
Pivotal Role of Ubiquitin Carboxyl-Terminal Hydrolase L1 (UCHL1) in Uterine Leiomyoma
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Study Approval
2.3. Immunohistochemistry
2.4. RNA Isolation and Quantitative Real-Time PCR Analysis
2.5. Protein Isolation and Western Blotting Analysis
2.6. Cell Culture
2.7. UCHL1 shRNA Transduction and Cell Treatment
2.8. UCHL1 Inhibition and Cell Treatment
2.9. Collagen Gel Contraction Assay
2.10. Wound-Healing Assay
2.11. Statistical Methods
3. Results
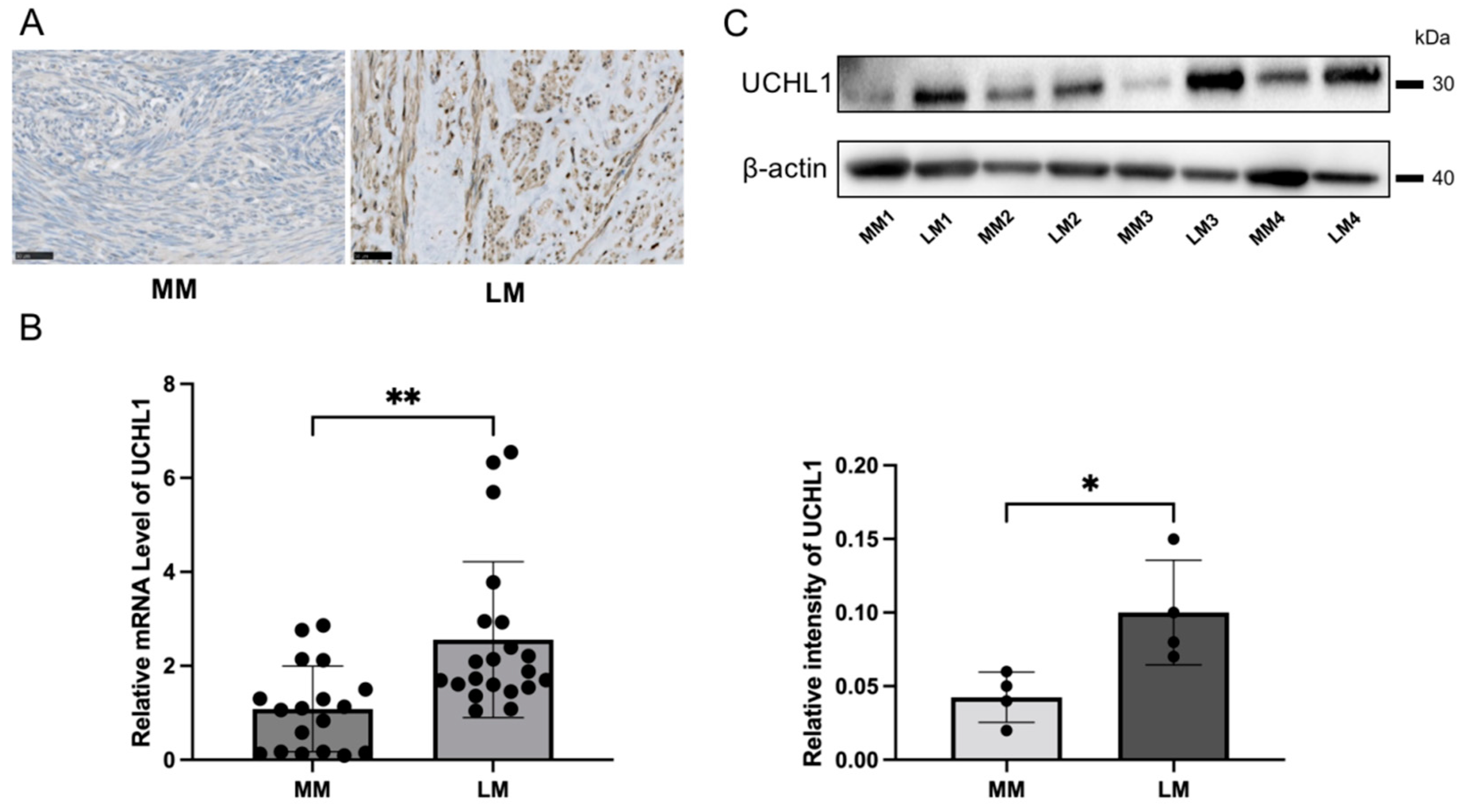
3.1. UCHL1 Expression in Human Uterine Leiomyoma Significantly Increased Compared to Expression in the Myometrium
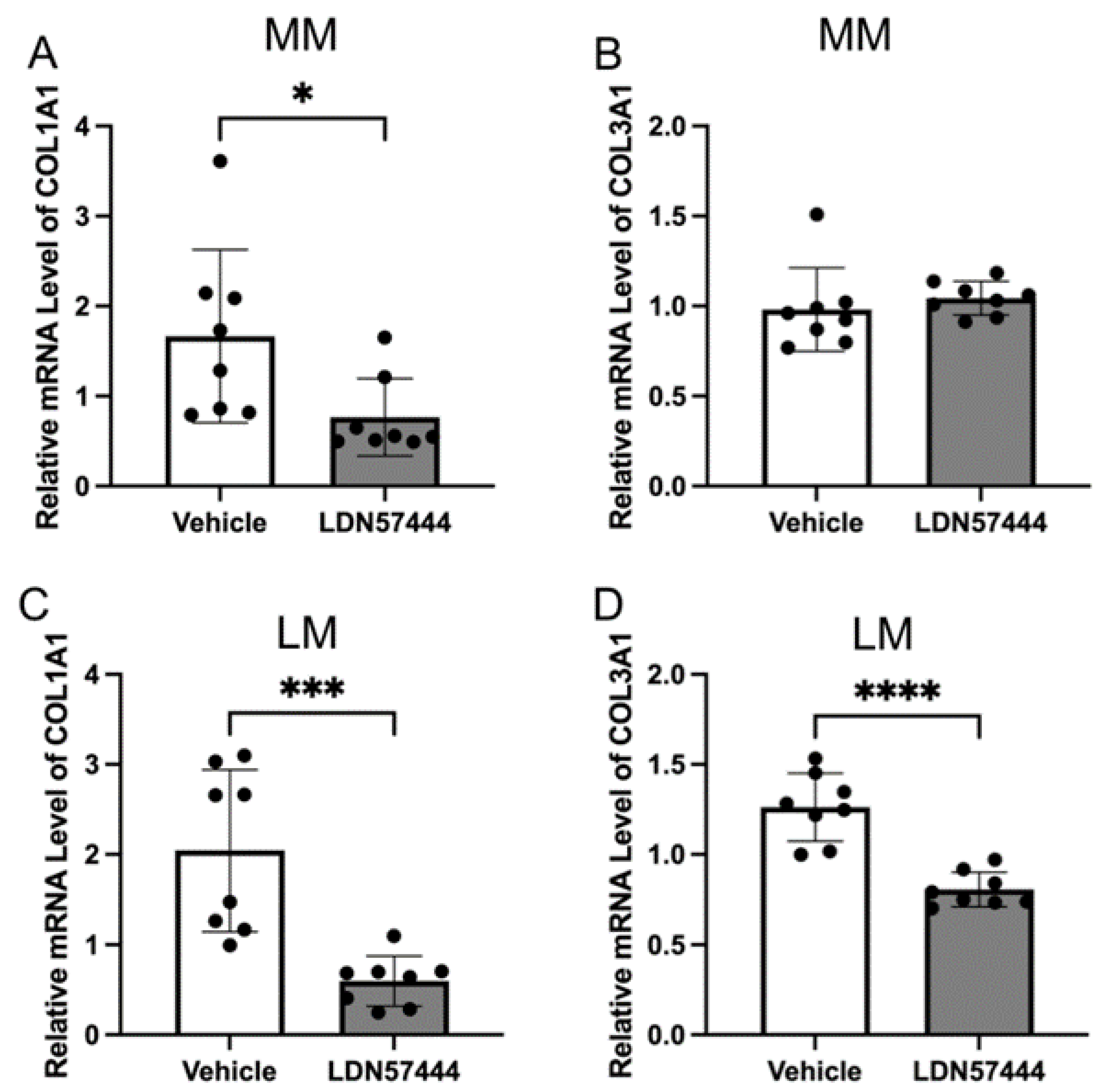
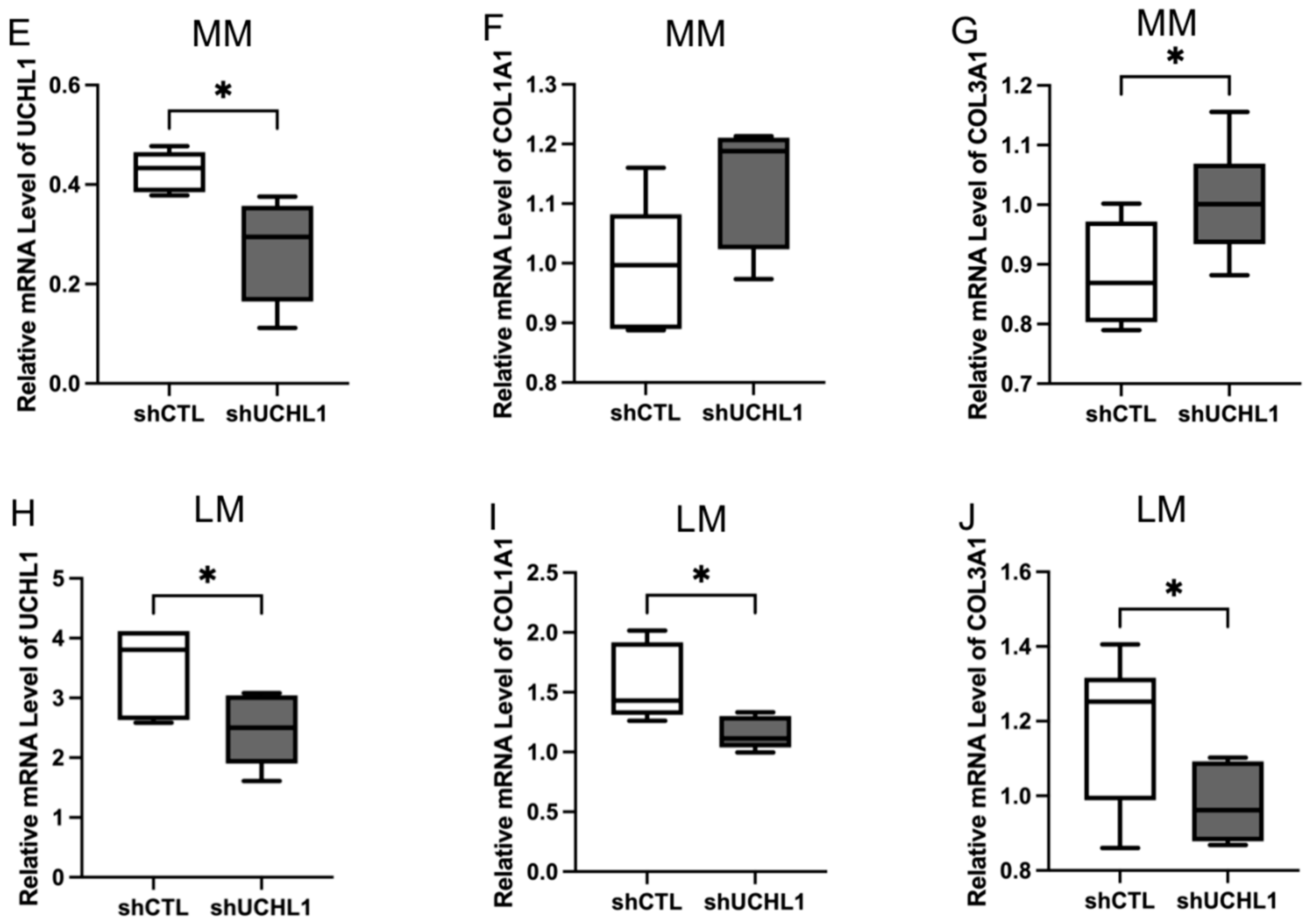
3.2. UCHL1 Silencing and Inhibition Downregulated the Collagen Production in Leiomyoma
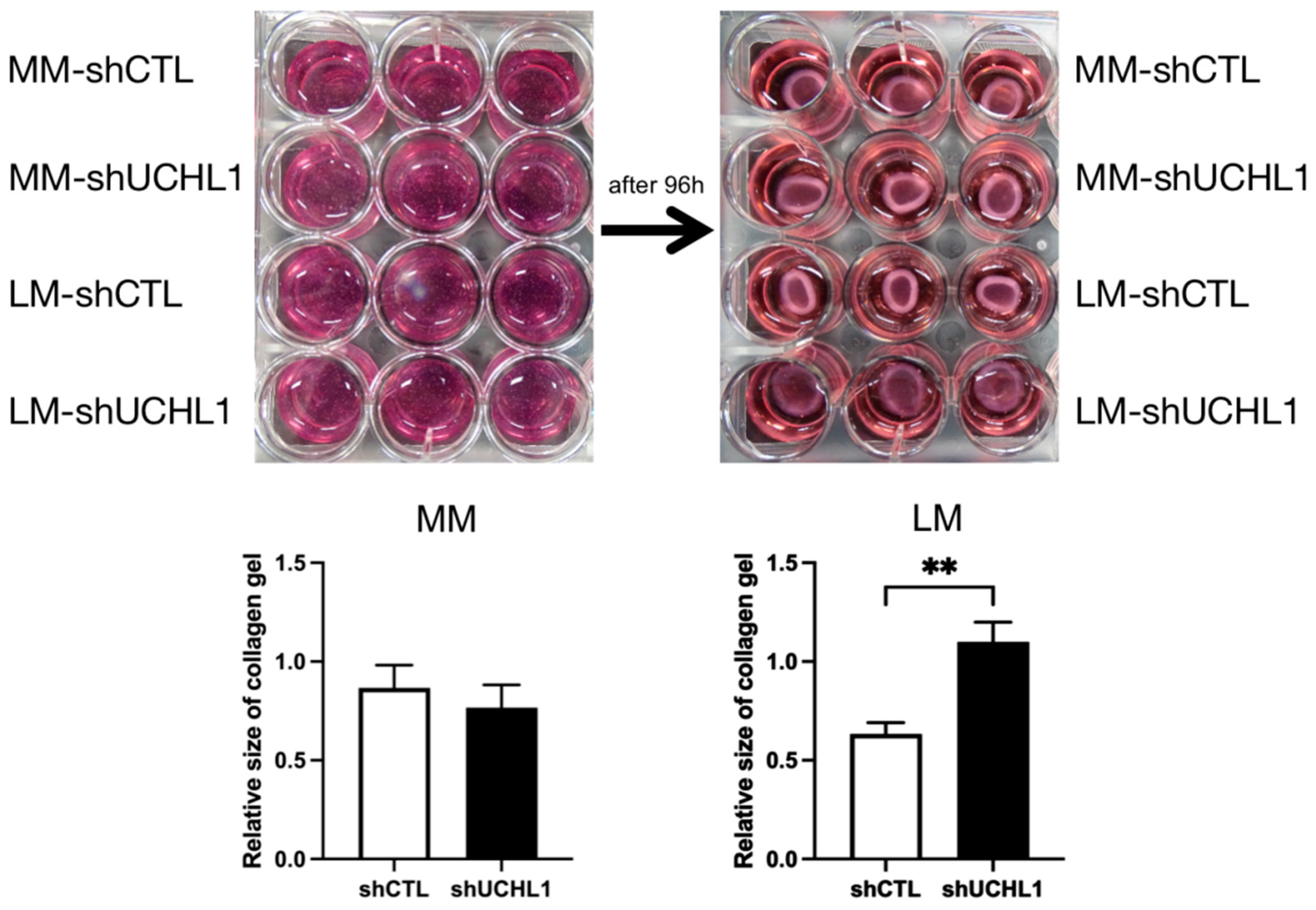
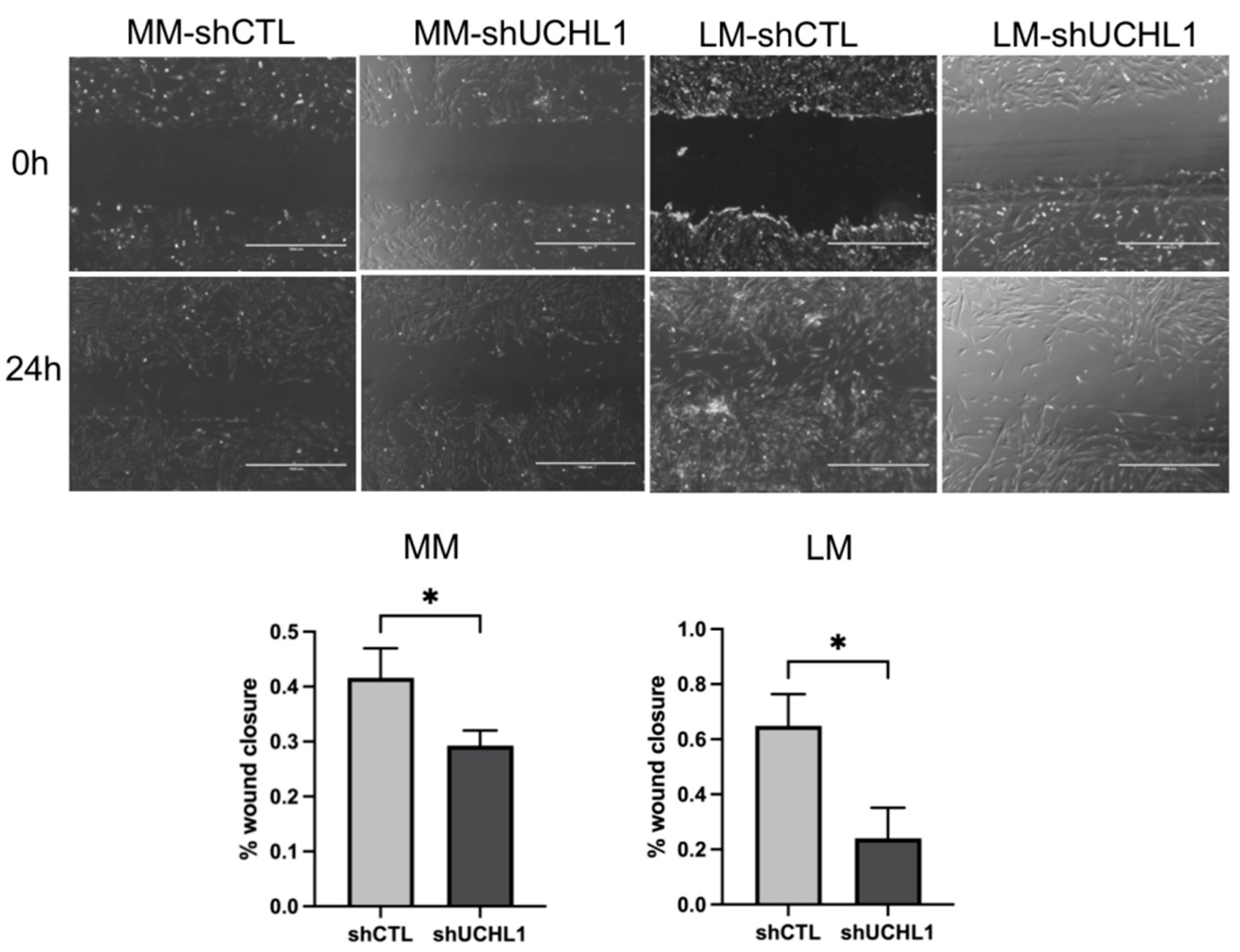
3.3. Effects of UCHL1 Knockdown on Collagen Gel Contraction and Wound-Healing Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stewart, E.A.; Cookson, C.L.; Gandolfo, R.A.; Schulze-Rath, R. Epidemiology of uterine fibroids: A systematic review. BJOG 2017, 124, 1501–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparic, R.; Mirkovic, L.; Malvasi, A.; Tinelli, A. Epidemiology of Uterine Myomas: A Review. Int. J. Fertil. Steril. 2016, 9, 424–435. [Google Scholar] [PubMed]
- Evans, P.; Brunsell, S. Uterine fibroid tumors: Diagnosis and treatment. Am. Fam. Physician. 2007, 75, 1503–1508. [Google Scholar]
- Wise, L.A.; Laughlin-Tommaso, S.K. Epidemiology of Uterine Fibroids: From Menarche to Menopause. Clin. Obstet. Gynecol. 2016, 59, 2–24. [Google Scholar] [CrossRef] [Green Version]
- Sheiner, E.; Bashiri, A.; Levy, A.; Hershkovitz, R.; Katz, M.; Mazor, M. Obstetric characteristics and perinatal outcome of pregnancies with uterine leiomyomas. J. Reprod. Med. 2004, 49, 182–186. [Google Scholar] [CrossRef]
