
Plasmalogens: Free Radical Reactivity and Identification of Trans Isomers Relevant to Biological Membranes

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Equipment
3. Methods
3.1. Transesterification Procedures of C18 Plasm-20:4 PC, FAME, Soybean Lecithin and RBC
3.2. Plasmalogen Cis-Trans Isomerization
3.3. Preparation of Liposomes
3.4. Preparation of RBC Ghosts
3.5. Isolation of Plasmalogens from RBC Membranes
3.6. Incubation Experiments under Oxidative Conditions
3.7. Statistical Analysis
4. Results and Discussion
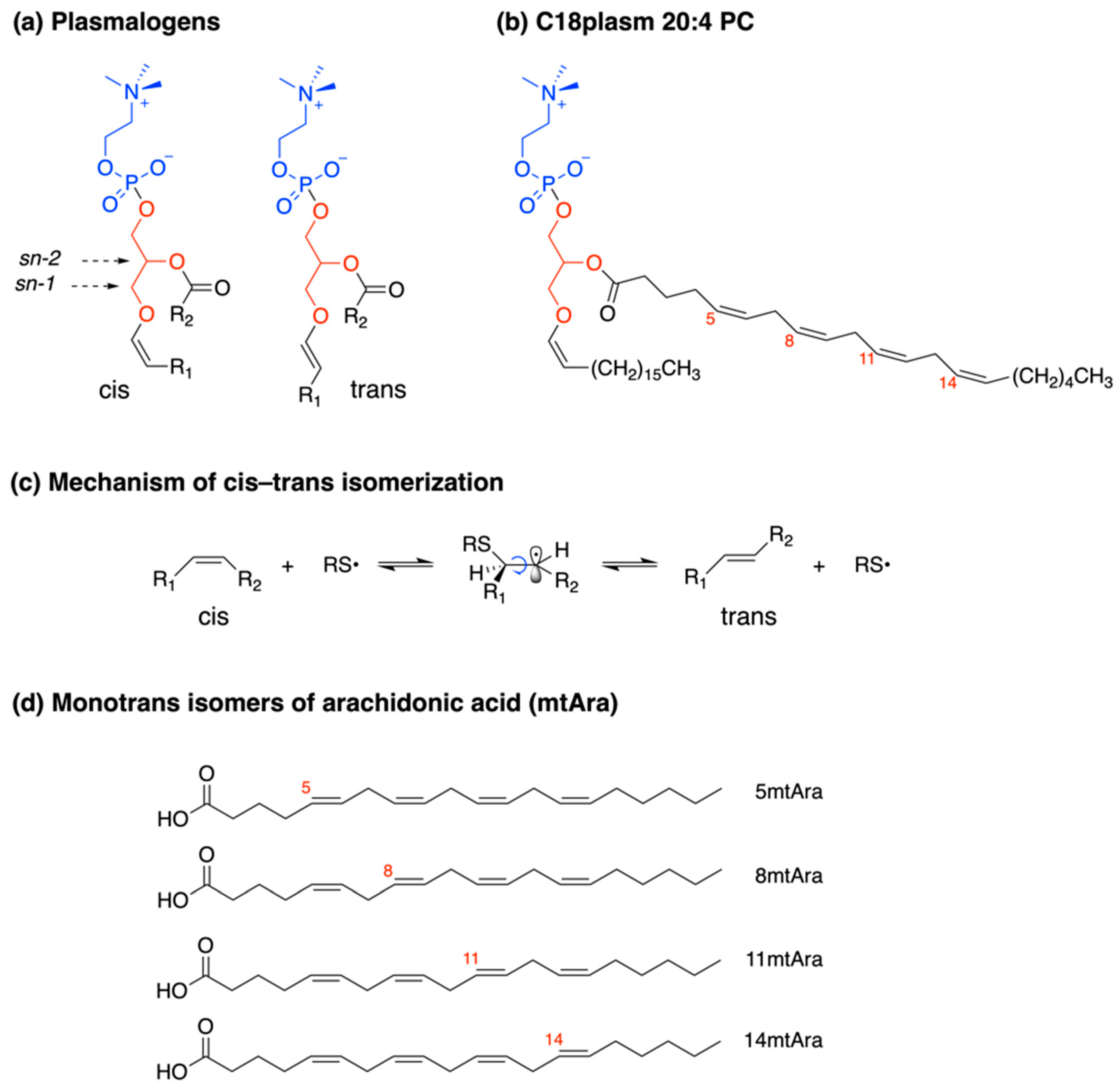
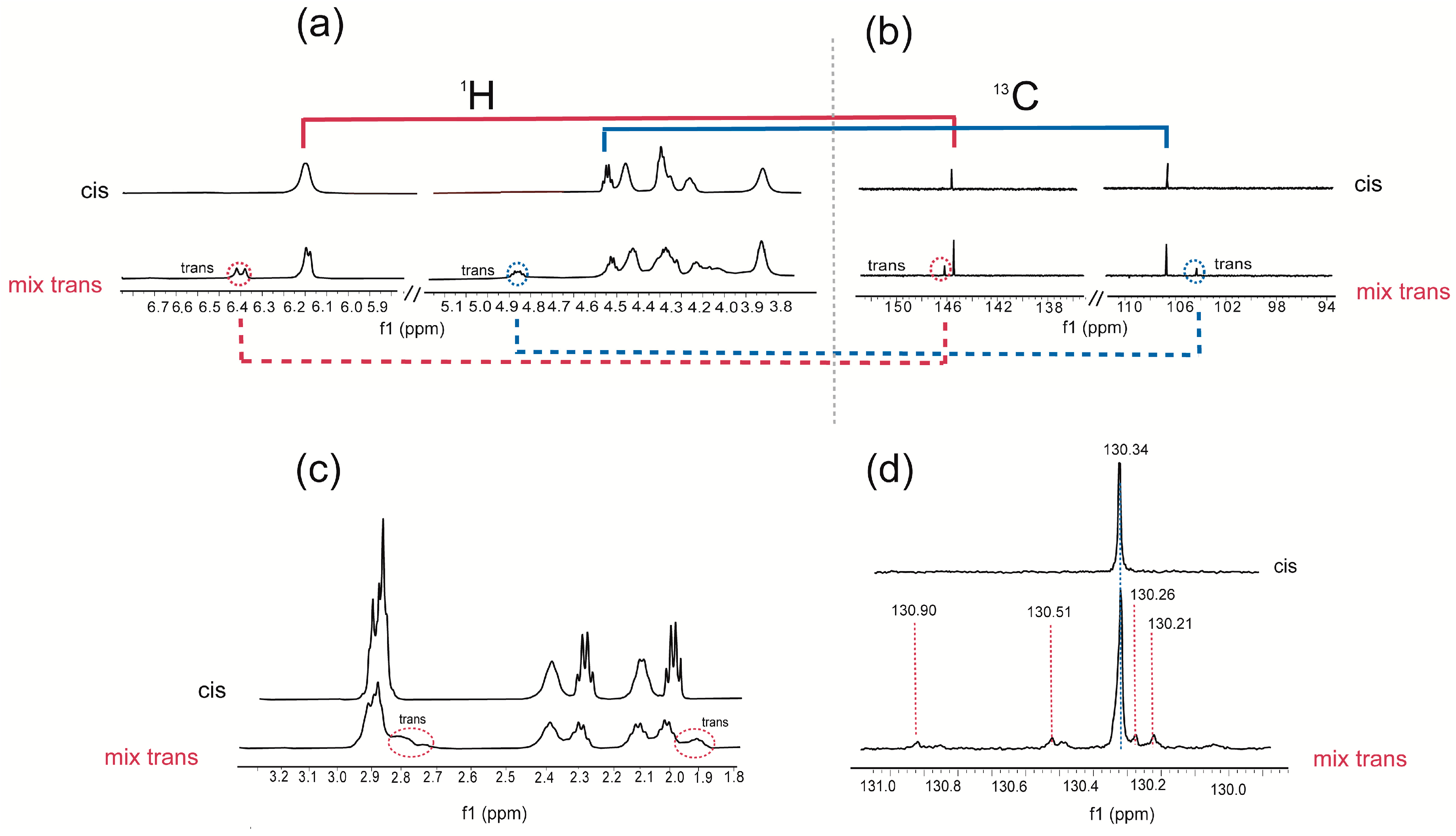
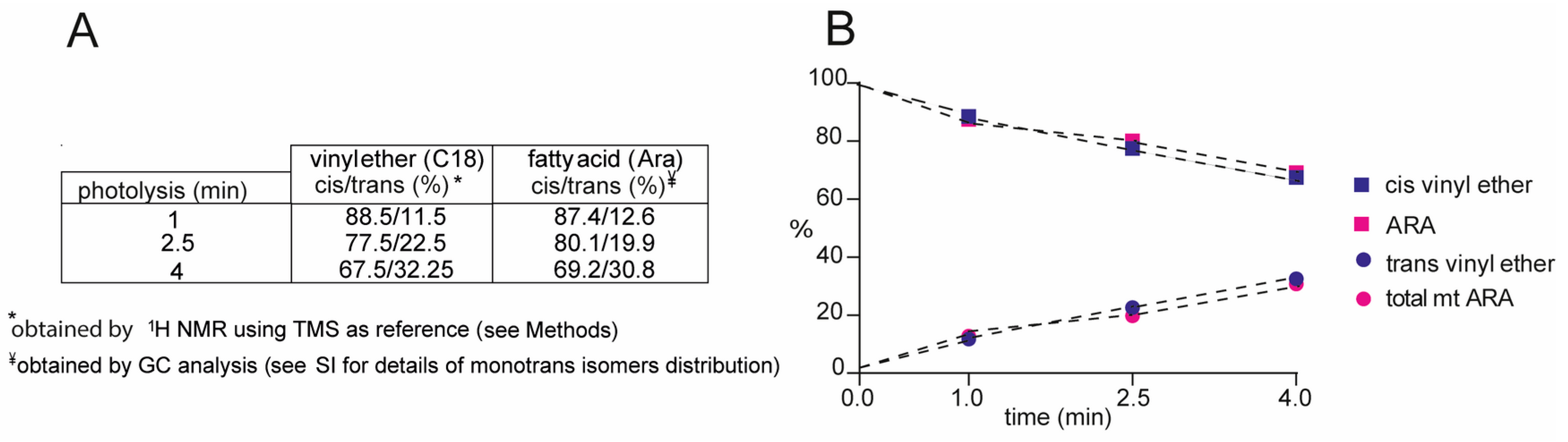
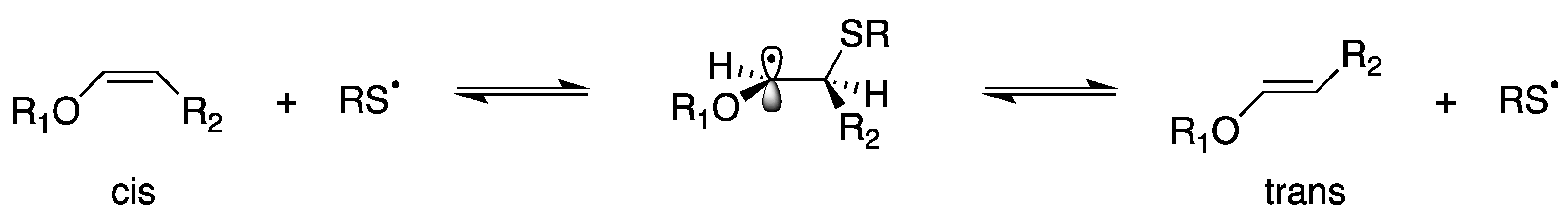
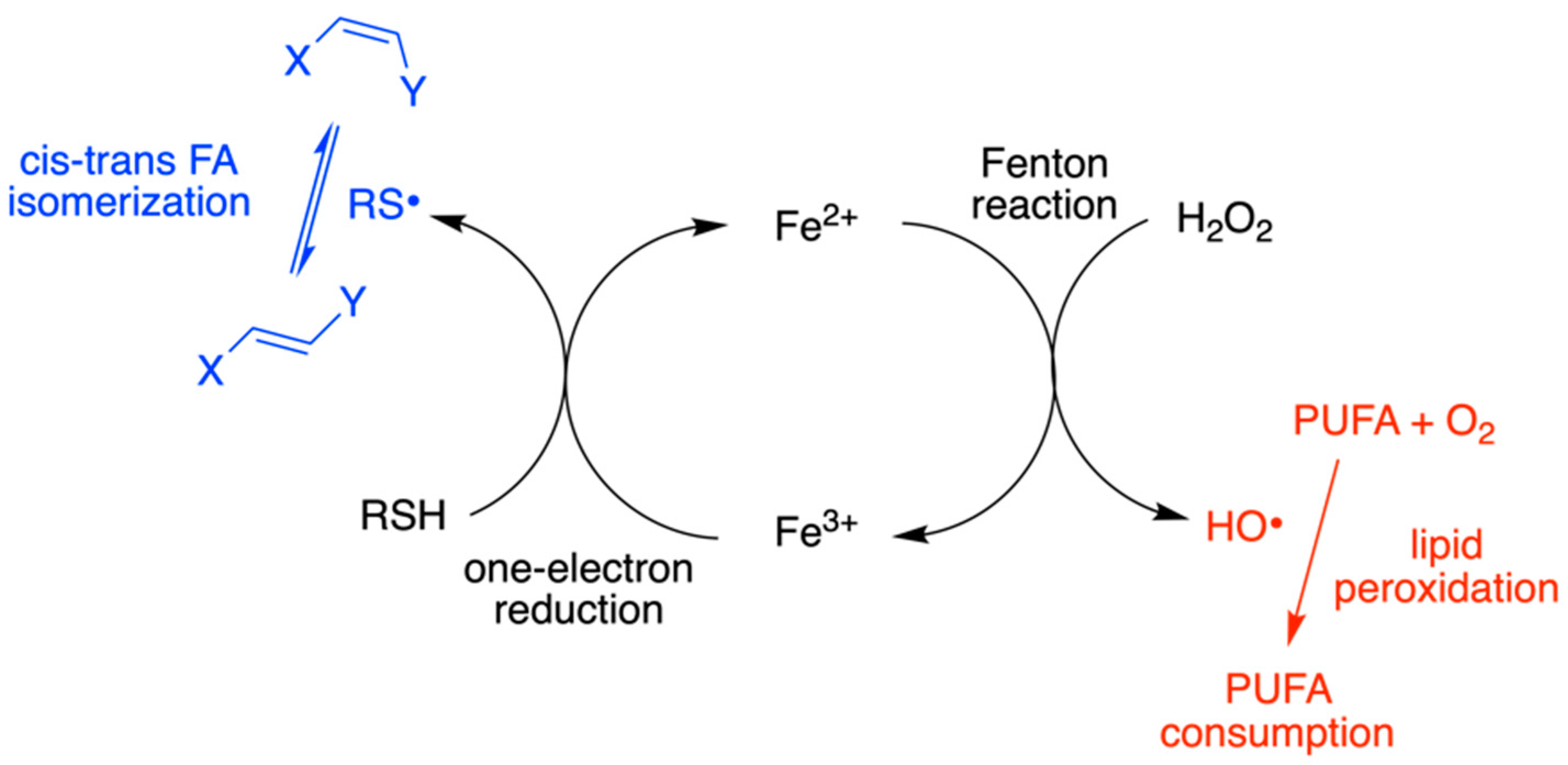
4.1. Plasmalogen Cis-Trans Isomerization in Solution
4.2. Transesterification Procedures for Plasmalogen-Containing Samples
4.3. Plasmalogen-Containing Liposomes as Model for Radical Stress Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flor, A.C.; Kron, S.J. Lipid-derived reactive aldehydes link oxidative stress to cell senescence. Cell Death Dis. 2016, 7, e2366. [Google Scholar] [CrossRef] [PubMed]
- Zhivaki, D.; Kagan, J.C. Innate immune detection of lipid oxidation as a threat assessment strategy. Nat. Rev. Immunol. 2022, 22, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Munir, R.; Lisec, J.; Swinnen, J.V.; Zaidi, N. Lipid metabolism in cancer cells under metabolic stress. Br. J. Cancer 2019, 120, 1090–1098. [Google Scholar] [CrossRef]
- Editorial. Let’s talk about lipid nanoparticles. Nat. Rev. Mater. 2021, 6, 99. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Skotland, T.; Sagini, K.; Sandvig, K.; Llorente, A. An emerging focus on lipids in extracellular vesicles. Adv. Drug Deliv. Rev. 2020, 159, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Koivuniemi, A. The biophysical properties of plasmalogens originating from their unique molecular architecture. FEBS Lett. 2017, 591, 2700–2713. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Garcia, A.; Monera-Girona, A.J.; Pajares-Martinez, E.; Bastida-Martinez, E.; Perez-Castano, R.; Iniesta, A.A.; Fontes, M.; Padmanabhan, S.; Elias-Arnanz, M. A bacterial light response reveals an orphan desaturase for human plasmalogen synthesis. Science 2019, 366, 128–132. [Google Scholar] [CrossRef]
