
The Cornea: No Difference in the Wound Healing Response to Injury Related to Whether, or Not, There’s a Bowman’s Layer


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
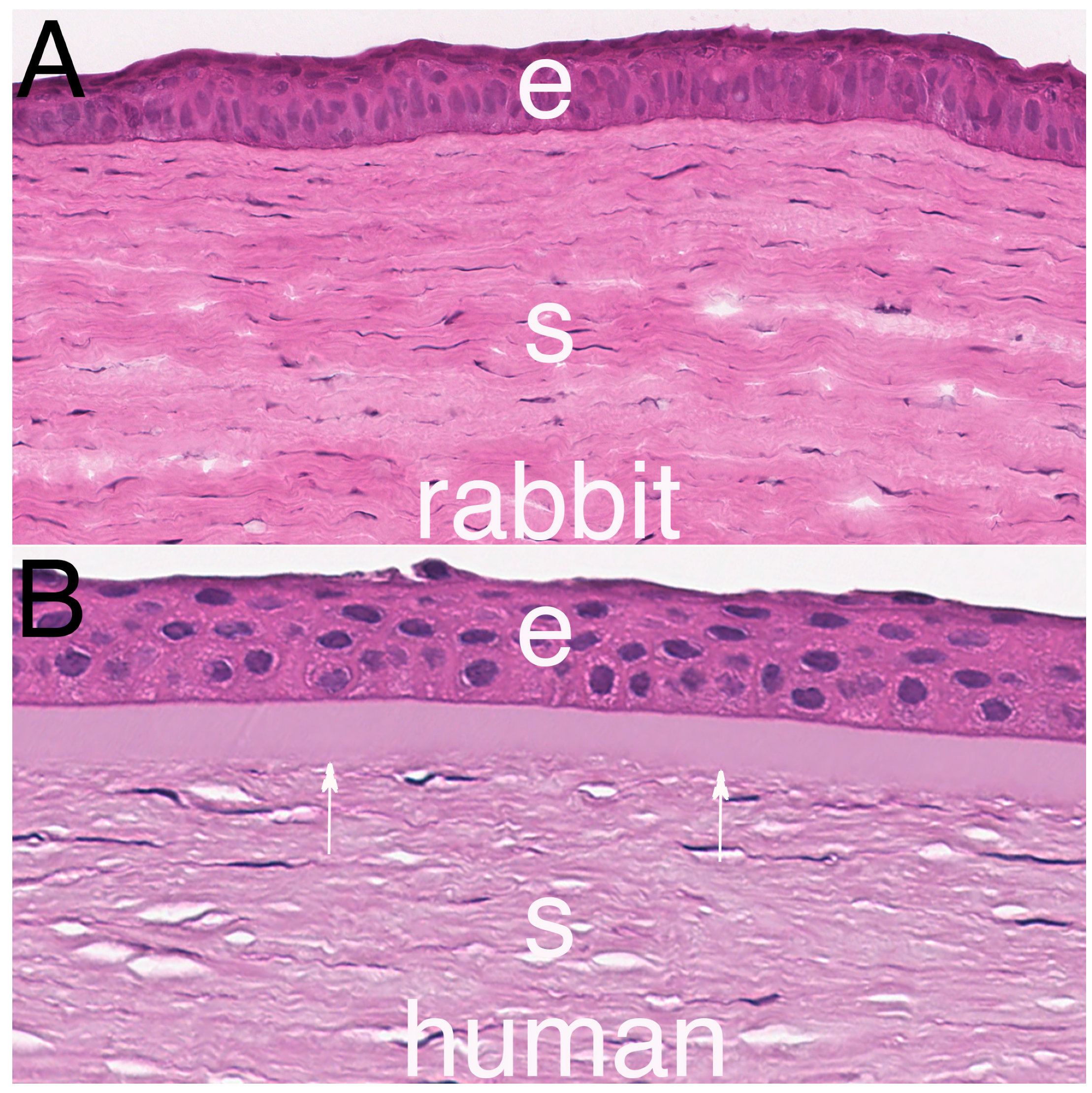
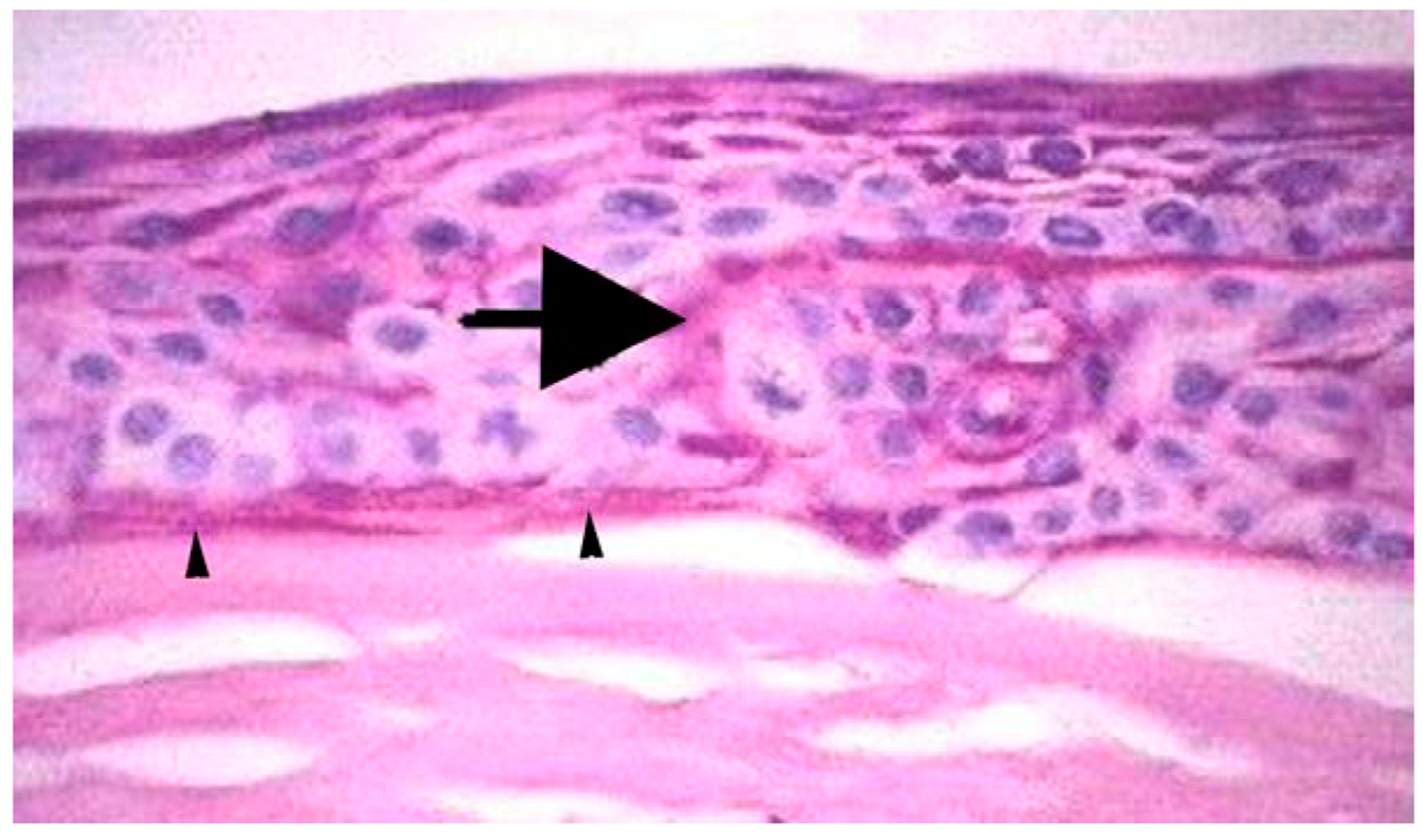
2. Bowman’s Layer Structure
3. Bowman’s Layer Function

4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Streeten, B.W.; Qi, Y.; Klintworth, G.K.; Eagle, R.C., Jr.; Strauss, J.A.; Bennett, K. Immunolocalization of beta Ig-h3 protein in 5q31-linked corneal dystrophies and normal corneas. Arch. Ophthalmol. 1999, 117, 67. [Google Scholar] [CrossRef]
- Hayashi, S.; Osawa, T.; Tohyama, K. Comparative observations on corneas, with special reference to Bowman’s layer and Descemet’s membrane in mammals and amphibians. J. Morphol. 2002, 254, 247–258. [Google Scholar] [CrossRef]
- Gealy, C.; Hayes, A.J.; Buckwell, R.; Young, R.D.; Caterson, B.; Quantock, A.J.; Ralphs, J.R. Actin and type I collagen propeptide distribution in the developing chick cornea. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Marchant, J.K.; Zhang, G.; Birk, D.E. Association of type XII collagen with regions of increased stability and keratocyte density in the cornea. Exp. Eye Res. 2002, 75, 683–694. [Google Scholar] [CrossRef]
- Nautscher, N.; Bauer, A.; Steffl, M.; Amselgruber, W.M. Comparative morphological evaluation of domestic animal cornea. Vet. Ophthalmol. 2016, 19, 297–304. [Google Scholar] [CrossRef]
- Miller, S.N.; Colitz, C.M.; Dubielzig, R.R. Anatomy of the California sea lion globe. Vet. Ophthalmol. 2010, 13 (Suppl. l), 63–71. [Google Scholar] [CrossRef]
- Gonçalves, G.C.; Pérez-Merino, P.; Martínez-García, M.C.; Barcía, A.; Merayo-Loves, J. Comparison of the characteristics in hen and quail corneas as experimental models of refractive surgery. Arch. Soc. Esp. Oftalmol. 2016, 91, 310–315. [Google Scholar] [CrossRef]
- Merayo-Lloves, J.; Yáñez, B.; Mayo, A.; Martín, R.; Pastor, J.C. Experimental model of corneal haze in chickens. J. Refract. Surg. 2001, 17, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Cafaro, T.A.; Ortiz, S.G.; Maldonado, C.; Espósito, F.A.; Croxatto, J.O.; Berra, A.; Ale, O.L.; Torrealday, J.I.; Urrets-Zavalía, E.A.; Urrets-Zavalía, J.A.; et al. The cornea of guinea pig: Structural and functional studies. Vet. Ophthalmol. 