- Mauskopf, J.; Flynn, M.; Thieda, P.; Spalding, J.; Duchane, J. The economic impact of uterine fibroids in the United States: A summary of published estimates. J. Womens Health 2005, 14, 692–703. [Google Scholar] [CrossRef]
- Marsh, E.E.; Steinberg, M.L.; Parker, J.B.; Wu, J.; Chakravarti, D.; Bulun, S.E. Decreased expression of microRNA-29 family in leiomyoma contributes to increased major fibrillar collagen production. Fertil. Steril. 2016, 106, 766–772. [Google Scholar] [CrossRef] [Green Version]
- Willis, M.S.; Patterson, C. Into the heart: The emerging role of the ubiquitin-proteasome system. J. Mol. Cell. Cardiol. 2006, 41, 567–579. [Google Scholar] [CrossRef]
- Bishop, P.; Rocca, D.; Henley, J.M. Ubiquitin C-terminal hydrolase L1 (UCH-L1): Structure, distribution and roles in brain function and dysfunction. Biochem. J. 2016, 473, 2453–2462. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Zhang, Y.L.; Fu, T.T.; Li, P.B.; Cong, T.; Li, H.H. Blockage of UCHL1 activity attenuates cardiac remodeling in spontaneously hypertensive rats. Hypertens. Res. 2020, 43, 1089–1098. [Google Scholar] [CrossRef]
- Gong, Z.; Ye, Q.; Wu, J.W.; Zhou, J.L.; Kong, X.Y.; Ma, L.K. UCHL1 inhibition attenuates cardiac fibrosis via modulation of nuclear factor-kappaB signaling in fibroblasts. Eur. J. Pharmacol. 2021, 900, 174045. [Google Scholar] [CrossRef] [PubMed]
- Woodman, M.F.; Ozcan, M.C.H.; Gura, M.A.; De La Cruz, P.; Gadson, A.K.; Grive, K.J. The requirement of ubiquitin C-terminal hydrolase L1 in mouse ovarian development and fertilitydagger. Biol. Reprod. 2022, 107, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.Q.; Chen, Q.J.; Gu, Z.; Shi, Y.; Yao, Y.W.; Wang, J.; Sun, Z.G.; Tso, J.K. Ubiquitin carboxyl-terminal hydrolase L1 contributes to the oocyte selective elimination in prepubertal mouse ovaries. Sheng Li Xue Bao 2009, 61, 175–184. [Google Scholar] [PubMed]
- Schnell, J.D.; Hicke, L. Non-traditional functions of ubiquitin and ubiquitin-binding proteins. J. Biol. Chem. 2003, 278, 35857–35860. [Google Scholar] [CrossRef] [Green Version]