- Wainberg, M.; Kamber, R.A.; Balsubramani, A.; Meyers, R.M.; Sinnott-Armstrong, N.; Hornburg, D.; Jiang, L.H.; Chan, J.; Jian, R.Q.; Gu, M.X.; et al. A genome-wide atlas of co-essential modules assigns function to uncharacterized genes. Nat. Genet. 2021, 53, 638–649. [Google Scholar] [CrossRef]
- Almsherqi, Z.A. Potential Role of Plasmalogens in the Modulation of Biomembrane Morphology. Front. Cell Dev. Biol. 2021, 9, 673917. [Google Scholar] [CrossRef] [PubMed]
- Honsho, M.; Abe, Y.; Fujiki, Y. Plasmalogen biosynthesis is spatiotemporally regulated by sensing plasmalogens in the inner leaflet of plasma membranes. Sci. Rep. 2017, 7, 43936. [Google Scholar] [CrossRef]
- Messias, M.C.F.; Mecatti, G.C.; Priolli, D.G.; Carvalho, P.D. Plasmalogen lipids: Functional mechanism and their involvement in gastrointestinal cancer. Lipids Health Dis. 2018, 17, 41. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta-Mol. Basis Dis. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Broniec, A.; Klosinski, R.; Pawlak, A.; Wrona-Krol, M.; Thompson, D.; Sarna, T. Interactions of plasmalogens and their diacyl analogs with singlet oxygen in selected model systems. Free Radic. Biol. Med. 2011, 50, 892–898. [Google Scholar] [CrossRef]
- Sindelar, P.J.; Guan, Z.Z.; Dallner, G.; Ernster, L. The protective role of plasmalogens in iron-induced lipid peroxidation. Free Radic. Biol. Med. 1999, 26, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Bueno, A.A.; Brand, A.; Neville, M.M.; Lehane, C.; Brierley, N.; Crawford, M.A. Erythrocyte phospholipid molecular species and fatty acids of Down syndrome children compared with non-affected siblings. Br. J. Nutr. 2015, 113, 72–81. [Google Scholar] [CrossRef]
- Dorninger, F.; Forss-Petter, S.; Berger, J. From peroxisomal disorders to common neurodegenerative diseases the role of ether phospholipids in the nervous system. FEBS Lett. 2017, 591, 2761–2788. [Google Scholar] [CrossRef]
