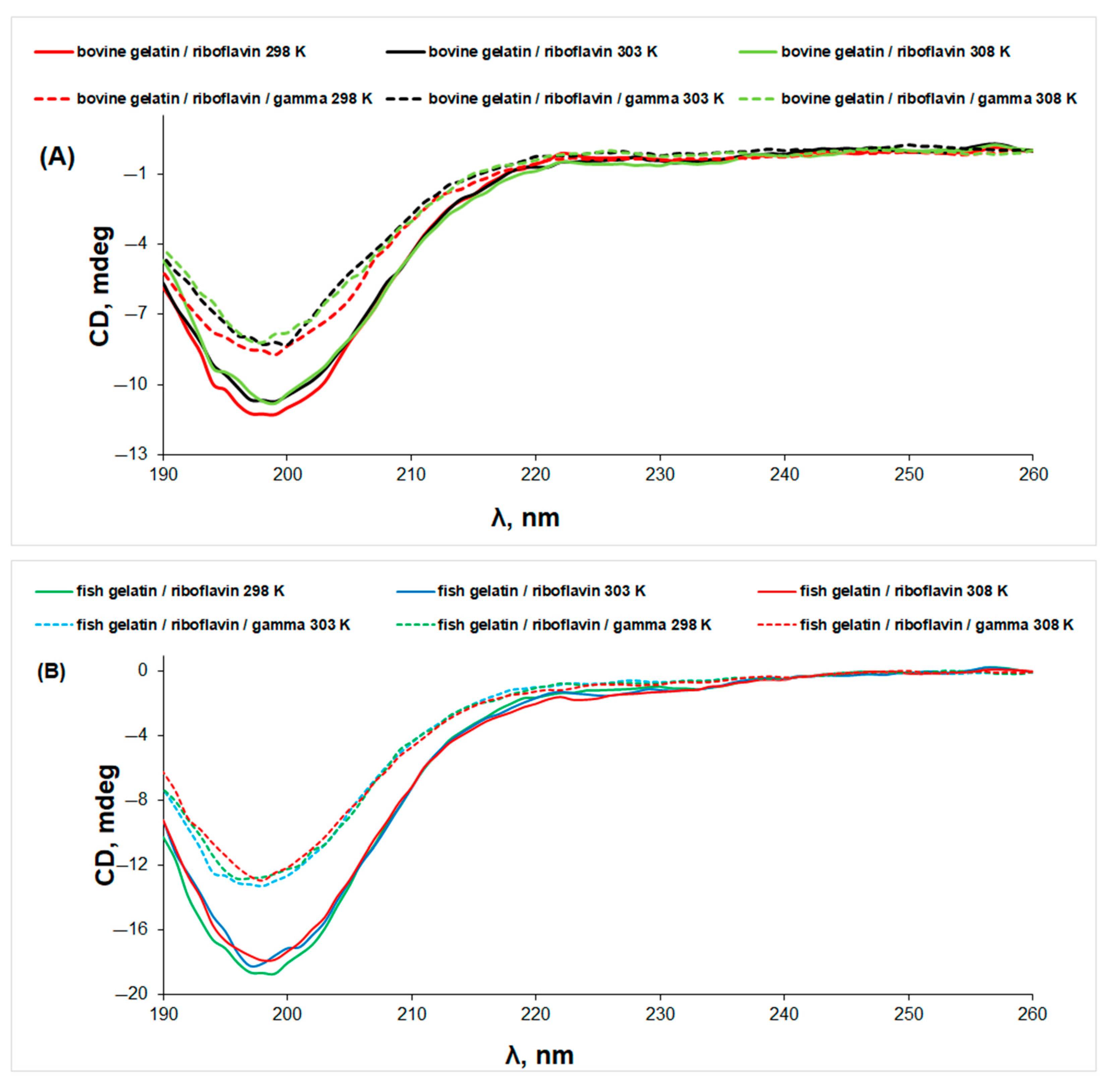
3.2. Circular Dichroism (CD) Spectroscopy
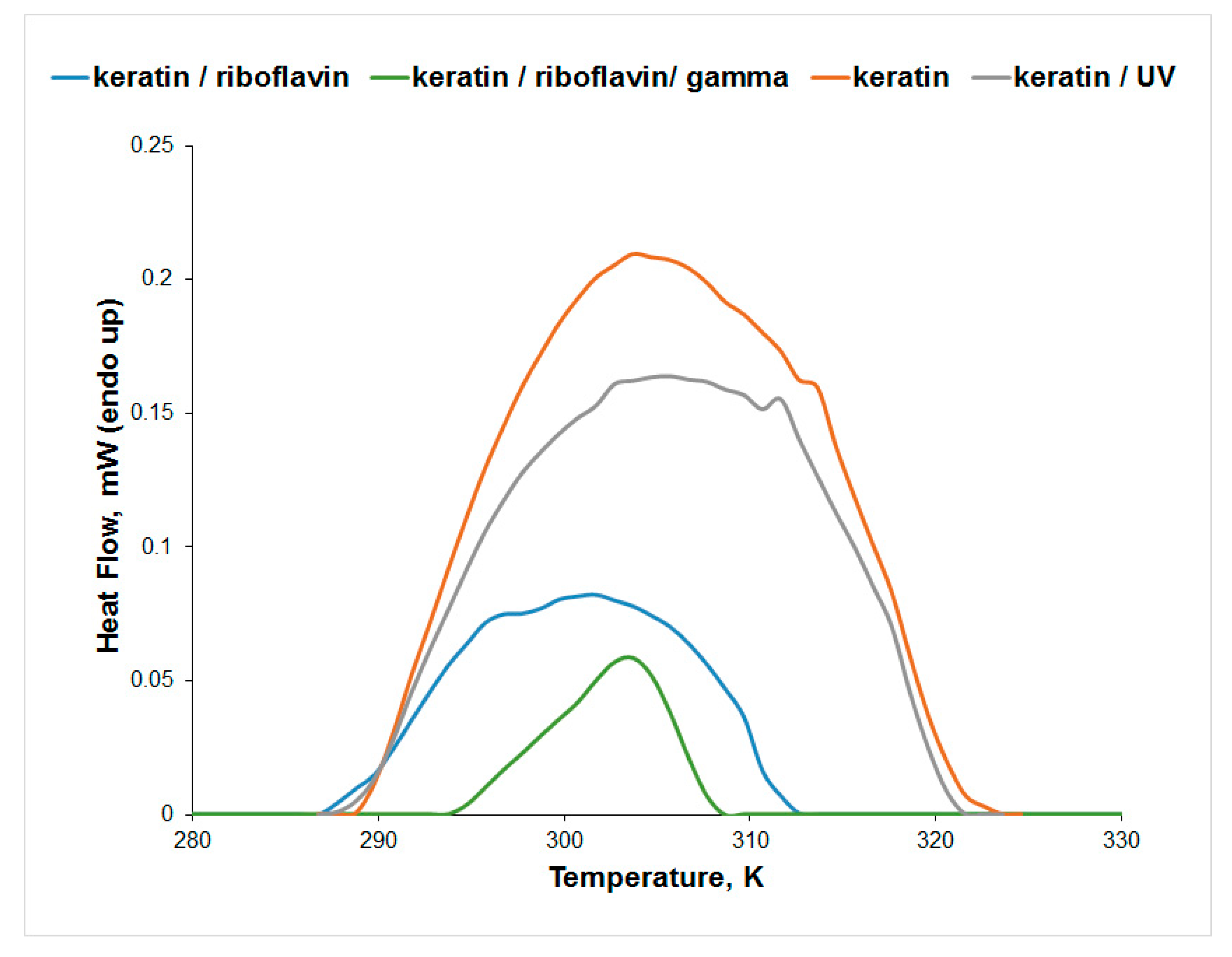
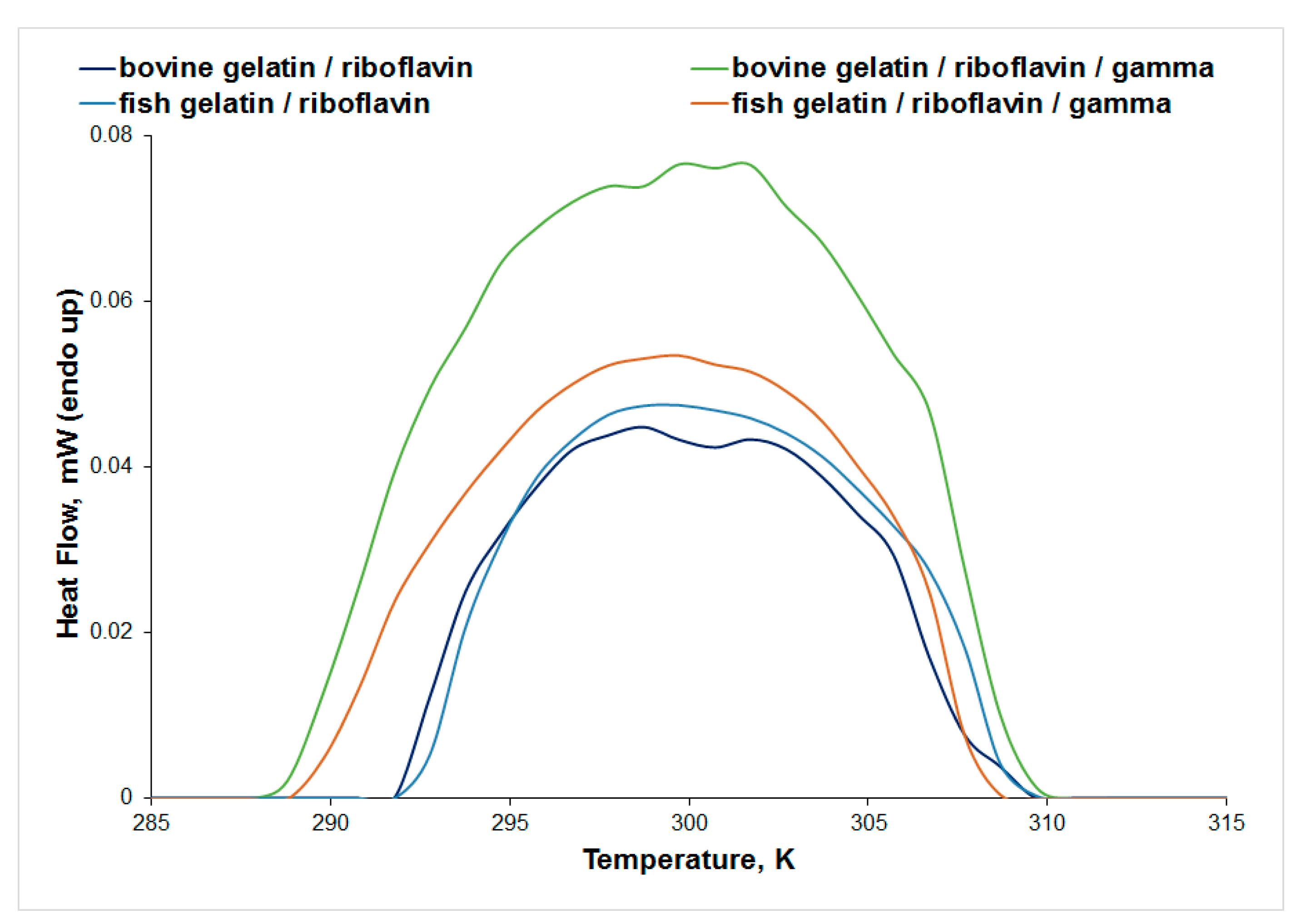
3.3. Differential Scanning Microcalorimetry (μDSC)
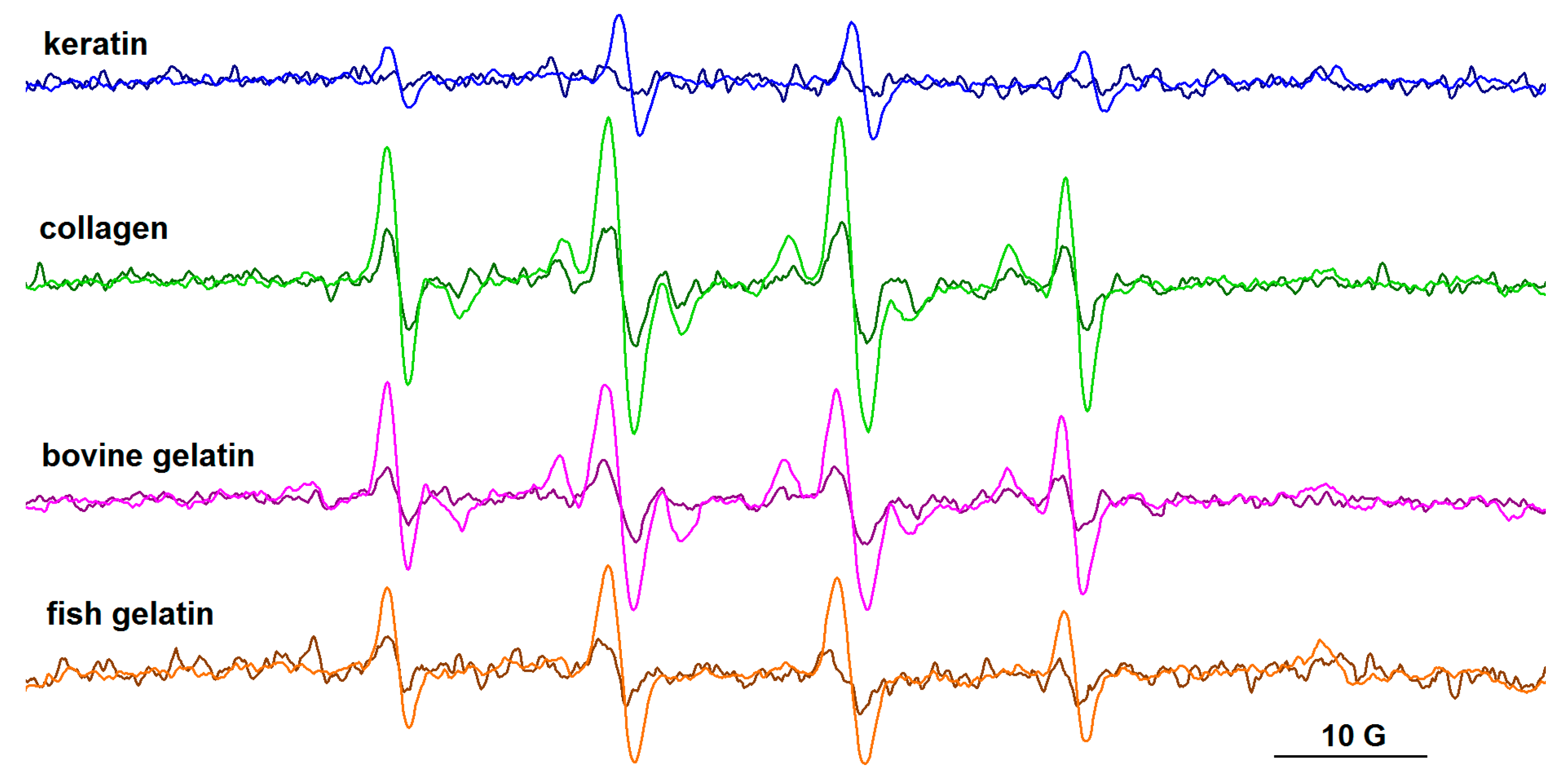
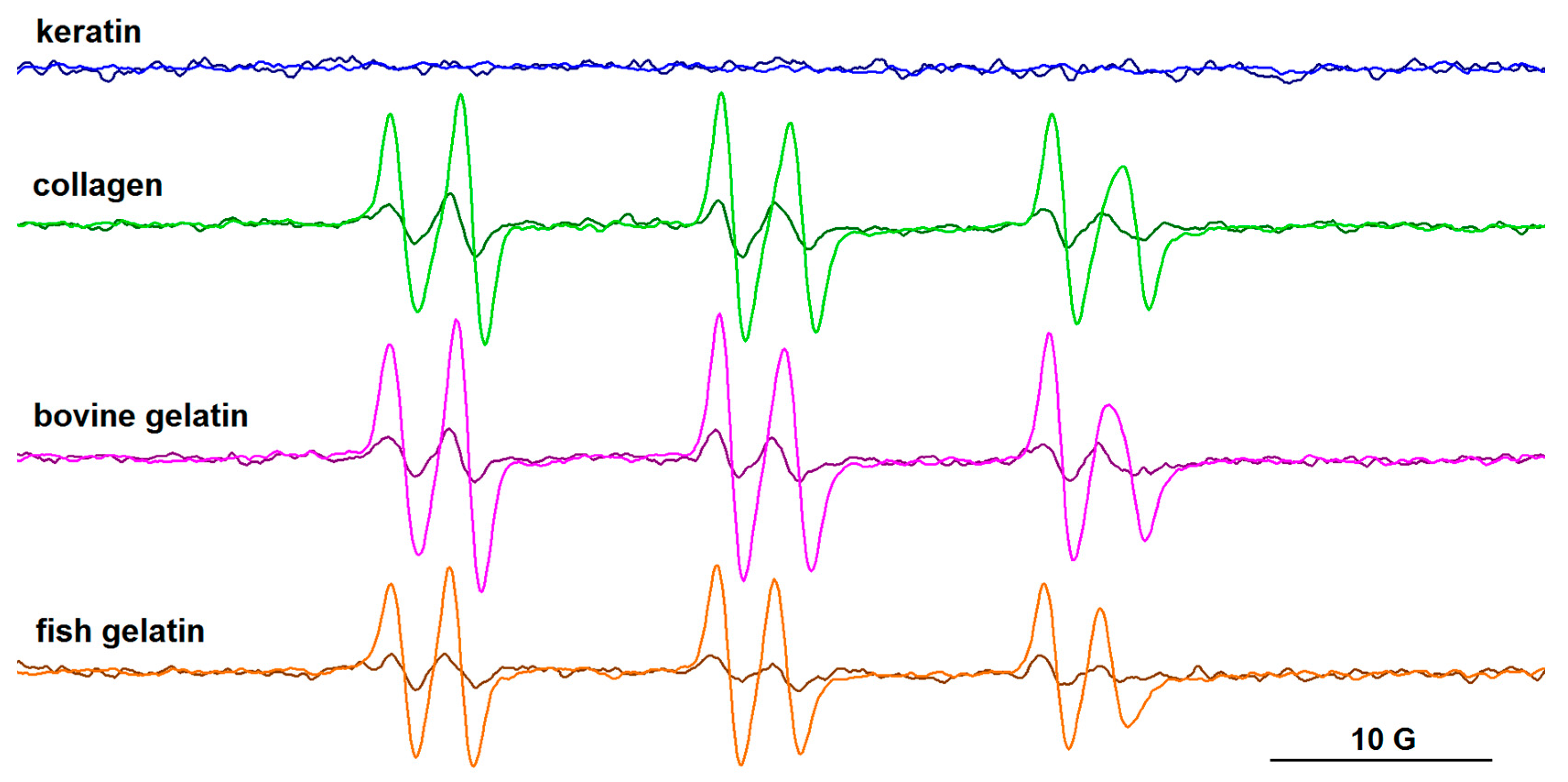
3.4. EPR Spectroscopy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodrigues, L.M.; Sales, L.A.; Fontes, P.R.; de Almeida Torres Filho, R.; Pereira Dutra Andrade, M.; de Lemos Souza Ramos, A.; Mendes Ramos, E. Combined effects of gamma irradiation and aging on tenderness and quality of beef from Nellore cattle. Food Chem. 2020, 313, 126137. [Google Scholar] [CrossRef] [PubMed]