- Zubiete-Franco, I.; Fernandez-Tussy, P.; Barbier-Torres, L.; Simon, J.; Fernandez-Ramos, D.; Lopitz-Otsoa, F.; Gutierrez-de Juan, V.; de Davalillo, S.L.; Duce, A.M.; Iruzubieta, P.; et al. Deregulated neddylation in liver fibrosis. Hepatology 2017, 65, 694–709. [Google Scholar] [CrossRef] [Green Version]
- Mehine, M.; Ahvenainen, T.; Khamaiseh, S.; Harkonen, J.; Reinikka, S.; Heikkinen, T.; Ayravainen, A.; Pakarinen, P.; Harkki, P.; Pasanen, A.; et al. A novel uterine leiomyoma subtype exhibits NRF2 activation and mutations in genes associated with neddylation of the Cullin 3-RING E3 ligase. Oncogenesis 2022, 11, 52. [Google Scholar] [CrossRef]
- Makinen, N.; Kampjarvi, K.; Frizzell, N.; Butzow, R.; Vahteristo, P. Characterization of MED12, HMGA2, and FH alterations reveals molecular variability in uterine smooth muscle tumors. Mol. Cancer 2017, 16, 101. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Qiang, W.; Serna, V.A.; Yin, P.; Coon, J.S.T.; Navarro, A.; Monsivais, D.; Kakinuma, T.; Dyson, M.; Druschitz, S.; et al. Role of stem cells in human uterine leiomyoma growth. PLoS ONE 2012, 7, e36935. [Google Scholar] [CrossRef]
- Ono, M.; Yin, P.; Navarro, A.; Moravek, M.B.; Coon, J.S.T.; Druschitz, S.A.; Serna, V.A.; Qiang, W.; Brooks, D.C.; Malpani, S.S.; et al. Paracrine activation of WNT/beta-catenin pathway in uterine leiomyoma stem cells promotes tumor growth. Proc. Natl. Acad. Sci. USA 2013, 110, 17053–17058. [Google Scholar] [CrossRef] [Green Version]
- Bulun, S.E. Uterine fibroids. N. Engl. J. Med. 2013, 369, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Chuang, T.D.; Khorram, O. Tranilast Inhibits Genes Functionally Involved in Cell Proliferation, Fibrosis, and Epigenetic Regulation and Epigenetically Induces miR-29c Expression in Leiomyoma Cells. Reprod. Sci. 2017, 24, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Papa, L.; Brophy, G.M.; Welch, R.D.; Lewis, L.M.; Braga, C.F.; Tan, C.N.; Ameli, N.J.; Lopez, M.A.; Haeussler, C.A.; Mendez Giordano, D.I.; et al. Time Course and Diagnostic Accuracy of Glial and Neuronal Blood Biomarkers GFAP and UCH-L1 in a Large Cohort of Trauma Patients with and Without Mild Traumatic Brain Injury. JAMA Neurol. 