- Dorninger, F.; Forss-Petter, S.; Wimmer, I.; Berger, J. Plasmalogens, platelet-activating factor and beyond—Ether lipids in signaling and neurodegeneration. Neurobiol. Dis. 2020, 145, 105061. [Google Scholar] [CrossRef]
- Paul, S.; Lancaster, G.I.; Meikle, P.J. Plasmalogens: A potential therapeutic target for neurodegenerative and cardiometabolic disease. Prog. Lipid Res. 2019, 74, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Maeba, R.; Maeda, T.; Kinoshita, M.; Takao, K.; Takenaka, H.; Kusano, J.; Yoshimura, N.; Takeoka, Y.; Yasuda, D.; Okazaki, T.; et al. Plasmalogens in human serum positively correlate with high-density lipoprotein and decrease with aging. J. Atheroscler. Thromb. 2007, 14, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Oberg, T.S.; Ward, R.E.; Steele, J.L.; Broadbent, J.R. Identification of plasmalogens in the cytoplasmic membrane of Bifidobacterium animalis subsp. lactis. Appl. Environ. Microbiol. 2012, 78, 880–884. [Google Scholar] [CrossRef]
- Pike, D.P.; McGuffee, R.M.; Geerling, E.; Albert, C.J.; Hoft, D.F.; Shashaty, M.G.S.; Meyer, N.J.; Pinto, A.K.; Ford, D.A. Plasmalogen loss in sepsis and SARS-CoV-2 infection. Front. Cell Dev. Biol. 2022, 10, 912880. [Google Scholar] [CrossRef] [PubMed]
- Acar, N.; Berdeaux, O.; Gregoire, S.; Cabaret, S.; Martine, L.; Gain, P.; Thuret, G.; Creuzot-Garcher, C.P.; Bron, A.M.; Bretillon, L. Lipid composition of the human eye: Are red blood cells a good mirror of retinal and optic nerve fatty acids? PLoS ONE 2012, 7, e35102. [Google Scholar] [CrossRef] [PubMed]
- Pallot, C.; Mazzocco, J.; Meillon, C.; Semama, D.S.; Chantegret, C.; Ternoy, N.; Martin, D.; Donier, A.; Gregoire, S.; Creuzot-Garcher, C.P.; et al. Alteration of erythrocyte membrane polyunsaturated fatty acids in preterm newborns with retinopathy of prematurity. Sci. Rep. 2019, 9, 7930. [Google Scholar] [CrossRef] [PubMed]
- Lankalapalli, R.S.; Eckelkamp, J.T.; Sircar, D.; Ford, D.A.; Subbaiah, P.V.; Bittman, R. Synthesis and antioxidant properties of an unnatural plasmalogen analogue bearing a trans O-vinyl ether linkage. Org. Lett. 2009, 11, 2784–2787. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Ferreri, C.; Melchiorre, M.; Sansone, A.; Torreggiani, A. Lipid geometrical isomerism: From chemistry to biology and diagnostics. Chem. Rev. 2014, 114, 255–284. [Google Scholar] [CrossRef]
