
Collagen Crosslinking for Keratoconus: Cellular Signaling Mechanisms


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics and Donor Information
2.2. Cell Isolation and 3D Self-Assembled Constructs
2.3. Collagen Crosslinking (CXL)—In Vitro
2.4. Protein Isolation and Western Blots
2.5. Statistical Analysis
3. Results
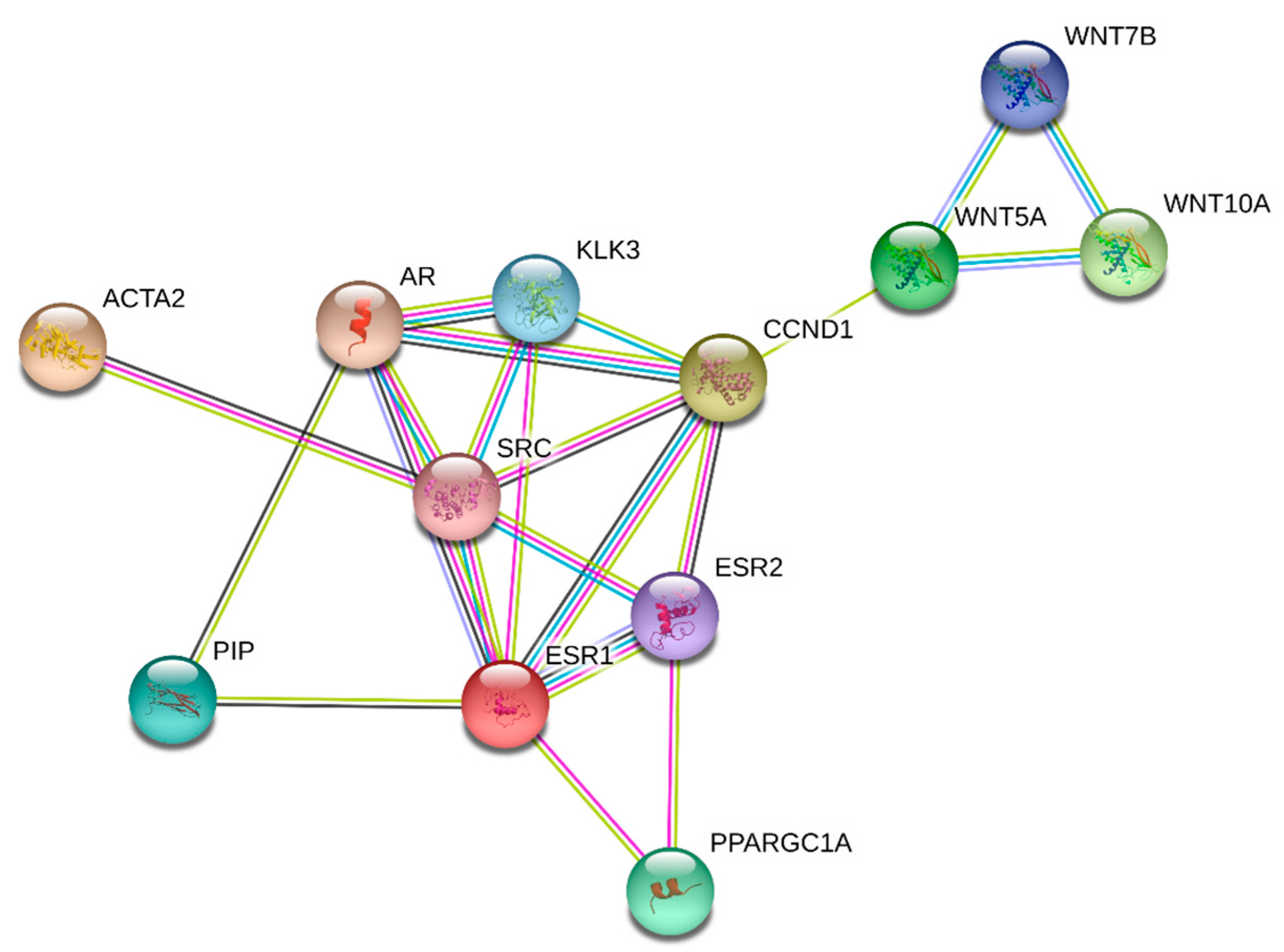
3.1. Signaling Pathways
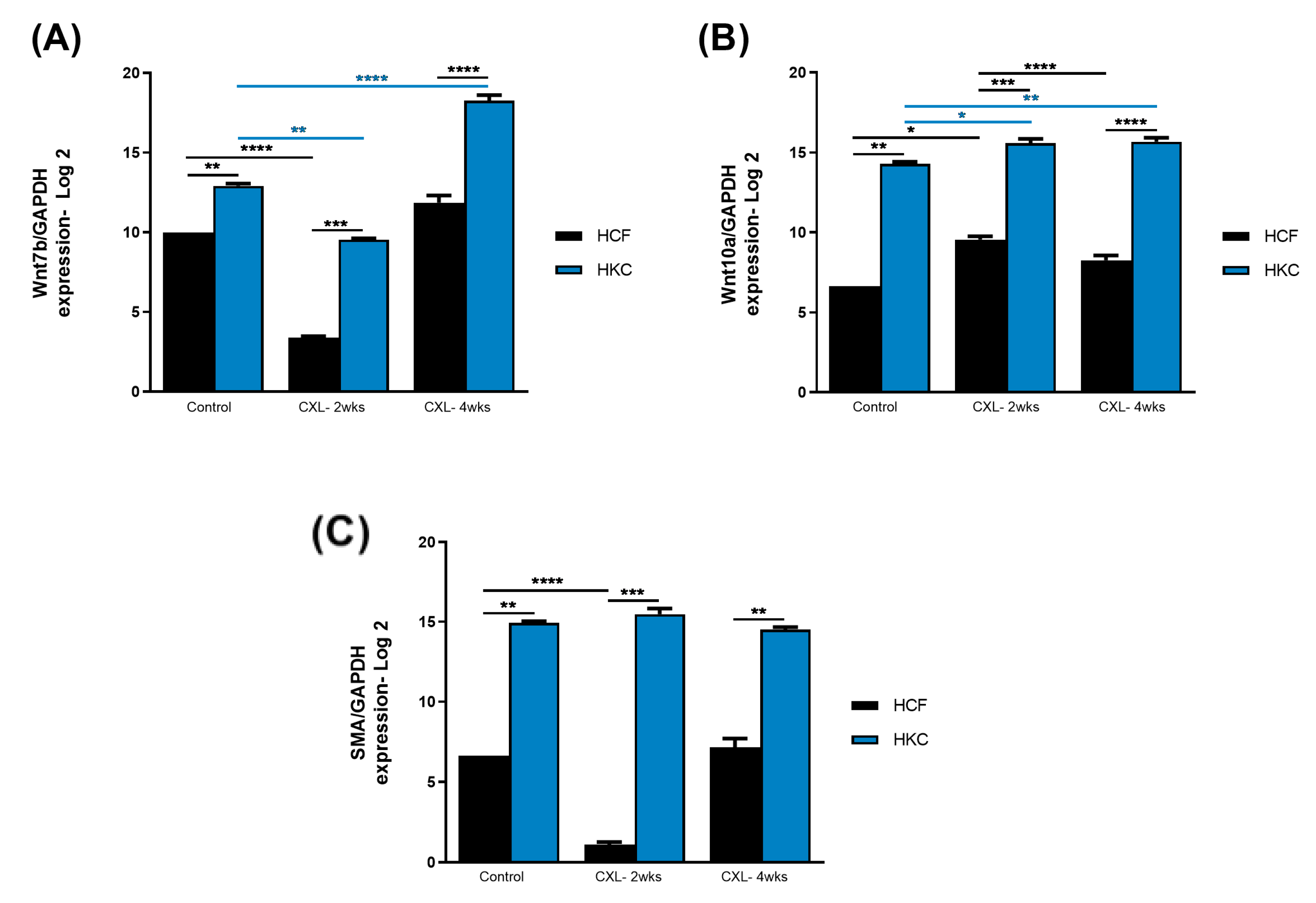
3.2. Wnt Signaling and Fibrosis
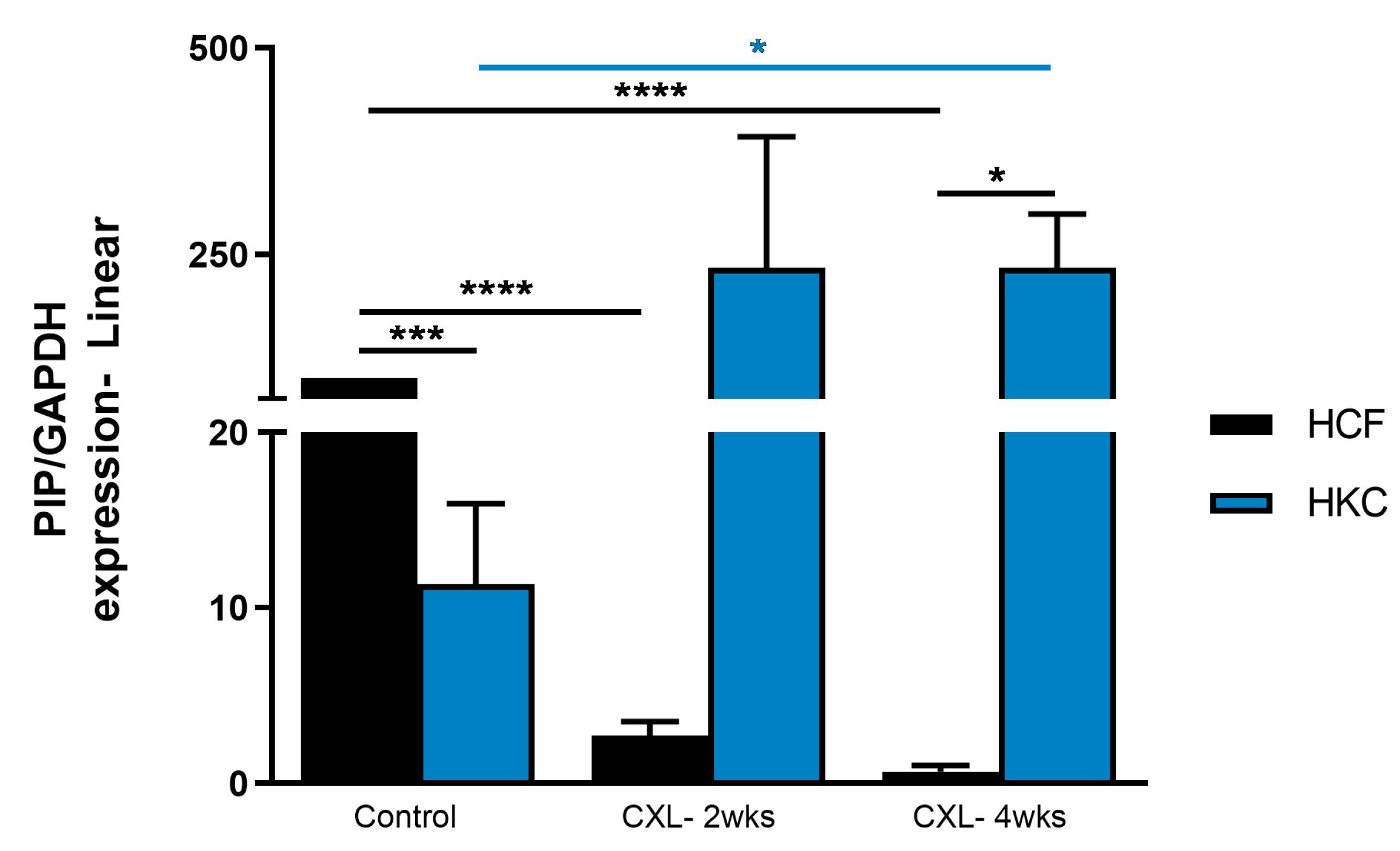
3.3. Prolactin-Induced Protein
3.4. Intermediate Targets
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sherwin, T.; Brookes, N.H. Morphological changes in keratoconus: Pathology or pathogenesis. Clin. Exp. Ophthalmol. 2004, 32, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Fournie, P.; Touboul, D.; Arne, J.L.; Colin, J.; Malecaze, F. Keratoconus. J. Fr. Ophtalmol. 2013, 36, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Chen, T.; Moktar, A.; Chan, E.; Chong, E.W.; Daniell, M.; Sahebjada, S. Diagnosis and Management of Keratoconus-A Narrative Review of Clinicians’ Perspectives. Children 2022, 9, 1973. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, R.M.; van den Berg, A.B.; Dodhia, M.; Shahid, M.; Jammal, A.A.; de Freitas, D.; Rocha, K.M. Scheimpflug-Derived Corneal Lower and Higher Order Aberrations Post Intrastromal Corneal Ring Segments for Keratoconus. Vision 2022, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.S.; Dockery, P.W.; Jacob, S.; Parker, J.S. Preimplantation dehydration for corneal allogenic intrastromal ring segment implantation. J. Cataract. Refract. Surg. 2021, 47, e37–e39. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, T.; Tuli, S.S.; Steigleman, W.A.; Shah, A.A. Overview of Corneal Transplantation for the Nonophthalmologist. Transplant. Direct 2023, 9, e1434. [Google Scholar] [CrossRef]
- Angelo, L.; Gokul Boptom, A.; McGhee, C.; Ziaei, M. Corneal Crosslinking: Present and Future. Asia Pac. J. Ophthalmol. 2022, 11, 441–452. [Google Scholar] [CrossRef]