2009, 12, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Merindano, M.D.; Costa, J.; Canals, M.; Potau, J.M.; Ruano, D. A comparative study of Bowman’s layer in some mammals: Relationships with other constituent corneal structures. Eur. J. Anat. 2002, 6, 133–139. [Google Scholar]
- Wilson, S.E.; He, Y.-G.; Weng, J.; Li, Q.; McDowall, A.W.; Vital, M.; Chwang, E.L. Epithelial injury induces keratocyte apoptosis: Hypothesized role for the interleukin-1 system in the modulation of corneal tissue organization and wound healing. Exp. Eye Res. 1996, 62, 325–328. [Google Scholar] [CrossRef]
- Wilson, S.E. Bowman’s layer in the cornea—Structure and function and regeneration. Exp. Eye Res. 2020, 195, 108033. [Google Scholar] [CrossRef]
- Dewitt, E.N. The histopathology of Bowman’s membrane. Trans. Am. Ophthalmol. Soc. 1931, 29, 461–485. [Google Scholar]
- Germundsson, J.; Karanis, G.; Fagerholm, P.; Lagali, N. Age-related thinning of Bowman’s layer in the human cornea in vivo. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6143–6149. [Google Scholar] [CrossRef]
- Kermani, O.; Fabian, W.; Lubatschowski, H. Real-time optical coherence tomography-guided femtosecond laser sub-Bowman keratomileusis on human donor eyes. Am. J. Ophthalmol. 2008, 146, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Li, H.F.; Petroll, W.M.; Moller-Pedersen, T.; Maurer, J.K.; Cavanagh, H.D.; Jester, J.V. Epithelial and corneal thickness measurements by in vivo confocal microscopy through focusing (CMTF). Curr. Eye Res. 1997, 16, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, I.E.; Jensen, O.A.; Prause, J.U. Structure and composition of Bowman’s membrane. Study by frozen resin cracking. Acta Ophthalmol. 1984, 62, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, A.S.; Spurr-Michaud, S.J.; Rodrigues, M.; Hackett, J.; Krachmer, J.; Gipson, I.K. Development of the anchoring structures of the epithelium in rabbit and human fetal corneas. Investig. Ophthalmol. Vis. Sci. 1988, 29, 727–736. [Google Scholar]
- Bach, L.B.; Seefelder, R. Atlas Zur Entwicklungsgeschichte Des Menschlichen Auges; Wilhelm Engelmann Verlag: Leipzig, Germany, 1914. [Google Scholar]