- Cort, A.; Ozben, T.; Sansone, A.; Barata-Vallejo, S.; Chatgilialoglu, C.; Ferreri, C. Bleomycin-induced trans lipid formation in cell membranes and in liposome models. Org. Biomol. Chem. 2015, 13, 1100–1105. [Google Scholar] [CrossRef]
- Ferreri, C.; Kratzsch, S.; Brede, O.; Marciniak, B.; Chatgilialoglu, C. Trans lipids formation induced by thiols in human monocytic leukemia cells. Free Radic. Biol. Med. 2005, 38, 1180–1187. [Google Scholar] [CrossRef]
- Zambonin, L.; Ferreri, C.; Cabrini, L.; Prata, C.; Chatgilialoglu, C.; Landi, L. Occurrence of trans fatty acids in rats fed a trans-free diet: A free radical-mediated formation? Free Radic. Biol. Med. 2006, 40, 1549–1556. [Google Scholar] [CrossRef]
- Krokidis, M.G.; Prasinou, P.; Efthimiadou, E.K.; Boari, A.; Ferreri, C.; Chatgilialoglu, C. Effects of Aging and Disease Conditions in Brain of Tumor-Bearing Mice: Evaluation of Purine DNA Damages and Fatty Acid Pool Changes. Biomolecules 2022, 12, 1075. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Mennella, M.R.F.; Formisano, C.; Landi, L.; Chatgilialoglu, C. Arachidonate geometrical isomers generated by thiyl radicals: The relationship with trans lipids detected in biological samples. Free Radic. Biol. Med. 2002, 33, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Del Duca, E.; Sansone, A.; Sgrulletti, M.; Di Nolfo, F.; Chini, L.; Ferreri, C.; Moschese, V. Fatty-acid-based membrane lipidome profile of peanut allergy patients: An exploratory study of a lifelong health condition. Int. J. Mol. Sci. 2023, 24, 120. [Google Scholar] [CrossRef] [PubMed]
- Vetica, F.; Sansone, A.; Meliota, C.; Batani, G.; Roberti, M.; Chatgilialoglu, C.; Ferreri, C. Free-radical-mediated formation of trans-cardiolipin isomers, analytical approaches for lipidomics and consequences of the structural organization of membranes. Biomolecules 2020, 10, 1189. [Google Scholar] [CrossRef]
