
Regulation Mechanisms of Meiotic Recombination Revealed from the Analysis of a Fission Yeast Recombination Hotspot ade6-M26


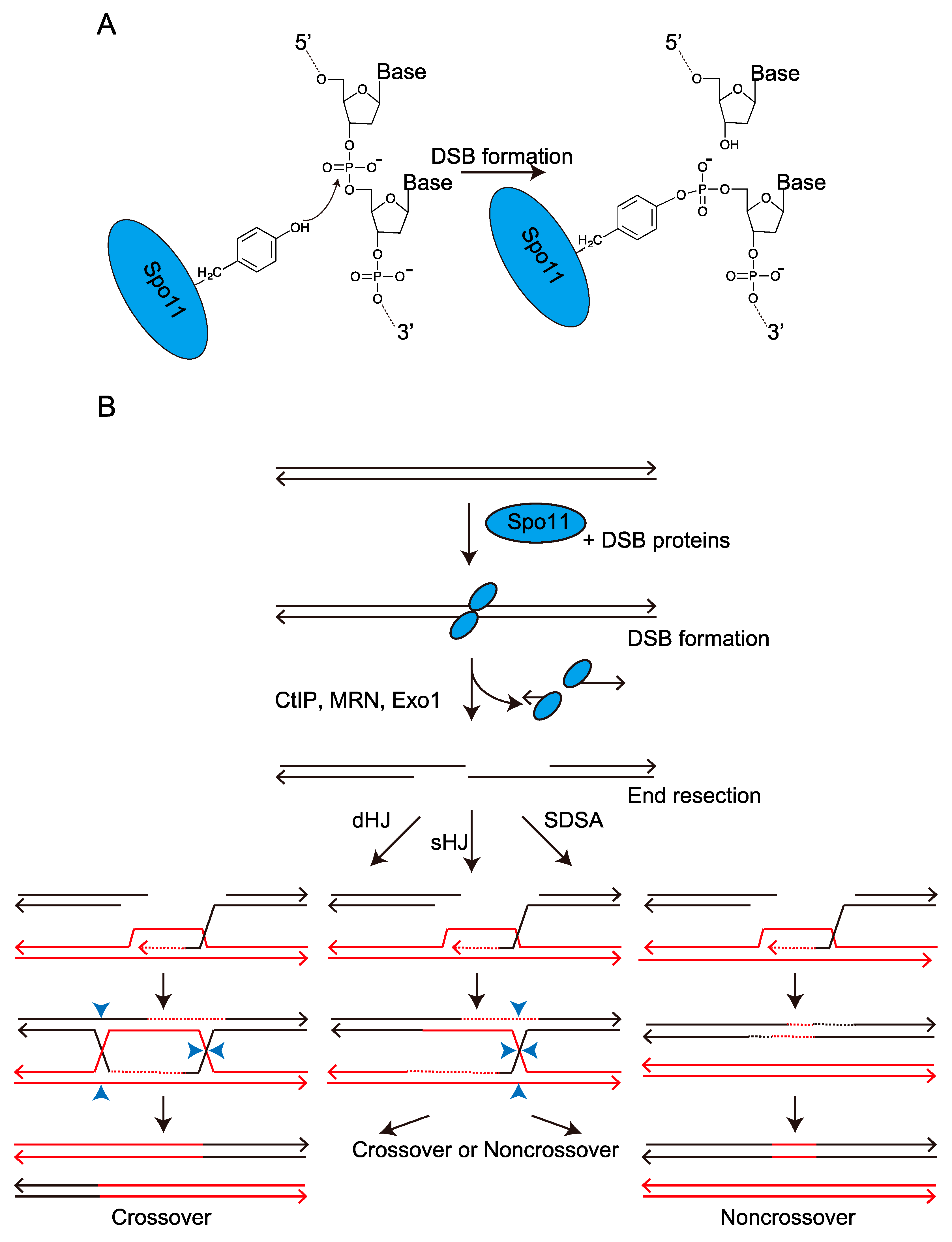{kind=link}
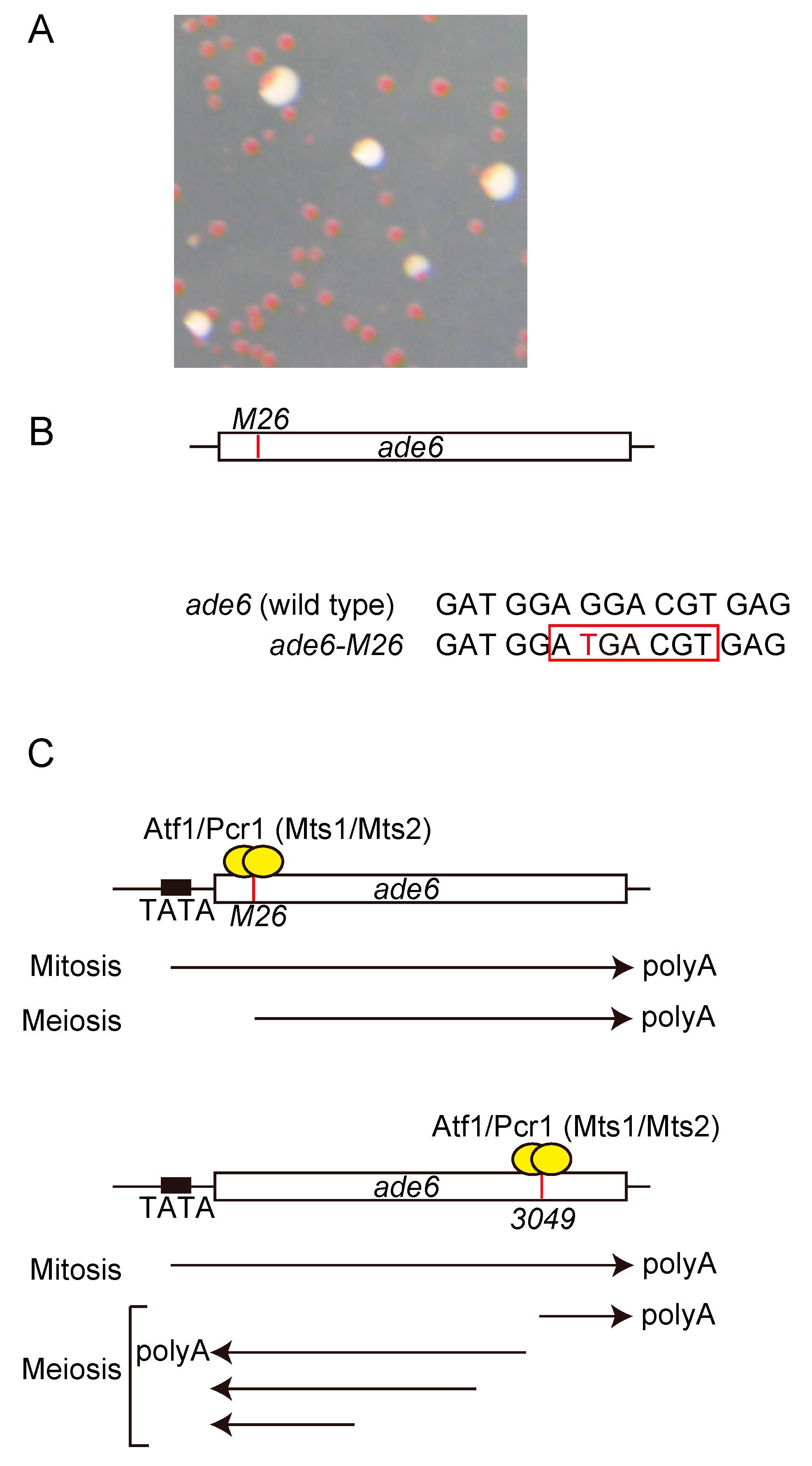{kind=link}
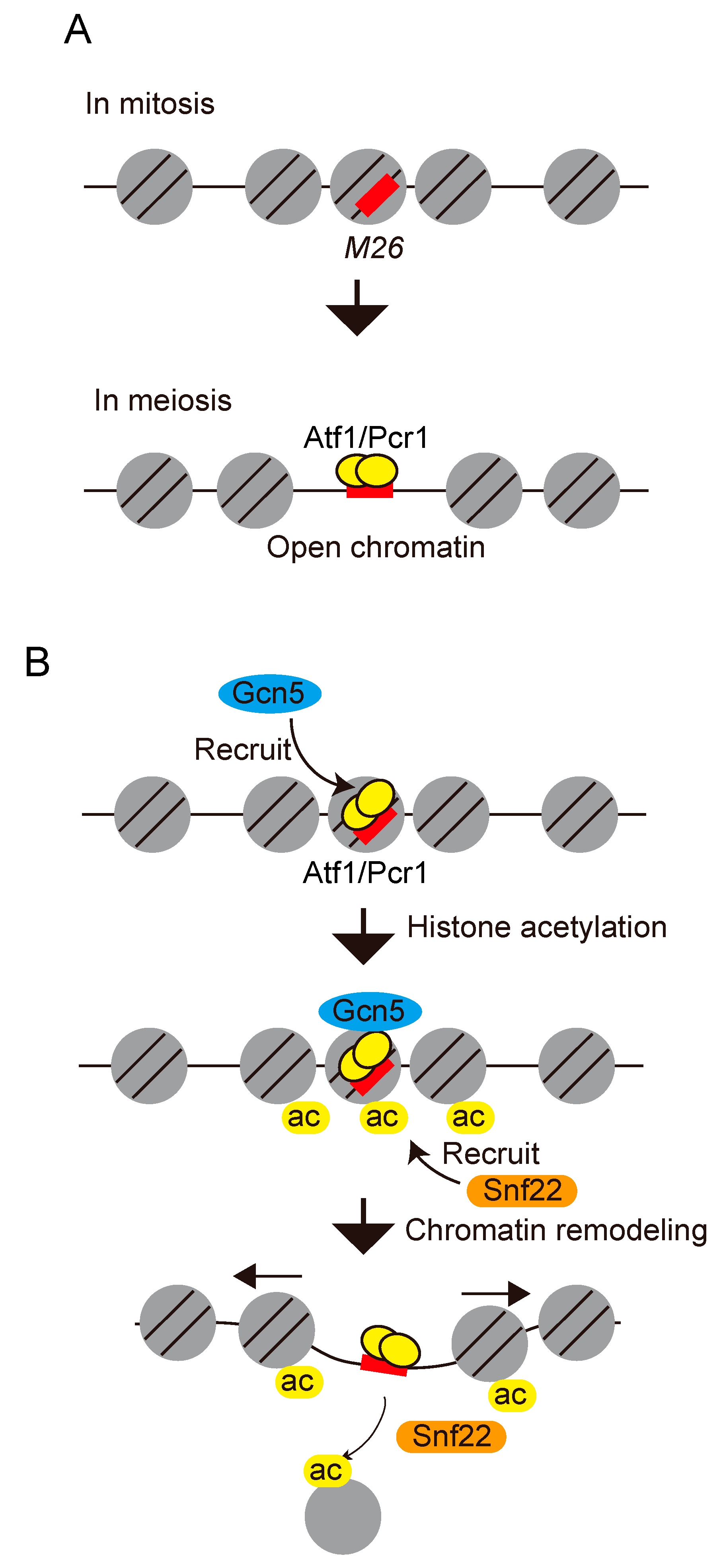{kind=link}
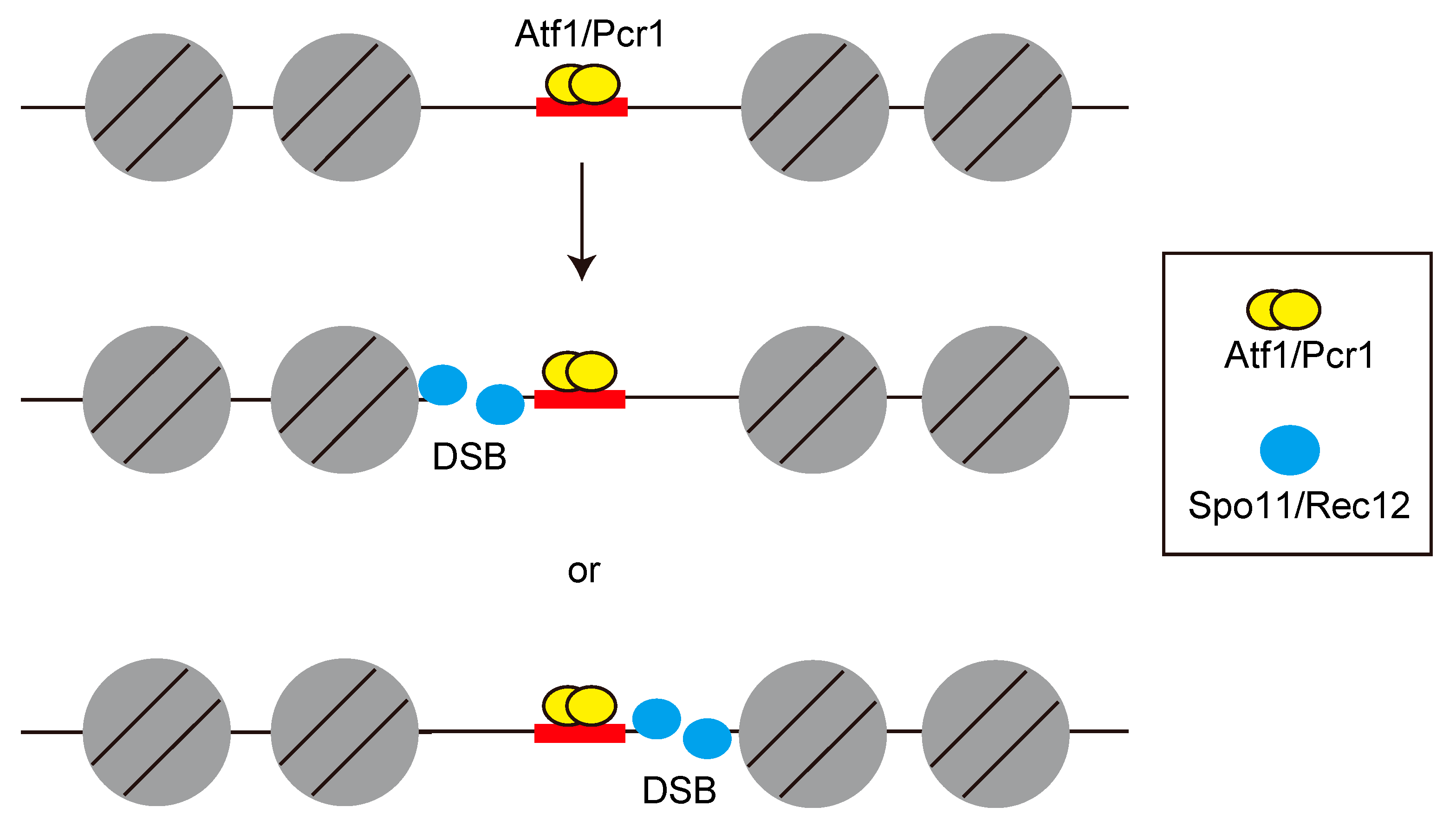{kind=link}
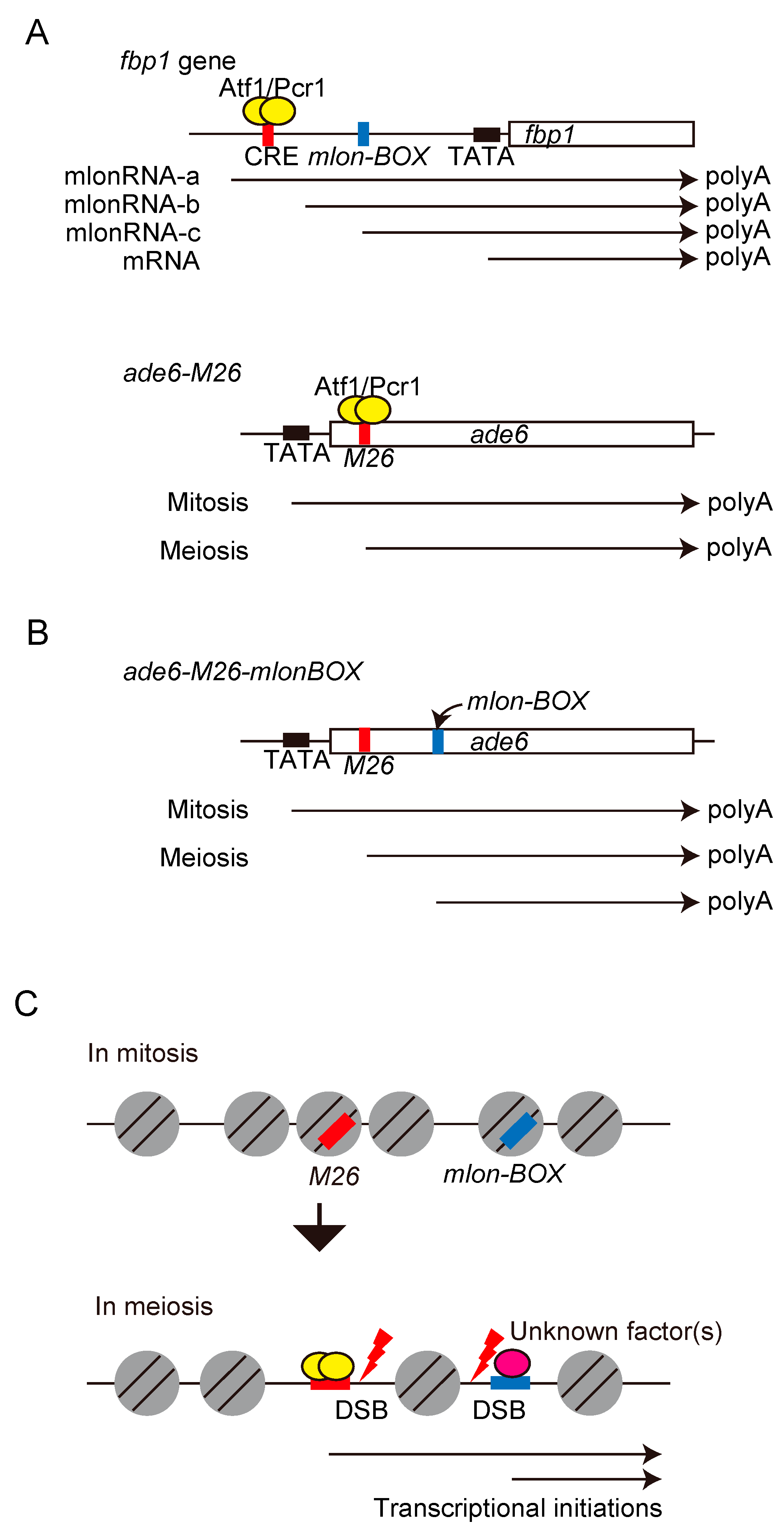{kind=link}
Abstract
:1. Introduction
2. ade6-M26, a Meiotic Hotspot Created by a Mono-Nucleotide Substitution Mutation
3. Transcription Factor Required for Meiotic Recombination at M26
4. Chromatin Modulation Mechanisms Involved in the Regulation of ade6-M26 Meiotic Recombination Hotspot
5. Meiotic Double-Strand Break Formation around M26 Mutation Site
6. Induction of Meiotic DSB via the Expression of Long ncRNA
7. Summary and Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Láscarez-Lagunas, L.; Martinez-Garcia, M.; Colaiácovo, M. SnapShot: Meiosis—Prophase I. Cell 2020, 181, 1442–1442.e1. [Google Scholar] [CrossRef] [PubMed]
- Zickler, D.; Kleckner, N. Meiotic chromosomes: Integrating structure and function. Annu. Rev. Genet. 1999, 33, 603–754. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S.; Neale, M.J. Initiation of meiotic recombination by formation of DNA double-strand breaks: Mechanism and regulation. Biochem. Soc. Trans. 2006, 34, 523–525. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Peterson, S.E.; Jasin, M.; Keeney, S. Mechanisms of germ line genome instability. Semin. Cell Dev. Biol. 2016, 54, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Lam, I.; Keeney, S. Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016634. [Google Scholar] [CrossRef] [Green Version]
- Arora, C.; Kee, K.; Maleki, S.; Keeney, S. Antiviral protein Ski8 is a direct partner of Spo11 in meiotic DNA break formation, independent of its cytoplasmic role in RNA metabolism. Mol. Cell 2004, 13, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hooker, G.W.; Roeder, G.S. Saccharomyces cerevisiae Mer2, Mei4 and Rec114 form a complex required for meiotic double-strand break formation. Genetics 2006, 173, 1969–1981. [Google Scholar] [CrossRef] [Green Version]