- Ozanics, V.; Jakobiec, F.A. Biomedical foundations of Ophthalmology; Harper and Row: Philadelphia, CA, USA, 1982; pp. 8–14. [Google Scholar]
- Zimmermann, D.R.; Fischer, R.W.; Winterhalter, K.H.; Witmer, R.; Vaughan, L. Comparative studies of collagens in normal and keratoconus corneas. Exp. Eye Res. 1988, 46, 431–442. [Google Scholar] [CrossRef]
- Marshall, G.E.; Konstas, A.G.; Lee, W.R. Immunogold fine structural localization of extracellular matrix components in aged human cornea. I. Types I–IV collagen and laminin. Graefes Arch. Clin. Exp. Ophthalmol. 1991, 229, 157–163. [Google Scholar] [CrossRef]
- Marshall, G.E.; Konstas, A.G.; Lee, W.R. Immunogold fine structural localization of extracellular matrix components in aged human cornea. II. Collagen types V and VI. Graefes Arch. Clin. Exp. Ophthalmol. 1991, 229, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.K.; Foley, J.W.; Birk, D.E.; Fitch, J.M.; Linsenmayer, T.F. Type V collagen and Bowman’s membrane. Quantitation of mRNA in corneal epithelium and stroma. J. Biol. Chem. 1994, 269, 24959–24966. Available online: https://www.jbc.org/article/S0021-9258(17)31483-7/pdf (accessed on 15 February 2023). [CrossRef]
- Tong, C.M.; van Dijk, K.; Melles, G.R.J. Update on Bowman layer transplantation. Curr. Opin. Ophthalmol. 2019, 30, 249. [Google Scholar] [CrossRef]
- García de Oteyza, G.; González Dibildox, L.A.; Vázquez-Romo, K.A.; Tapia Vázquez, A.; Dávila Alquisiras, J.H.; Martínez-Báez, B.E.; García-Albisua, A.M.; Ramírez, M.; Hernández-Quintela, E. Bowman layer transplantation using a femtosecond laser. J. Cataract Refract. Surg. 2019, 45, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.; Hussain, I.; Borroni, D.; Khan, B.S.; Wahab, S.; Mahar, P.S. Bowman’s layer transplantation in advanced keratoconus; 18-months outcomes. Int. Ophthalmol. 2022, 42, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Walker, J.W.; Chwang, E.L.; He, Y.-G. Hepatocyte growth factor (HGF), keratinocyte growth factor (KGF), their receptors, FGF receptor-2, and the cells of the cornea. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2544–2561. [Google Scholar]
- Wilson, S.E.; He, Y.-G.; Weng, J.; Zieske, J.D.; Jester, J.V.; Schultz, G.S. Effect of epidermal growth factor, hepatocyte growth factor, and keratinocyte growth factor, on proliferation, motility, and differentiation of human corneal epithelial cells. Exp. Eye Res. 1994, 59, 665–678. [Google Scholar] [CrossRef]