- Tartaro-Bujak, I.; Mihaljević, B.; Ferreri, C.; Chatgilialoglu, C. The influence of antioxidants in the thiyl radical induced lipid peroxidation and geometrical isomerization in micelles of linoleic acid. Free Radic. Res. 2016, 50, S18–S23. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Yu, N.; Zhao, J.; Xie, Z.M.; Yang, Z.N.; Tian, B. Advances in the biosynthetic pathways and application potential of plasmalogens in medicine. Front. Cell Dev. Biol. 2020, 8, 765. [Google Scholar] [CrossRef]
- Kramer, J.F.K.; Fellner, V.; Dugan, M.E.R.; Sauer, F.D.; Mossoba, M.M.; Yurawecz, M.P. Evaluating acid and base catalysts in the methylation of milk and rumen fatty acids with special emphasis on conjugated dienes and total trans fatty acids. Lipids 1997, 32, 1219–1228. [Google Scholar] [CrossRef]
- Dong, J.L.; Yu, L.S.H.; Xie, J.W. A simple and versatile method for the formation of acetals/ketals using trace conventional acids. Acs Omega 2018, 3, 4974–4985. [Google Scholar] [CrossRef]
- Hanus, L.O.; Levitsky, D.O.; Shkrob, I.; Dembitsky, V.M. Plasmalogens, fatty acids and alkyl glyceryl ethers of marine and freshwater clams and mussels. Food Chem. 2009, 116, 491–498. [Google Scholar] [CrossRef]
- Morrison, W.R.; Smith, L.M. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride-methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Sansone, A.; Tolika, E.; Louka, M.; Sunda, V.; Deplano, S.; Melchiorre, M.; Anagnostopoulos, D.; Chatgilialoglu, C.; Formisano, C.; Di Micco, R.; et al. Hexadecenoic fatty acid isomers in human blood lipids and their relevance for the interpretation of lipidomic profiles. PLoS ONE 2016, 11, 152378. [Google Scholar] [CrossRef] [PubMed]
- Sansone, A.; Melchiorre, M.; Chatgilialoglu, C.; Ferreri, C. Hexadecenoic fatty acid isomers: A chemical biology approach for human plasma biomarker development. Chem. Res. Toxicol. 2013, 26, 1703–1709. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.Y.; Wang, L.; Liu, J.; Ma, K.L.; Liu, C.P.; Zhang, Y.; Zou, W. Choline Plasmalogens Isolated from Swine Liver Inhibit Hepatoma Cell Proliferation Associated with Caveolin-1/Akt Signaling. PLoS ONE 2013, 8, 77387. [Google Scholar] [CrossRef]