- Wollensak, G.; Spoerl, E.; Seiler, T. Riboflavin/ultraviolet-a-induced collagen crosslinking for the treatment of keratoconus. Am. J. Ophthalmol. 2003, 135, 620–627. [Google Scholar] [CrossRef]
- Vohra, V.; Tuteja, S.; Gurnani, B.; Chawla, H. Collagen Cross Linking For Keratoconus. In StatPearls; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Srivatsa, S.; Jacob, S.; Agarwal, A. Contact lens assisted corneal cross linking in thin ectatic corneas—A review. Indian J. Ophthalmol. 2020, 68, 2773–2778. [Google Scholar] [CrossRef]
- Agarwal, R.; Jain, P.; Arora, R. Complications of corneal collagen cross-linking. Indian J. Ophthalmol. 2022, 70, 1466–1474. [Google Scholar] [CrossRef]
- O’Brart, D.P.; Patel, P.; Lascaratos, G.; Wagh, V.K.; Tam, C.; Lee, J.; O’Brart, N.A. Corneal Cross-linking to Halt the Progression of Keratoconus and Corneal Ectasia: Seven-Year Follow-up. Am. J. Ophthalmol. 2015, 160, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, N.; Varssano, D. Corneal collagen crosslinking: A systematic review. Ophthalmologica 2014, 232, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Petroll, W.M.; Varner, V.D.; Schmidtke, D.W. Keratocyte mechanobiology. Exp. Eye Res. 2020, 200, 108228. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Fowler, B.; Karamichos, D. Collagen cross-linking impact on keratoconus extracellular matrix. PLoS ONE 2018, 13, e0200704. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Sejersen, H.; Frank, G.; Hjortdal, J.; Karamichos, D. Effects of collagen cross-linking on the keratoconus metabolic network. Eye 2018, 32, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Hjortdal, J.; Sejersen, H.; Frank, G.; Karamichos, D. Human in vitro Model Reveals the Effects of Collagen Cross-linking on Keratoconus Pathogenesis. Sci. Rep. 2017, 7, 12517. [Google Scholar] [CrossRef]