- Maleki, S.; Neale, M.J.; Arora, C.; Henderson, K.A.; Keeney, S. Interactions between Mei4, Rec114, and other proteins required for meiotic DNA double-strand break formation in Saccharomyces cerevisiae. Chromosoma 2007, 116, 471–486. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, T.; Ito, M.; Kugou, K.; Yamada, S.; Furuichi, M.; Oda, A.; Yamada, T.; Hirota, K.; Masai, H.; Ohta, K. A central coupler for recombination initiation linking chromosome architecture to S phase checkpoint. Mol. Cell 2012, 47, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Panizza, S.; Mendoza, M.A.; Berlinger, M.; Huang, L.; Nicolas, A.; Shirahige, K.; Klein, F. Spo11-accessory proteins link double-strand break sites to the chromosome axis in early meiotic recombination. Cell 2011, 146, 372–383. [Google Scholar] [CrossRef]
- Sasanuma, H.; Hirota, K.; Fukuda, T.; Kakusho, N.; Kugou, K.; Kawasaki, Y.; Shibata, T.; Masai, H.; Ohta, K. Cdc7-dependent phosphorylation of Mer2 facilitates initiation of yeast meiotic recombination. Genes Dev. 2008, 22, 398–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pommier, Y.; Sun, Y.; Huang, S.N.; Nitiss, J.L. Roles of eukaryotic topoisomerases in transcription, replication and genomic stability. Nat. Rev. Mol. Cell Biol. 2016, 17, 703–721. [Google Scholar] [CrossRef] [PubMed]
- Borde, V.; Lin, W.; Novikov, E.; Petrini, J.H.; Lichten, M.; Nicolas, A. Association of Mre11p with double-strand break sites during yeast meiosis. Mol. Cell 2004, 13, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Farah, J.A.; Cromie, G.A.; Smith, G.R. Ctp1 and Exonuclease 1, alternative nucleases regulated by the MRN complex, are required for efficient meiotic recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 9356–9361. [Google Scholar] [CrossRef] [Green Version]
- Mimitou, E.P.; Yamada, S.; Keeney, S. A global view of meiotic double-strand break end resection. Science 2017, 355, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjha, L.; Howard, S.M.; Cejka, P. Main steps in DNA double-strand break repair: An introduction to homologous recombination and related processes. Chromosoma 2018, 127, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Shibata, T.; Iwasaki, W.; Hirota, K. The intrinsic ability of double-stranded DNA to carry out D-loop and R-loop formation. Comput. Struct. Biotechnol. J. 2020, 18, 3350–3360. [Google Scholar] [CrossRef]
- Shinohara, A.; Shinohara, M. Roles of RecA homologues Rad51 and Dmc1 during meiotic recombination. Cytogenet. Genome Res. 2004, 107, 201–207. [Google Scholar] [CrossRef]