- Ferreri, C.; Samadi, A.; Sassatelli, F.; Landi, L.; Chatgilialoglu, C. Regioselective cis-trans isomerization of arachidonic double bonds by thiyl radicals: The influence of phospholipid supramolecular organization. J. Am. Chem. Soc. 2004, 126, 1063–1072. [Google Scholar] [CrossRef]
- Williams, C.M.; Mander, L.N. Chromatography with silver nitrate. Tetrahedron 2001, 57, 425–447. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Altieri, A.; Fischer, H. The kinetics of thiyl radical induced reactions of monounsaturated fatty acid esters. J. Am. Chem. Soc. 2002, 124, 12816–12823. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Samadi, A.; Guerra, M.; Fischer, H. The kinetics of Z/E isomerization of methyl oleate, catalyzed by photogenerated thiyl radicals. ChemPhysChem 2005, 6, 286–291. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Bowry, V.W. Why not trans? Inhibited radical isomerization cycles and coupling chains of lipids and alkenes with alkane-thiols. J. Org. Chem. 2018, 83, 9178–9189. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Ferreri, C.; Guerra, M.; Samadi, A.; Bowry, V.W. The reaction of thiyl radical with methyl linoleate: Completing the picture. J. Am. Chem. Soc. 2017, 139, 4704–4714. [Google Scholar] [CrossRef]
- Chatgilialoglu, C.; Studer, A. (Eds.) Encyclopedia of Radicals in Chemistry, Biology and Materials; Wiley: Chichester, UK, 2012. [Google Scholar]
- Fife, T.H. Vinyl ether hydrolysis. The facile general acid catalyzed conversion of 2-ethoxy-1-cyclopentene-1-carboxylic acid to cyclopentanone. J. Am. Chem. Soc. 1965, 87, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Alves, S.P.; Santos-Silva, J.; Cabrita, A.R.J.; Fonseca, A.J.M.; Bessa, R.J.B. Detailed dimethylacetal and fatty acid composition of rumen content from lambs fed lucerne or concentrate supplemented with soybean oil. PLoS ONE 2013, 8, 58386. [Google Scholar] [CrossRef] [PubMed]
- See the LIPID MAPS library. Available online: https://www.lipidmaps.org/resources/lipidweb/lipidweb_html/ms/others/misclipids/index.htm (accessed on 18 April 2022).