- Gao, X.; Nannini, D.R.; Corrao, K.; Torres, M.; Chen, Y.I.; Fan, B.J.; Wiggs, J.L.; International Glaucoma Genetics Consortium; Taylor, K.D.; Gauderman, W.J.; et al. Genome-wide association study identifies WNT7B as a novel locus for central corneal thickness in Latinos. Hum. Mol. Genet. 2016, 25, 5035–5045. [Google Scholar] [CrossRef]
- Cuellar-Partida, G.; Springelkamp, H.; Lucas, S.E.; Yazar, S.; Hewitt, A.W.; Iglesias, A.I.; Montgomery, G.W.; Martin, N.G.; Pennell, C.E.; van Leeuwen, E.M.; et al. WNT10A exonic variant increases the risk of keratoconus by decreasing corneal thickness. Hum. Mol. Genet. 2015, 24, 5060–5068. [Google Scholar] [CrossRef]
- Khaled, M.L.; Bykhovskaya, Y.; Yablonski, S.E.R.; Li, H.; Drewry, M.D.; Aboobakar, I.F.; Estes, A.; Gao, X.R.; Stamer, W.D.; Xu, H.; et al. Differential Expression of Coding and Long Noncoding RNAs in Keratoconus-Affected Corneas. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2717–2728. [Google Scholar] [CrossRef]
- Loukovitis, E.; Sfakianakis, K.; Syrmakesi, P.; Tsotridou, E.; Orfanidou, M.; Bakaloudi, D.R.; Stoila, M.; Kozei, A.; Koronis, S.; Zachariadis, Z.; et al. Genetic Aspects of Keratoconus: A Literature Review Exploring Potential Genetic Contributions and Possible Genetic Relationships with Comorbidities. Ophthalmol. Ther. 2018, 7, 263–292. [Google Scholar] [CrossRef]
- Nacaroglu, S.A.; Kirgiz, A.; Kandemir Besek, N.; Taskapili, M. Predictive Factors for Corneal Scarring in Progressive Keratoconus Patients after Corneal Collagen Cross-linking. Ophthalmic Epidemiol. 2021, 28, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Stachon, T.; Stachon, A.; Hartmann, U.; Seitz, B.; Langenbucher, A.; Szentmary, N. Urea, Uric Acid, Prolactin and fT4 Concentrations in Aqueous Humor of Keratoconus Patients. Curr. Eye Res. 2017, 42, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, S.; Hjortdal, J.; Sarker-Nag, A.; Sejersen, H.; Asara, J.M.; Karamichos, D. Gross cystic disease fluid protein-15/prolactin-inducible protein as a biomarker for keratoconus disease. PLoS ONE 2014, 9, e113310. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, L.; Rossi, F.; Ruocco, C.; Ragni, M.; Carruba, M.O.; Valerio, A.; Nisoli, E. An original amino acid formula favours in vitro corneal epithelial wound healing by promoting Fn1, ITGB1, and PGC-1alpha expression. Exp. Eye Res. 2022, 219, 109060. [Google Scholar] [CrossRef] [PubMed]
- Yeung, V.; Sriram, S.; Tran, J.A.; Guo, X.; Hutcheon, A.E.K.; Zieske, J.D.; Karamichos, D.; Ciolino, J.B. FAK Inhibition Attenuates Corneal Fibroblast Differentiation In Vitro. Biomolecules 2021, 11, 1682. [Google Scholar] [CrossRef] [PubMed]
- Amit, C.; Padmanabhan, P.; Narayanan, J. Deciphering the mechanoresponsive role of beta-catenin in keratoconus epithelium. Sci. Rep. 2020, 10, 21382. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Hutcheon, A.E.; Melotti, S.A.; Zieske, J.D.; Trinkaus-Randall, V.; Ruberti, J.W. Morphologic characterization of organized extracellular matrix deposition by ascorbic acid-stimulated human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4050–4060. [Google Scholar] [CrossRef]
- Priyadarsini, S.; McKay, T.B.; Sarker-Nag, A.; Karamichos, D. Keratoconus in vitro and the key players of the TGF-beta pathway. Mol. Vis. 2015, 21, 577–588. [Google Scholar]
- Karamichos, D.; Guo, X.Q.; Hutcheon, A.E.; Zieske, J.D. Human corneal fibrosis: An in vitro model. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1382–1388. [Google Scholar] [CrossRef]
- Escandon, P.; Nicholas, S.E.; Cunningham, R.L.; Murphy, D.A.; Riaz, K.M.; Karamichos, D. The Role of Estriol and Estrone in Keratoconic Stromal Sex Hormone Receptors. Int. J. Mol. Sci. 2022, 23, 916. [Google Scholar] [CrossRef]
- Kabza, M.; Karolak, J.A.; Rydzanicz, M.; Szczesniak, M.W.; Nowak, D.M.; Ginter-Matuszewska, B.; Polakowski, P.; Ploski, R.; Szaflik, J.P.; Gajecka, M. Collagen synthesis disruption and downregulation of core elements of TGF-beta, Hippo, and Wnt pathways in keratoconus corneas. Eur. J. Hum. Genet. 