- Haber, J.E. DNA Repair: The Search for Homology. Bioessays 2018, 40, e1700229. [Google Scholar] [CrossRef]
- Bannister, L.A.; Pezza, R.J.; Donaldson, J.R.; de Rooij, D.G.; Schimenti, K.J.; Camerini-Otero, R.D.; Schimenti, J.C. A dominant, recombination-defective allele of Dmc1 causing male-specific sterility. PLoS Biol. 2007, 5, e105. [Google Scholar] [CrossRef]
- Cao, D.; Shi, F.; Guo, C.; Liu, Y.; Lin, Z.; Zhang, J.; Li, R.H.W.; Yao, Y.; Liu, K.; Ng, E.H.Y.; et al. A pathogenic DMC1 frameshift mutation causes nonobstructive azoospermia but not primary ovarian insufficiency in humans. Mol. Hum. Reprod. 2021, 27, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yang, H.; Pavletich, N.P. Mechanism of homologous recombination from the RecA-ssDNA/dsDNA structures. Nature 2008, 453, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Cunningham, R.P.; DasGupta, C.; Radding, C.M. Homologous pairing in genetic recombination: Complexes of recA protein and DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 5100–5104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allers, T.; Lichten, M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 2001, 106, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Bishop, D.K.; Zickler, D. Early decision; meiotic crossover interference prior to stable strand exchange and synapsis. Cell 2004, 117, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Cromie, G.A.; Hyppa, R.W.; Taylor, A.F.; Zakharyevich, K.; Hunter, N.; Smith, G.R. Single Holliday junctions are intermediates of meiotic recombination. Cell 2006, 127, 1167–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, T.J.; Garcia, V.; Neale, M.J. Meiotic DSB patterning: A multifaceted process. Cell Cycle 2016, 15, 13–21. [Google Scholar] [CrossRef] [Green Version]
- de Massy, B. Initiation of meiotic recombination: How and where? Conservation and specificities among eukaryotes. Annu. Rev. Genet. 2013, 47, 563–599. [Google Scholar] [CrossRef]
- Ergoren, M.C. The Control of Meiotic Recombination in the Human Genome. Crit. Rev. Eukaryot. Gene Expr. 2018, 28, 187–204. [Google Scholar] [CrossRef]
- Tock, A.J.; Henderson, I.R. Hotspots for Initiation of Meiotic Recombination. Front. Genet. 2018, 9, 521. [Google Scholar] [CrossRef]