- Delmonte, P.; Belaunzaran, X.; Ridge, C.D.; Aldai, N.; Kramer, J.K.G. Separation and characterization of products from acidic methanolysis of plasmalogenic lipids by two-dimensional gas chromatography with online reduction. J. Chromatogr. A 2020, 1619, 460955. [Google Scholar] [CrossRef] [PubMed]
- Otoki, Y.; Hennebelle, M.; Levitt, A.J.; Nakagawa, K.; Swardfager, W.; Taha, A.Y. Plasma Phosphatidylethanolamine and triacylglycerol fatty acid concentrations are altered in major depressive disorder patients with seasonal pattern. Lipids 2017, 52, 559–571. [Google Scholar] [CrossRef]
- Aldai, N.; Murray, B.E.; Najera, A.I.; Troy, D.J.; Osoro, K. Derivatization of fatty acids and its application for conjugated linoleic acid studies in ruminant meat lipids. J. Sci. Food Agric. 2005, 85, 1073–1083. [Google Scholar] [CrossRef]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef]
- Hame, K.; Fujiwara, Y.; Yokoyama, K. Quantitative and qualitative method for sphingomyelin by LC-MS using two stable isotopically labeled sphingomyelin species. J. Vis. Exp. 2018, 135, e57293. [Google Scholar] [CrossRef]
- Devle, H.; Naess-Andresen, C.F.; Stenstrom, Y.; Ekeberg, D. Rapid method for analysis of sphingomyelin by microwave derivatisation for gas chromatography-mass spectrometry. Eur. J. Lipid Sci. Technol. 2011, 113, 708–710. [Google Scholar] [CrossRef]
- Takashima, S.; Toyoshi, K.; Itoh, T.; Kajiwara, N.; Honda, A.; Ohba, A.; Takemoto, S.; Yoshida, S.; Shimozawa, N. Detection of unusual very-long-chain fatty acid and ether lipid derivatives in the fibroblasts and plasma of patients with peroxisomal diseases using liquid chromatography-mass spectrometry. Mol. Genet. Metab. 2017, 120, 255–268. [Google Scholar] [CrossRef]
- Wang, L.Y.; Summerhill, K.; Rodriguez-Canas, C.; Mather, I.; Patel, P.; Eiden, M.; Young, S.; Forouhi, N.G.; Koulman, A. Development and validation of a robust automated analysis of plasma phospholipid fatty acids for metabolic phenotyping of large epidemiological studies. Genome Med. 2013, 5, 39. [Google Scholar] [CrossRef]
- Metherel, A.H.; Henao, J.J.A.; Ciobanu, F.; Taha, A.Y.; Stark, K.D. Microwave energy increases fatty acid methyl ester yield in human whole blood due to increased sphingomyelin transesterification. Lipids 2015, 50, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.-L.; Hwang, L.S.; Shahidi, F.; Pan, M.-H.; Wang, Y.; Ho, C.-T. Endogenous formation of trans fatty acids: Health implications and potential dietary intervention. J. Funct. Foods 2016, 25, 14–24. [Google Scholar] [CrossRef]
- Brosche, T.; Platt, D. Decrease of cholesterol concentration in human erythrocyte-ghosts in old-age. Exp. Gerontol. 1990, 25, 23–28. [Google Scholar] [CrossRef]
- Zhang, X.X.; Barraza, K.M.; Beauchamp, J.L. Cholesterol provides nonsacrificial protection of membrane lipids from chemical damage at air-water interface. Proc. Natl. Acad. Sci. USA 2018, 115, 3255–3260. [Google Scholar] [CrossRef] [PubMed]