2017, 25, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W.; Parikh, R.N.; Wang, J.; Bower, K.S.; Matthaei, M.; Chakravarti, S.; Jun, A.S.; Eberhart, C.G.; Soiberman, U.S. Transcriptomic and Immunohistochemical Analysis of Progressive Keratoconus Reveal Altered WNT10A in Epithelium and Bowman’s Layer. Investig. Ophthalmol. Vis. Sci. 2021, 62, 16. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Bak-Nielsen, S.; Sejersen, H.; Ding, K.; Hjortdal, J.; Karamichos, D. Prolactin-Induced Protein is a novel biomarker for Keratoconus. Exp. Eye Res. 2019, 179, 55–63. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Corley, S.M.; Wen, L.; Hodge, C.; Hollhumer, R.; Madigan, M.C.; Wilkins, M.R.; Sutton, G. RNA-Seq analysis and comparison of corneal epithelium in keratoconus and myopia patients. Sci. Rep. 2018, 8, 389. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Estes, A.; Liu, Y. Omics analyses in keratoconus: From transcriptomics to proteomics. Curr. Ophthalmol. Rep. 2020, 8, 216–225. [Google Scholar] [CrossRef]
- Nakatsu, M.N.; Ding, Z.; Ng, M.Y.; Truong, T.T.; Yu, F.; Deng, S.X. Wnt/beta-catenin signaling regulates proliferation of human cornea epithelial stem/progenitor cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4734–4741. [Google Scholar] [CrossRef]
- Kulkarni, M.; Leszczynska, A.; Wei, G.; Winkler, M.A.; Tang, J.; Funari, V.A.; Deng, N.; Liu, Z.; Punj, V.; Deng, S.X.; et al. Genome-wide analysis suggests a differential microRNA signature associated with normal and diabetic human corneal limbus. Sci. Rep. 2017, 7, 3448. [Google Scholar] [CrossRef]
- Liang, W.; Huang, L.; Ma, X.; Dong, L.; Cheng, R.; Dehdarani, M.; Karamichos, D.; Ma, J.X. Pathogenic Role of Diabetes-Induced Overexpression of Kallistatin in Corneal Wound Healing Deficiency Through Inhibition of Canonical Wnt Signaling. Diabetes 2022, 71, 747–761. [Google Scholar] [CrossRef]
- Sharif, R.; Bak-Nielsen, S.; Hjortdal, J.; Karamichos, D. Pathogenesis of Keratoconus: The intriguing therapeutic potential of Prolactin-inducible protein. Prog. Retin. Eye Res. 2018, 67, 150–167. [Google Scholar] [CrossRef]