- Wilson, S.E.; Li, Q.; Weng, J.; Barry-Lane, P.A.; Jester, J.V.; Liang, Q.; Wordinger, R.J. The Fas/Fas ligand system and other modulators of apoptosis in the cornea. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1582–1592. [Google Scholar]
- Mohan, R.R.; Liang, Q.; Kim, W.-J.; Helena, M.C.; Baerveldt, F.; Wilson, S.E. Apoptosis in the cornea: Further characterization of Fas-Fas ligand system. Exp. Eye Res. 1997, 65, 575–589. [Google Scholar] [CrossRef]
- de Oliveira, R.C.; Tye, G.; Sampaio, L.P.; Shiju, T.M.; Dedreu, J.; Menko, A.S.; Santhiago, M.R.; Wilson, S.E. TGF 1 and TGF 2 proteins in corneas with and without stromal fibrosis: Delayed regeneration of epithelial barrier function and the epithelial basement membrane in corneas with stromal fibrosis. Exp. Eye Res. 2021, 202, 108325. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.C.; Sampaio, L.P.; Shiju, T.M.; Santhiago, M.R.; Wilson, S.E. Epithelial basement membrane regeneration after PRK-induced epithelial-stromal injury in rabbits: Fibrotic vs. non-fibrotic corneal healing. J. Ref. Surg. 2022, 38, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-J.; Mohan, R.R.; Mohan, R.R.; Wilson, S.E. Effect of PDGF, IL-1 alpha, and BMP2/4 on corneal fibroblast chemotaxis: Expression of the platelet-derived growth factor system in the cornea. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1364–1372. [Google Scholar]
- Wilson, S.E. The Yin and Yang of mesenchymal cells in the corneal stromal fibrosis response to injury: The cornea as a model of fibrosis in other organs. Biomolecules 2022, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E. Defective perlecan-associated basement membrane regeneration and altered modulation of transforming growth factor beta in fibrosis. Cell. Molec. Life Sci. 2022, 79, 144. [Google Scholar] [CrossRef] [PubMed]
- Ritchey, E.R.; Code, K.; Zelinka, C.P.; Scott, M.A.; Fischer, A.J. The chicken cornea as a model of wound healing and neuronal re-innervation. Mol. Vis. 2011, 17, 2440–2454. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3185018/ (accessed on 15 February 2023).