- Burton, B.; Gaspar, A.; Josey, D.; Tupy, J.; Grynpas, M.D.; Willett, T.L. Bone embrittlement and collagen modifications due to high-dose gamma-irradiation sterilization. Bone 2014, 61, 71–81. [Google Scholar] [CrossRef]
- Azorin, E.; González-Martínez, P.R.; Azorin, J. Collagen I confers gamma radiation resistance. Appl. Radiat. Isot. 2012, 71, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Morgan, D.A.; Forwood, M.R. Sterilization of allograft bone: Effects of gamma irradiation on allograft biology and biomechanics. Cell Tissue Bank. 2007, 8, 93–105. [Google Scholar] [CrossRef]
- Gomes, A.D.; de Oliveira, A.A.R.; Houmard, M.; Nunes, E.H.M. Gamma sterilization of collagen/hydroxyapatite composites: Validation and radiation effects. Appl. Radiat. Isot. 2021, 174, 109758. [Google Scholar] [CrossRef] [PubMed]
- Takitoh, T.; Bessho, M.; Hirose, M.; Ohgushi, H.; Mori, H.; Hara, M. Gamma-cross-linked nonfibrillar collagen gel as a scaffold for osteogenic differentiation of mesenchymal stem cells. J. Biosci. Bioeng. 2015, 119, 217–225. [Google Scholar] [CrossRef]
- Liu, B.; Huang, W.; Yang, G.; An, Y.; Yin, Y.; Wang, N.; Jiang, B. Preparation of gelatin/poly (γ-glutamic acid) hydrogels with stimulated response by hot-pressing preassembly and radiation crosslinking. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 116, 111259. [Google Scholar] [CrossRef]
- Sun, L.L.; Li, L.H.; Wang, Y.Q.; Li, M.B.; Xu, S.M.; Zhang, C.P. A collagen-based bi-layered composite dressing for accelerated wound healing. J. Tissue Viability 2022, 31, 180–189. [Google Scholar] [CrossRef]