- Liang, W.; Ma, J.X.; Van, L.; Vasini, B.; Karamichos, D. Prolactin-Induced Protein facilitates corneal wound healing. Exp. Eye Res. 2022, 225, 109300. [Google Scholar] [CrossRef]
- Haier, J.; Gallick, G.E.; Nicolson, G.L. Src protein kinase pp60c-src influences adhesion stabilization of HT-29 colon carcinoma cells to extracellular matrix components under dynamic conditions of laminar flow. J. Exp. Ther. Oncol. 2002, 2, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Tannour-Louet, M.; York, B.; Tang, K.; Stashi, E.; Bouguerra, H.; Zhou, S.; Yu, H.; Wong, L.J.; Stevens, R.D.; Xu, J.; et al. Hepatic SRC-1 activity orchestrates transcriptional circuitries of amino acid pathways with potential relevance for human metabolic pathogenesis. Mol. Endocrinol. 2014, 28, 1707–1718. [Google Scholar] [CrossRef]
- Puigserver, P.; Spiegelman, B.M. Peroxisome proliferator-activated receptor-gamma coactivator 1 alpha (PGC-1 alpha): Transcriptional coactivator and metabolic regulator. Endocr. Rev. 2003, 24, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.D.; Yang, D.I.; Lin, T.K.; Shaw, F.Z.; Liou, C.W.; Chuang, Y.C. Roles of oxidative stress, apoptosis, PGC-1alpha and mitochondrial biogenesis in cerebral ischemia. Int. J. Mol. Sci. 2011, 12, 7199–7215. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karamichos, D.; Nicholas, S.E.; Khan, A.; Riaz, K.M. Collagen Crosslinking for Keratoconus: Cellular Signaling Mechanisms. Biomolecules 2023, 13, 696. https://doi.org/10.3390/biom13040696
Karamichos D, Nicholas SE, Khan A, Riaz KM. Collagen Crosslinking for Keratoconus: Cellular Signaling Mechanisms. Biomolecules. 2023; 13(4):696. https://doi.org/10.3390/biom13040696
Chicago/Turabian StyleKaramichos, Dimitrios, Sarah E. Nicholas, Asher Khan, and Kamran M. Riaz. 2023. "Collagen Crosslinking for Keratoconus: Cellular Signaling Mechanisms" Biomolecules 13, no. 4: 696. https://doi.org/10.3390/biom13040696