- Fowler, W.C.; Chang, D.H.; Roberts, B.C.; Zarovnaya, E.L.; Proia, A.D. A new paradigm for corneal wound healing research: The white leghorn chicken (Gallus gallus domesticus). Curr. Eye Res. 2004, 28, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, M.C.; Merayo-Llovés, J.; Blanco-Mezquita, T.; Mar-Sardaña, S. Wound healing following refractive surgery in hens. Exp. Eye Res. 2006, 83, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Ambrósio, R., Jr.; Kara-José, N.; Wilson, S.E. Early keratocyte apoptosis after epithelial scrape injury in the human cornea. Exp. Eye Res. 2009, 89, 597–599. [Google Scholar] [CrossRef]
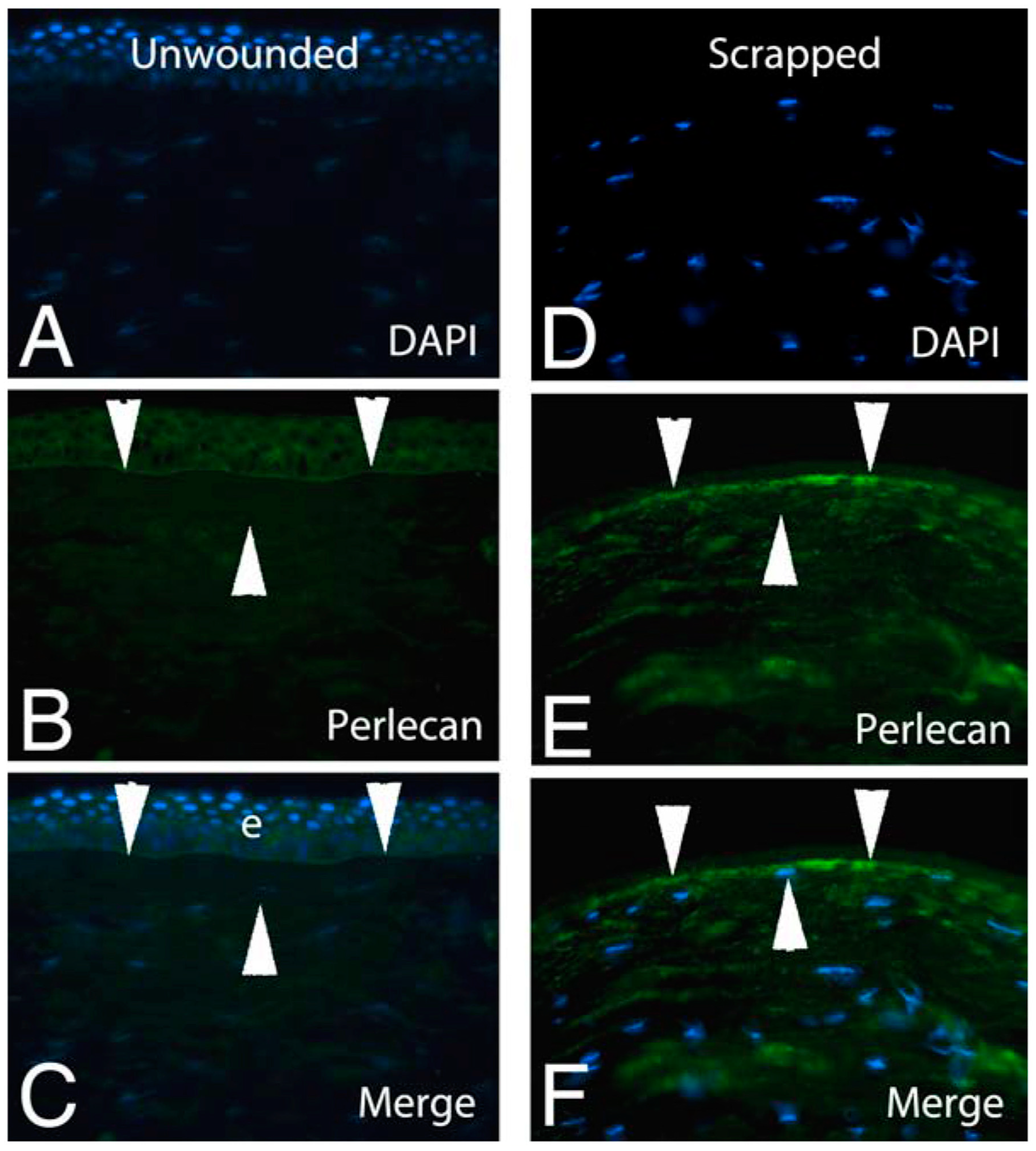
- Torricelli, A.A.M.; Marino, G.K.; Santhanam, A.; Wu, J.; Singh, A.; Wilson, S.E. Epithelial basement membrane proteins perlecan and nidogen-2 are up-regulated in stromal cells after epithelial injury in human corneas. Exp. Eye Res. 2015, 134, 33–38. [Google Scholar] [CrossRef]
- Santhanam, A.; Marino, G.K.; Torricelli, A.A.M.; Wilson, S.E. EBM regeneration and changes in EBM component mRNA expression in the anterior stroma after corneal injury. Mol. Vision. 2017, 23, 39–51. [Google Scholar]
- Lipshitz, I.; Loewenstein, A.; Varssano, D.; Lazar, M. Late onset corneal haze after photorefractive keratectomy for moderate and high myopia. Ophthalmology 1997, 104, 369–373. [Google Scholar] [CrossRef]
- Mohan, R.R.; Hutcheon, A.E.K.; Choi, R.; Hong, J.-W.; Lee, J.-S.; Mohan, R.R.; Ambrósio, R., Jr.; Zieske, J.D.; Wilson, S.E. Apoptosis, necrosis, proliferation, and myofibroblast generation in the stroma following LASIK and PRK. Exp. Eye Res. 2003, 76, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.R.; Stapleton, W.M.; Sinha, S.; Netto, M.V.; Wilson, S.E. A novel method for generating corneal haze in anterior stroma of the mouse eye with the excimer laser. Exp. Eye Res. 2008, 86, 235–240. [Google Scholar] [CrossRef]