- Rapa, M.; Zaharescu, T.; Stefan, L.M.; Gaidau, C.; Stanculescu, I.; Constantinescu, R.R.; Stanca, M. Bioactivity and thermal stability of collagen–chitosan containing lemongrass essential oil for potential medical applications. Polymers 2022, 14, 3884. [Google Scholar] [CrossRef]
- Olariu, L.; Brindusa, G.D.; Gaidau, C.; Stanca, M.; Tanase, L.M.; Ene, M.D.; Stanculescu, I.R.; Tablet, C. Bioactive low molecular weight keratin hydrolysates for improving skin wound healing. Polymers 2022, 14, 1125. [Google Scholar] [CrossRef]
- Constantin, M.M.; Corbu, C.G.; Mocanu, S.; Popescu, E.I.; Micutz, M.; Staicu, T.; Şomoghi, R.; Trică, B.; Popa, V.T.; Precupas, A.; et al. Model systems for evidencing the mediator role of riboflavin in the UVA cross-linking treatment of keratoconus. Molecules 2022, 27, 190. [Google Scholar] [CrossRef] [PubMed]
- Wollensak, G.; Spoerl, E.; Seiler, T. Riboflavin/ultraviolet-a-induced collagen crosslinking for the treatment of keratoconus. Am. J. Ophthalmol. 2003, 135, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Frullini, A.; Manetti, L.; Di Cicco, E.; Fortuna, D. Photoinduced collagen cross-linking: A new approach to venous insufficiency. Dermatol. Surg. 2011, 37, 1113–1118. [Google Scholar] [CrossRef]
- Hardan, L.; Daood, U.; Bourgi, R.; Cuevas-Suárez, C.E.; Devoto, W.; Zarow, M.; Jakubowicz, N.; Zamarripa-Calderón, J.E.; Radwanski, M.; Orsini, G.; et al. Effect of collagen crosslinkers on dentin bond strength of adhesive systems: A systematic review and meta-analysis. Cells 2022, 11, 2417. [Google Scholar] [CrossRef] [PubMed]
- Akagündüz, Y.; Mosquera, M.; Giménez, B.; Alemán, A.; Montero, P.; Gómez-Guillén, M.C. Sea bream bones and scales as a source of gelatin and ACE inhibitory peptides. LWT-Food Sci. Technol. 2014, 55, 579–585. [Google Scholar] [CrossRef]
- Voicescu, M.; Ionescu, S.; Nistor, C.L. Spectroscopic study of 3-Hydroxyflavone–protein interaction in lipidic bi-layers immobilized on silver nanoparticles. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2017, 170, 1–8. [Google Scholar] [CrossRef]
- Stoian, A.-M.; Matei, I.; Ionescu, S. Global and local conformational changes in albumin–ligand interactions through spectroscopic methods. Rev. Roum. Chim. 2017, 62, 623–628. Available online: https://revroum.lew.ro/wp-content/uploads/2017/8/Art%2006.pdf (accessed on 27 April 2023).
- Duling, D.R. PEST Winsim; version 0.96; National Institute of Environmental Health Sciences: Triangle Park, NC, USA, 1996.