- Elbaradei, A.; Wang, Z.K.; Malmstadt, N. Oxidation of membrane lipids alters the activity of the human serotonin 1A receptor. Langmuir 2022, 38, 6798–6807. [Google Scholar] [CrossRef]
- Kagan, V.E.; Mao, G.W.; Qu, F.; Angeli, J.P.F.; Doll, S.; St Croix, C.; Dar, H.H.; Liu, B.; Tyurin, V.A.; Ritov, V.B.; et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat. Chem. Biol. 2017, 13, 81–90. [Google Scholar] [CrossRef]
- Chen, X.; Yu, C.H.; Kang, R.; Tang, D.L. Iron metabolism in ferroptosis. Front. Cell Dev. Biol. 2020, 8, 590226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAME 1 | KOH/MeOH (% Rel. Quant.) Mean ± SD (n = 10) | HCl/MeOH (% Rel. Quant.) Mean ± SD (n = 10) 2 |
|---|---|---|
| 16:0 | 23.3 ± 0.6 | 34.0 ± 3.2 *** |
| 9cis-16:1 | 0.24 ± 0.03 | 0.18 ± 0.05 ** |
| 18:0 | 16.0 ± 1.3 | 21.8 ± 1.3 *** |
| 9trans-18:1 | 0.01 ± 0.01 | 0.03 ± 0.01 *** |
| 9cis-18:1 | 17.0 ± 0.5 | 13.8 ± 1.0 *** |
| 11cis-18:1 | 1.2 ± 0.2 | 1.0 ± 0.1 *** |
| 9cis,12cis-18:2 | 12.1±0.6 | 9.2 ± 0.4 *** |
| 20:3 omega-6, DGLA | 2.3 ± 0.1 | 1.5 ± 0.3 *** |
| 20:4 omega-6, Ara | 18.8 ± 0.4 | 13.3 ± 1.4 *** |
| 5mtAra | 0.02 ± 0.01 | 0.03 ± 0.01 * |
| 20:5 omega-3, EPA | 0.8 ± 0.1 | 0.6 ± 0.3 * |
| 22:5 omega-3, DPA | 2.7 ± 0.5 | 1.7 ± 0.4 *** |
| 22:6 omega-3, DHA | 5.4 ± 0.1 | 3.0 ± 0.5 *** |
| SFA | 39.3 ± 1.3 | 55.8 ± 3.3 *** |
| MUFA | 18.5 ± 0.7 | 14.9 ± 0.9 *** |
| PUFA | 42.1 ± 0.7 | 29.3 ± 2.6 *** |
| omega-6 | 33.2 ± 0.7 | 24.0 ± 1.8 *** |
| omega-3 | 8.9 ± 0.4 | 5.2 ± 0.9 *** |
| TOT TRANS | 0.03 ± 0.01 | 0.05 ± 0.02 * |
| Thiol (μM) | 9cis-18:1 (%) | 9trans-18:1 (%) | Ara (%) | Ara Loss (%) | mtAra (%) |
|---|---|---|---|---|---|
| 0 | 99.9 | - | 54.0 | 46.0 | n.d. |
| 10 | 99.0 | 0.55 | 77.1 | 21.0 | 1.9 |
| Thiol (μM) | 9cis-18:1 | 9trans-18:1 | Ara | Ara Loss | mtAra | 18:2 | 18:2 Loss | mt-18:2 |
|---|---|---|---|---|---|---|---|---|
| 0 | 99.0 ± 1.0 | - | 66.9 ± 1.1 | 33.1 ± 1.1 | - | 73.4 ± 2.1 | 26.5 ± 2.1 | - |
| 10 | 99.2 ± 0.1 | 0.7 ± 0.1 | 95.0 ± 0.2 | 2.3 ± 0.01 | 2.3 ± 0.2 | 96.1 ± 0.8 | 2.5 ± 0.6 | 1.4 ± 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreri, C.; Ferocino, A.; Batani, G.; Chatgilialoglu, C.; Randi, V.; Riontino, M.V.; Vetica, F.; Sansone, A. Plasmalogens: Free Radical Reactivity and Identification of Trans Isomers Relevant to Biological Membranes. Biomolecules 2023, 13, 730. https://doi.org/10.3390/biom13050730
Ferreri C, Ferocino A, Batani G, Chatgilialoglu C, Randi V, Riontino MV, Vetica F, Sansone A. Plasmalogens: Free Radical Reactivity and Identification of Trans Isomers Relevant to Biological Membranes. Biomolecules. 2023; 13(5):730. https://doi.org/10.3390/biom13050730
Chicago/Turabian StyleFerreri, Carla, Alessandra Ferocino, Gessica Batani, Chryssostomos Chatgilialoglu, Vanda Randi, Maria Vittoria Riontino, Fabrizio Vetica, and Anna Sansone. 2023. "Plasmalogens: Free Radical Reactivity and Identification of Trans Isomers Relevant to Biological Membranes" Biomolecules 13, no. 5: 730. https://doi.org/10.3390/biom13050730