- Singh, V.; Torricelli, A.A.M.; Nayeb-Hashemi, N.; Agrawal, V.; Wilson, S.E. Mouse strain variation in SMA+ myofibroblast development after corneal injury. Exp. Eye Res. 2013, 115, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Hong, J.W. Bowman’s layer structure and function: Critical or dispensable to corneal function? A hypothesis. Cornea 2000, 19, 417–420. [Google Scholar] [CrossRef]
- Wilson, S.E.; Bourne, W.M. Fuchs’ dystrophy. Cornea 1988, 7, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Melles, G.R.; Binder, P.S.; Moore, M.N.; Anderson, J.A. Epithelial-stromal interactions in human keratotomy wound healing. Arch. Ophthalmol. 1995, 113, 1124–1130. [Google Scholar] [CrossRef]
- Marino, G.K.; Santhiago, M.R.; Santhanam, A.; Torricelli, A.A.M.; Wilson, S.E. Regeneration of defective epithelial basement membrane and restoration of corneal transparency after photorefractive keratectomy. J. Refract. Surg. 2017, 33, 337–346. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, S.E. The Cornea: No Difference in the Wound Healing Response to Injury Related to Whether, or Not, There’s a Bowman’s Layer. Biomolecules 2023, 13, 771. https://doi.org/10.3390/biom13050771
Wilson SE. The Cornea: No Difference in the Wound Healing Response to Injury Related to Whether, or Not, There’s a Bowman’s Layer. Biomolecules. 2023; 13(5):771. https://doi.org/10.3390/biom13050771
Chicago/Turabian StyleWilson, Steven E. 2023. "The Cornea: No Difference in the Wound Healing Response to Injury Related to Whether, or Not, There’s a Bowman’s Layer" Biomolecules 13, no. 5: 771. https://doi.org/10.3390/biom13050771