- Duling, D.R. Simulation of multiple isotropic spin-trap EPR spectra. J. Magn. Reson. B 1994, 104, 105–110. [Google Scholar] [CrossRef]
- Bart, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta 2007, 1767, 1073–1101. [Google Scholar] [CrossRef]
- Stani, C.; Vaccari, L.; Mitri, E.; Birarda, G. FTIR investigation of the secondary structure of type I collagen: New insight into the amide III band. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 229, 118006. [Google Scholar] [CrossRef]
- de Campos Vidal, B.; Mello, M.L.S. Collagen type I amide I band infrared spectroscopy. Micron 2011, 42, 283–289. [Google Scholar] [CrossRef]
- Wojciechowska, E.; Włochowicz, A.; Wesełucha-Birczyńska, A. Application of Fourier-transform infrared and Raman spectroscopy to study degradation of the wool fiber keratin. J. Mol. Struct. 1999, 511–512, 307–318. [Google Scholar] [CrossRef]
- Gaidau, C.; Stanca, M.; Niculescu, M.-D.; Alexe, C.-A.; Becheritu, M.; Horoias, R.; Cioineag, C.; Rapa, M.; Stanculescu, I.R. Wool keratin hydrolysates for bioactive additives preparation. Materials 2021, 14, 4696. [Google Scholar] [CrossRef] [PubMed]
- Gaidau, C.; Epure, D.-G.; Enascuta, C.E.; Carsote, C.; Sendrea, C.; Proietti, N.; Chen, W.; Gu, H. Wool keratin total solubilisation for recovery and reintegration—An ecological approach. J. Clean. Prod. 2019, 236, 117586. [Google Scholar] [CrossRef]
- Odlyha, M.; Theodorakopoulos, C.; Campana, R. Studies on woollen threads from historical tapestries. AUTEX Res. J. 2007, 7, 1. [Google Scholar]
- Sionkowska, A.; Skopinska-Wisniewska, J.; Kozlowska, J.; Planecka, A.; Kurzawa, M. Photochemical behaviour of hydrolysed keratin. Int. J. Cosmet. Sci. 2011, 33, 503–508. [Google Scholar] [CrossRef]
- Pelton, J.T.; McLean, L.R. Spectroscopic methods for analysis of protein secondary structure. Anal. Biochem. 2000, 277, 167–176. [Google Scholar] [CrossRef]
- Drobota, M.; Gradinaru, L.M.; Vlad, S.; Bargan, A.; Butnaru, M.; Angheloiu, M.; Aflori, M. Preparation and characterization of electrospun collagen based composites for biomedical applications. Materials 2020, 13, 3961. [Google Scholar] [CrossRef]
- Pielesz, A.; Wlochowicz, A.; Binias, W. The evaluation of structural changes in wool fibre keratin treated with azo dyes by Fourier Transform Infrared Spectroscopy. Spectrochim. Acta A 2000, 56, 1409–1420. [Google Scholar] [CrossRef]
- Muyonga, J.H.; Cole, C.G.B.; Duodu, K.G. Fourier transform infrared (FTIR) spectroscopic study of acid soluble collagen and gelatin from skins and bones of young and adult Nile perch (Lates niloticus). Food Chem. 2004, 86, 325–332. [Google Scholar] [CrossRef]
- Kakkar, P.; Madhan, B.; Shanmugam, G. Extraction and characterization of keratin from bovine hoof: A potential material for biomedical applications. SpringerPlus 2014, 3, 596. [Google Scholar] [CrossRef] [PubMed]
- Gopal, R.; Park, J.S.; Seo, C.H.; Park, Y. Applications of circular dichroism for structural analysis of gelatin and antimicrobial peptides. Int. J. Mol. Sci. 2012, 13, 3229–3244. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Xu, G.; Sun, Y.; Zhang, H.; Mao, H.; Feng, Y. Interaction between proteins and cationic gemini surfactant. Biomacromolecules 2007, 8, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Tiffany, M.L.; Krimm, S. Circular dichroism of the “random” polypeptide chain. Biopolymers 1969, 8, 347–359. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S.; Ocen, D.; Yang, N.; Xu, B.; Zhang, L.; Xu, X. Physical and chemical properties of gelatin from the skin of cultured Amur sturgeon (Acipenser schrenckii). J. Appl. Ichtyol. 2013, 29, 943–950. [Google Scholar] [CrossRef]