- Blitzblau, H.G.; Bell, G.W.; Rodriguez, J.; Bell, S.P.; Hochwagen, A. Mapping of meiotic single-stranded DNA reveals double-stranded-break hotspots near centromeres and telomeres. Curr. Biol. 2007, 17, 2003–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhler, C.; Borde, V.; Lichten, M. Mapping meiotic single-strand DNA reveals a new landscape of DNA double-strand breaks in Saccharomyces cerevisiae. PLoS Biol. 2007, 5, e324. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.R.; Sasaki, M.; Milman, N.; Keeney, S.; Smith, G.R. Evolutionarily diverse determinants of meiotic DNA break and recombination landscapes across the genome. Genome. Res. 2014, 24, 1650–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerton, J.L.; DeRisi, J.; Shroff, R.; Lichten, M.; Brown, P.O.; Petes, T.D. Global mapping of meiotic recombination hotspots and coldspots in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2000, 97, 11383–11390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Sasaki, M.; Kniewel, R.; Murakami, H.; Blitzblau, H.G.; Tischfield, S.E.; Zhu, X.; Neale, M.J.; Jasin, M.; Socci, N.D.; et al. A hierarchical combination of factors shapes the genome-wide topography of yeast meiotic recombination initiation. Cell 2011, 144, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Wahls, W.P.; Siegel, E.R.; Davidson, M.K. Meiotic recombination hotspots of fission yeast are directed to loci that express non-coding RNA. PLoS ONE 2008, 3, e2887. [Google Scholar] [CrossRef] [Green Version]
- Yamada, S.; Okamura, M.; Oda, A.; Murakami, H.; Ohta, K.; Yamada, T. Correlation of Meiotic DSB Formation and Transcription Initiation Around Fission Yeast Recombination Hotspots. Genetics 2017, 206, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Wolffe, A.P. Nucleosome positioning and modification: Chromatin structures that potentiate transcription. Trends Biochem. Sci. 1994, 19, 240–244. [Google Scholar] [CrossRef]
- Wolffe, A.P. Histones, nucleosomes and the roles of chromatin structure in transcriptional control. Biochem. Soc. Trans. 1997, 25, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Modliszewski, J.L.; Copenhaver, G.P. Meiotic recombination gets stressed out: CO frequency is plastic under pressure. Curr. Opin. Plant. Biol. 2017, 36, 95–102. [Google Scholar] [CrossRef]