2016, 73, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, Y.; Zeng, L.; Yeom, C.J.; Zhu, Y.; Morinibu, A.; Shinomiya, K.; Kobayashi, M.; Hirota, K.; Itasaka, S.; Yoshimura, M.; et al. UCHL1 provides diagnostic and antimetastatic strategies due to its deubiquitinating effect on HIF-1alpha. Nat. Commun. 2015, 6, 6153. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, E.; Aga, M.; Kondo, S.; Whitehurst, C.; Yoshizaki, T.; Pagano, J.S.; Shackelford, J. C-Terminal Farnesylation of UCH-L1 Plays a Role in Transport of Epstein-Barr Virus Primary Oncoprotein LMP1 to Exosomes. mSphere 2018, 3, e00030-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattuada, D.; Vigano, P.; Mangioni, S.; Sassone, J.; Di Francesco, S.; Vignali, M.; Di Blasio, A.M. Accumulation of retinoid X receptor-alpha in uterine leiomyomas is associated with a delayed ligand-dependent proteasome-mediated degradation and an alteration of its transcriptional activity. Mol. Endocrinol. 2007, 21, 602–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Kim, Y.M.; Lim, S.; Nam, Y.K.; Jeong, J.; Kim, H.J.; Lee, K.J. Ubiquitin C-terminal hydrolase-L1 is a key regulator of tumor cell invasion and metastasis. Oncogene 2009, 28, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartnett, S.; Zhang, F.; Abitz, A.; Li, Y. Ubiquitin C-terminal hydrolase L1 interacts with choline transporter in cholinergic cells. Neurosci. Lett. 2014, 564, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Levey, A.I.; Weintraub, S.T.; Rees, H.D.; Gearing, M.; Chin, L.S.; Li, L. Oxidative modifications and down-regulation of ubiquitin carboxyl-terminal hydrolase L1 associated with idiopathic Parkinson’s and Alzheimer’s diseases. J. Biol. Chem. 2004, 279, 13256–13264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drobysheva, A.; Ahmad, M.; White, R.; Wang, H.W.; Leenen, F.H. Cardiac sympathetic innervation and PGP9.5 expression by cardiomyocytes after myocardial infarction: Effects of central MR blockade. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1817–H1829. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, V.; Zhao, M.; Reddy, S.; Fajardo, G.; Wang, X.; Dewey, S.; Gomes, A.V.; Bernstein, D. Altered ubiquitin-proteasome signaling in right ventricular hypertrophy and failure. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H551–H562. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.; Hockel, M.; Wree, A.; Leo, C.; Horn, L.C.; Vaupel, P. Lack of hypoxic response in uterine leiomyomas despite severe tissue hypoxia. Cancer Res. 2008, 68, 4719–4726. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Xu, L.; Sone, K.; Kobayashi, T.; Wang, G.; Shozu, M. Hypoxia Induces Hypoxia-Inducible Factor 1alpha and Potential HIF-Responsive Gene Expression in Uterine Leiomyoma. Reprod. Sci. 2019, 26, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Maruyama, T.; Masuda, H.; Kajitani, T.; Nagashima, T.; Arase, T.; Ito, M.; Ohta, K.; Uchida, H.; Asada, H.; et al. Side population in human uterine myometrium displays phenotypic and functional characteristics of myometrial stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 18700–18705. [Google Scholar] [CrossRef] [Green Version]
- Miyashita-Ishiwata, M.; El Sabeh, M.; Reschke, L.D.; Afrin, S.; Borahay, M.A. Differential response to hypoxia in leiomyoma and myometrial cells. Life Sci. 2022, 290, 120238. [Google Scholar] [CrossRef]
- Zhou, S.; Yi, T.; Shen, K.; Zhang, B.; Huang, F.; Zhao, X. Hypoxia: The driving force of uterine myometrial stem cells differentiation into leiomyoma cells. Med. Hypotheses 2011, 77, 985–986. [Google Scholar] [CrossRef]