- Lois Tifany, M.; Krimm, S. Effect of temperature on the circular dichroism spectra of polypeptides in the extended state. Biopolymers 1972, 11, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Komsa-Penkova, R.; Koynova, R.; Kostov, G.; Tenchov, B.G. Thermal stability of calf skin collagen type I in salt solutions. BBA Protein Struct. Mol. Enzymol. 1996, 1297, 171–181. [Google Scholar] [CrossRef]
- Wright, N.T.; Humphrey, J.D. Denaturation of collagen via heating: An irreversible rate process. Annu. Rev. Biomed. Eng. 2002, 4, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Sarbon, N.M.; Badii, F.; Howell, N.K. The effect of chicken skin gelatin and whey protein interactions on rheological and thermal properties. Food Hydrocoll. 2015, 45, 83–92. [Google Scholar] [CrossRef]
- Buettner, G.R. Spin trapping: ESR parameters of spin adducts. Free Radic. Biol. Med. 1987, 3, 259–303. [Google Scholar] [CrossRef]
- Davies, M.J.; Fu, S.; Dean, R.T. Protein hydroperoxides can give rise to reactive free radicals. Biochem. J. 1995, 305, 643–649. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Wavenumber (cm−1) | Assignment |
|---|---|---|
| keratin | 3275, 3060 2963, 2935 1639 (s) 1544 (s) 1448 (w), 1399 1350 (w), 1242 (w) 1136 (s) | O-H, N-H stretching (amide A and B) C-H stretching (asymmetric, symmetric) C=O stretching/N-H bending (amide I) C-N stretching/N-H bending (amide II) C-H bending C-N stretching/N-H bending (amide III) S-O stretching, symmetric (-SO2-S-, cystine dioxide) |
| keratin/riboflavin | 3275, 3072 2963, 2935 1642 (s) 1540 (s) 1455 (w), 1398 1347 (w), 1250 1135 (s) | N-H stretching (amide A and B) C-H stretching (asymmetric, symmetric) C=O stretching/N-H bending (amide I) C-N stretching/N-H bending (amide II) C-H bending C-N stretching/N-H bending (amide III) S-O stretching, symmetric (-SO2-S-, cystine dioxide) |
| keratin/riboflavin/gamma | 3275, 3068 2971, 2934 1644 (s) 1542 (s) 1394 1251 1133 (s) | N-H stretching (amide A and B) C-H stretching (asymmetric, symmetric) C=O stretching/N-H bending (amide I) C-N stretching/N-H bending (amide II) C-H bending C-N stretching/N-H bending (amide III) S-O stretching, symmetric (-SO2-S-, cystine dioxide) |
| Sample | Band Assignment | ν (cm−1) | % Area |
|---|---|---|---|
| keratin | β-sheet random coil α-helix β-turn | 1606 1637 1666 1679 | 19 42 28 11 |
| keratin/riboflavin | β-sheet random coil α-helix β-turn | 1606 1637 1667 1682 | 22 35 30 13 |
| keratin/riboflavin/gamma | β-sheet random coil α-helix β-turn | 1603 1636 1666 1686 | 9 49 23 19 |
| Sample | Tpeak (K) | ΔH (J/g) |
|---|---|---|
| keratin | 303.87 | 2.69 |
| keratin/UV-irradiated | 305.50 | 2.15 |
| keratin/riboflavin | 301.68 | 0.76 |
| keratin/riboflavin/gamma-irradiated | 303.44 | 0.26 |
| collagen/riboflavin | 299.09 | 0.55 |
| collagen/riboflavin/gamma-irradiated | 299.59 | 0.79 |
| bovine gelatin/riboflavin | 298.96 | 0.32 |
| bovine gelatin/riboflavin/gamma-irradiated | 301.32 | 0.66 |
| fish gelatin/riboflavin | 299.56 | 0.35 |
| fish gelatin/riboflavin/gamma-irradiated | 299.49 | 0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanca, M.; Gaidau, C.; Zaharescu, T.; Balan, G.-A.; Matei, I.; Precupas, A.; Leonties, A.R.; Ionita, G. Physico-Chemical Changes Induced by Gamma Irradiation on Some Structural Protein Extracts. Biomolecules 2023, 13, 774. https://doi.org/10.3390/biom13050774
Stanca M, Gaidau C, Zaharescu T, Balan G-A, Matei I, Precupas A, Leonties AR, Ionita G. Physico-Chemical Changes Induced by Gamma Irradiation on Some Structural Protein Extracts. Biomolecules. 2023; 13(5):774. https://doi.org/10.3390/biom13050774
Chicago/Turabian StyleStanca, Maria, Carmen Gaidau, Traian Zaharescu, George-Alin Balan, Iulia Matei, Aurica Precupas, Anca Ruxandra Leonties, and Gabriela Ionita. 2023. "Physico-Chemical Changes Induced by Gamma Irradiation on Some Structural Protein Extracts" Biomolecules 13, no. 5: 774. https://doi.org/10.3390/biom13050774