- Fisher, C.R. Enzymology of the pigmented adenine-requiring mutants of Saccharomyces and Schizosaccharomyces. Biochem. Biophys. Res. Commun 1969, 34, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Gutz, H. Site Specific Induction of Gene Conversion in Schizosaccharomyces pombe. Genetics 1971, 69, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Ponticelli, A.S.; Sena, E.P.; Smith, G.R. Genetic and physical analysis of the M26 recombination hotspot of Schizosaccharomyces pombe. Genetics 1988, 119, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Szankasi, P.; Heyer, W.D.; Schuchert, P.; Kohli, J. DNA sequence analysis of the ade6 gene of Schizosaccharomyces pombe. Wild-type and mutant alleles including the recombination host spot allele ade6-M26. J. Mol. Biol. 1988, 204, 917–925. [Google Scholar] [CrossRef]
- De Veaux, L.C.; Hoagland, N.A.; Smith, G.R. Seventeen complementation groups of mutations decreasing meiotic recombination in Schizosaccharomyces pombe. Genetics 1992, 130, 251–262. [Google Scholar] [CrossRef]
- Ponticelli, A.S.; Smith, G.R. Meiotic recombination-deficient mutants of Schizosaccharomyces pombe. Genetics 1989, 123, 45–54. [Google Scholar] [CrossRef]
- Schuchert, P.; Kohli, J. The Ade6-M26 Mutation of Schizosaccharomyces pombe Increases the Frequency of Crossing over. Genetics 1988, 119, 507–515. [Google Scholar] [CrossRef]
- Kon, N.; Krawchuk, M.D.; Warren, B.G.; Smith, G.R.; Wahls, W.P. Transcription factor Mts1/Mts2 (Atf1/Pcr1, Gad7/Pcr1) activates the M26 meiotic recombination hotspot in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1997, 94, 13765–13770. [Google Scholar] [CrossRef] [Green Version]
- Kon, N.; Schroeder, S.C.; Krawchuk, M.D.; Wahls, W.P. Regulation of the Mts1-Mts2-dependent ade6-M26 meiotic recombination hot spot and developmental decisions by the Spc1 mitogen-activated protein kinase of fission yeast. Mol. Cell Biol. 1998, 18, 7575–7583. [Google Scholar] [CrossRef] [Green Version]
- Wahls, W.P.; Smith, G.R. A heteromeric protein that binds to a meiotic homologous recombination hot spot: Correlation of binding and hot spot activity. Genes Dev. 1994, 8, 1693–1702. [Google Scholar] [CrossRef]