- Osaka, H.; Wang, Y.L.; Takada, K.; Takizawa, S.; Setsuie, R.; Li, H.; Sato, Y.; Nishikawa, K.; Sun, Y.J.; Sakurai, M.; et al. Ubiquitin carboxy-terminal hydrolase L1 binds to and stabilizes monoubiquitin in neuron. Hum. Mol. Genet. 2003, 12, 1945–1958. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Freeling, J.; Wu, P.; Liang, A.P.; Wang, X.; Li, Y. UCHL1 regulates muscle fibers and mTORC1 activity in skeletal muscle. Life Sci. 2019, 233, 116699. [Google Scholar] [CrossRef]
- Gao, H.; Hartnett, S.; Li, Y. Ubiquitin C-Terminal Hydrolase L1 regulates myoblast proliferation and differentiation. Biochem. Biophys. Res. Commun. 2017, 492, 96–102. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, L.; Niu, T.; Shao, L.; Li, H.; Wu, W.; Wang, W.; Lv, L.; Qin, Q.; Wang, F.; et al. Ubiquitin carboxyl terminal hydrolyase L1-suppressed autophagic degradation of p21WAF1/Cip1 as a novel feedback mechanism in the control of cardiac fibroblast proliferation. PLoS ONE 2014, 9, e94658. [Google Scholar] [CrossRef]
- Nakashima, R.; Goto, Y.; Koyasu, S.; Kobayashi, M.; Morinibu, A.; Yoshimura, M.; Hiraoka, M.; Hammond, E.M.; Harada, H. UCHL1-HIF-1 axis-mediated antioxidant property of cancer cells as a therapeutic target for radiosensitization. Sci. Rep. 2017, 7, 6879. [Google Scholar] [CrossRef]
- Zaitseva, M.; Vollenhoven, B.J.; Rogers, P.A. In vitro culture significantly alters gene expression profiles and reduces differences between myometrial and fibroid smooth muscle cells. Mol. Hum. Reprod. 2006, 12, 187–207. [Google Scholar] [CrossRef]
- Wilson, C.L.; Murphy, L.B.; Leslie, J.; Kendrick, S.; French, J.; Fox, C.R.; Sheerin, N.S.; Fisher, A.; Robinson, J.H.; Tiniakos, D.G.; et al. Ubiquitin C-terminal hydrolase 1: A novel functional marker for liver myofibroblasts and a therapeutic target in chronic liver disease. J. Hepatol. 2015, 63, 1421–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, M.; Catherino, W.H. Novel method to characterize primary cultures of leiomyoma and myometrium with the use of confirmatory biomarker gene arrays. Fertil. Steril. 2007, 87, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.A.; Friedman, A.J.; Peck, K.; Nowak, R.A. Relative overexpression of collagen type I and collagen type III messenger ribonucleic acids by uterine leiomyomas during the proliferative phase of the menstrual cycle. J. Clin. Endocrinol. Metab. 1994, 79, 900–906. [Google Scholar]