- Kanoh, J.; Watanabe, Y.; Ohsugi, M.; Iino, Y.; Yamamoto, M. Schizosaccharomyces pombe gad7+ encodes a phosphoprotein with a bZIP domain, which is required for proper G1 arrest and gene expression under nitrogen starvation. Genes Cells 1996, 1, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Shiozaki, K.; Russell, P. Conjugation, meiosis, and the osmotic stress response are regulated by Spc1 kinase through Atf1 transcription factor in fission yeast. Genes Dev. 1996, 10, 2276–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, T.; Toda, T.; Kominami, K.; Kohnosu, A.; Yanagida, M.; Jones, N. Schizosaccharomyces pombe atf1+ encodes a transcription factor required for sexual development and entry into stationary phase. EMBO J. 1995, 14, 6193–6208. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Yamamoto, M. Schizosaccharomyces pombe pcr1+ encodes a CREB/ATF protein involved in regulation of gene expression for sexual development. Mol. Cell Biol. 1996, 16, 704–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M. Regulation of meiosis in fission yeast. Cell Struct. Funct. 1996, 21, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Adachi, A.; Senmatsu, S.; Asada, R.; Abe, T.; Hoffman, C.S.; Ohta, K.; Hirota, K. Interplay between chromatin modulators and histone acetylation regulates the formation of accessible chromatin in the upstream regulatory region of fission yeast fbp1. Genes Genet. Syst. 2018, 92, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Hasemi, T.; Yamada, T.; Mizuno, K.I.; Hoffman, C.S.; Shibata, T.; Ohta, K. Fission yeast global repressors regulate the specificity of chromatin alteration in response to distinct environmental stresses. Nucleic Acids Res. 2004, 32, 855–862. [Google Scholar] [CrossRef]
- Hirota, K.; Hoffman, C.S.; Shibata, T.; Ohta, K. Fission yeast Tup1-like repressors repress chromatin remodeling at the fbp1+ promoter and the ade6-M26 recombination hotspot. Genetics 2003, 165, 505–515. [Google Scholar] [CrossRef]
- Hirota, K.; Miyoshi, T.; Kugou, K.; Hoffman, C.S.; Shibata, T.; Ohta, K. Stepwise chromatin remodelling by a cascade of transcription initiation of non-coding RNAs. Nature 2008, 456, 130–134. [Google Scholar] [CrossRef]
- Janoo, R.T.; Neely, L.A.; Braun, B.R.; Whitehall, S.K.; Hoffman, C.S. Transcriptional regulators of the Schizosaccharomyces pombe fbp1 gene include two redundant Tup1p-like corepressors and the CCAAT binding factor activation complex. Genetics 2001, 157, 1205–1215. [Google Scholar] [CrossRef]
- Koda, W.; Senmatsu, S.; Abe, T.; Hoffman, C.S.; Hirota, K. Reciprocal stabilization of transcription factor binding integrates two signaling pathways to regulate fission yeast fbp1 transcription. Nucleic Acids Res. 2021, 49, 9809–9820. [Google Scholar] [CrossRef]
- Neely, L.A.; Hoffman, C.S. Protein kinase A and mitogen-activated protein kinase pathways antagonistically regulate fission yeast fbp1 transcription by employing different modes of action at two upstream activation sites. Mol. Cell Biol. 2000, 20, 6426–6434. [Google Scholar] [CrossRef]
- Takemata, N.; Oda, A.; Yamada, T.; Galipon, J.; Miyoshi, T.; Suzuki, Y.; Sugano, S.; Hoffman, C.S.; Hirota, K.; Ohta, K. Local potentiation of stress-responsive genes by upstream noncoding transcription. Nucleic Acids Res. 2016, 44, 5174–5189. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Mir, L.; Fraile, R.; Ayté, J.; Hidalgo, E. Phosphorylation of the Transcription Factor Atf1 at Multiple Sites by the MAP Kinase Sty1 Controls Homologous Recombination and Transcription. J. Mol. Biol. 2020, 432, 5430–5446. [Google Scholar] [CrossRef] [PubMed]
- Senmatsu, S.; Asada, R.; Oda, A.; Hoffman, C.S.; Ohta, K.; Hirota, K. lncRNA transcription induces meiotic recombination through chromatin remodelling in fission yeast. Commun. Biol. 2021, 4, 295. [Google Scholar] [CrossRef]
- Ponticelli, A.S.; Smith, G.R. Chromosomal context dependence of a eukaryotic recombinational hot spot. Proc. Natl. Acad. Sci. USA 1992, 89, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Zahn-Zabal, M.; Lehmann, E.; Kohli, J. Hot spots of recombination in fission yeast: Inactivation of the M26 hot spot by deletion of the ade6 promoter and the novel hotspot ura4-aim. Genetics 1995, 140, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Steiner, W.W.; Davidow, P.A.; Bagshaw, A.T. Important characteristics of sequence-specific recombination hotspots in Schizosaccharomyces pombe. Genetics 2011, 187, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, W.W.; Steiner, E.M.; Girvin, A.R.; Plewik, L.E. Novel nucleotide sequence motifs that produce hotspots of meiotic recombination in Schizosaccharomyces pombe. Genetics 2009, 182, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Protacio, R.U.; Mukiza, T.O.; Davidson, M.K.; Wahls, W.P. Molecular mechanisms for environmentally induced and evolutionarily rapid redistribution (plasticity) of meiotic recombination. Genetics 2022, 220, iyab212. [Google Scholar] [CrossRef]