- Janjusevic, M.; Greco, S.; Islam, M.S.; Castellucci, C.; Ciavattini, A.; Toti, P.; Petraglia, F.; Ciarmela, P. Locostatin, a disrupter of Raf kinase inhibitor protein, inhibits extracellular matrix production, proliferation, and migration in human uterine leiomyoma and myometrial cells. Fertil. Steril. 2016, 106, 1530–1538.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okochi-Takada, E.; Nakazawa, K.; Wakabayashi, M.; Mori, A.; Ichimura, S.; Yasugi, T.; Ushijima, T. Silencing of the UCHL1 gene in human colorectal and ovarian cancers. Int. J. Cancer 2006, 119, 1338–1344. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Yamashita, K.; Osada, M.; Nomoto, S.; Sun, D.I.; Xiao, Y.; Hoque, M.O.; Westra, W.H.; Califano, J.A.; Sidransky, D. Inverse correlation between cyclin A1 hypermethylation and p53 mutation in head and neck cancer identified by reversal of epigenetic silencing. Cancer. Res. 2004, 64, 5982–5987. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Tao, Q.; Cheung, K.F.; Jin, H.; Poon, F.F.; Wang, X.; Li, H.; Cheng, Y.Y.; Rocken, C.; Ebert, M.P.; et al. Epigenetic identification of ubiquitin carboxyl-terminal hydrolase L1 as a functional tumor suppressor and biomarker for hepatocellular carcinoma and other digestive tumors. Hepatology 2008, 48, 508–518. [Google Scholar] [CrossRef]
- Sato, N.; Fukushima, N.; Maitra, A.; Matsubayashi, H.; Yeo, C.J.; Cameron, J.L.; Hruban, R.H.; Goggins, M. Discovery of novel targets for aberrant methylation in pancreatic carcinoma using high-throughput microarrays. Cancer Res. 2003, 63, 3735–3742. [Google Scholar]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug. Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef]
- Yan, C.; Huo, H.; Yang, C.; Zhang, T.; Chu, Y.; Liu, Y. Ubiquitin C-Terminal Hydrolase L1 regulates autophagy by inhibiting autophagosome formation through its deubiquitinating enzyme activity. Biochem. Biophys. Res. Commun. 2018, 497, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Hwang, D.; Bheda-Malge, A.; Whitehurst, C.B.; Kabanov, A.V.; Kondo, S.; Aga, M.; Yoshizaki, T.; Pagano, J.S.; Sokolsky, M.; et al. Inhibition of UCH-L1 Deubiquitinating Activity with Two Forms of LDN-57444 Has Anti-Invasive Effects in Metastatic Carcinoma Cells. Int. J. Mol. Sci. 2019, 20, 3733. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sets |
|---|---|
| UCHL1 | 5′-TGGATGGCCACCTCTATGAAC-3′ 5′-CTTGCTCACGCTCGGTGAAT-3′ |
| COL1A1 | 5′-GGTGAACAGGGTGTTCCTGGAGAC-3′ 5′-AGCATCACCCTTAGCACCATCGTT-3′ |
| COL3A1 | 5′-ATTATTTTGGCACAACAGGAAGCT-3′ 5′-TCCGCATAGGACTGACCAAGAT-3′ |
| ACTB | 5′-CCAACCGCGAGAAGATGA-3′ 5′-CCAGAGGCGTACAGGGATAG-3′ |
| Antibodies | Isotype | Supplier |
|---|---|---|
| UCHL1 | mouse IgG2A | R&D Systems (MAB6007) |
| β-actin (C4) | mouse IgG1κ | Millipore (MAB1501R) |
| HMGA2 | rabbit IgG | Abcam (ab97276) |
| Desmin | mouse IgG | Dako (D33) |
| Vimentin | mouse IgG1 | Merck Millipore (MAB3400) |
| ACTA2 | mouse IgG2a | Dako (M085101) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Dai, Y.; Ono, M.; Kojima, J.; Sasaki, T.; Fujiwara, H.; Kuji, N.; Nishi, H. Pivotal Role of Ubiquitin Carboxyl-Terminal Hydrolase L1 (UCHL1) in Uterine Leiomyoma. Biomolecules 2023, 13, 193. https://doi.org/10.3390/biom13020193
Suzuki T, Dai Y, Ono M, Kojima J, Sasaki T, Fujiwara H, Kuji N, Nishi H. Pivotal Role of Ubiquitin Carboxyl-Terminal Hydrolase L1 (UCHL1) in Uterine Leiomyoma. Biomolecules. 2023; 13(2):193. https://doi.org/10.3390/biom13020193
Chicago/Turabian StyleSuzuki, Tomoo, Yidan Dai, Masanori Ono, Junya Kojima, Toru Sasaki, Hiroshi Fujiwara, Naoaki Kuji, and Hirotaka Nishi. 2023. "Pivotal Role of Ubiquitin Carboxyl-Terminal Hydrolase L1 (UCHL1) in Uterine Leiomyoma" Biomolecules 13, no. 2: 193. https://doi.org/10.3390/biom13020193