- Wahls, W.P.; Davidson, M.K. Discrete DNA sites regulate global distribution of meiotic recombination. Trends Genet. 2010, 26, 202–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukiza, T.O.; Protacio, R.U.; Davidson, M.K.; Steiner, W.W.; Wahls, W.P. Diverse DNA Sequence Motifs Activate Meiotic Recombination Hotspots Through a Common Chromatin Remodeling Pathway. Genetics 2019, 213, 789–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, P.A.; Sterner, D.E.; Duggan, L.J.; Workman, J.L.; Berger, S.L. The SAGA unfolds: Convergence of transcription regulators in chromatin-modifying complexes. Trends Cell Biol. 1998, 8, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.H.; Allis, C.D. Roles of histone acetyltransferases and deacetylases in gene regulation. Bioessays 1998, 20, 615–626. [Google Scholar] [CrossRef]
- Travers, A. An engine for nucleosome remodeling. Cell 1999, 96, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Chandy, M.; Gutierrez, J.L.; Prochasson, P.; Workman, J.L. SWI/SNF displaces SAGA-acetylated nucleosomes. Eukaryot. Cell 2006, 5, 1738–1747. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.H.; Awad, S.; Prochasson, P. The Swi2/Snf2 bromodomain is required for the displacement of SAGA and the octamer transfer of SAGA-acetylated nucleosomes. J. Biol. Chem 2006, 281, 18126–18134. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Mizuno, K.; Hirota, K.; Kon, N.; Wahls, W.P.; Hartsuiker, E.; Murofushi, H.; Shibata, T.; Ohta, K. Roles of histone acetylation and chromatin remodeling factor in a meiotic recombination hotspot. EMBO J. 2004, 23, 1792–1803. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, K.; Emura, Y.; Baur, M.; Kohli, J.; Ohta, K.; Shibata, T. The meiotic recombination hot spot created by the single-base substitution ade6-M26 results in remodeling of chromatin structure in fission yeast. Genes Dev. 1997, 11, 876–886. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Mizuno, K.; Shibata, T.; Ohta, K. Distinct chromatin modulators regulate the formation of accessible and repressive chromatin at the fission yeast recombination hotspot ade6-M26. Mol. Biol. Cell 2008, 19, 1162–1173. [Google Scholar] [CrossRef]
- Yamada, S.; Kugou, K.; Ding, D.Q.; Fujita, Y.; Hiraoka, Y.; Murakami, H.; Ohta, K.; Yamada, T. The histone variant H2A.Z promotes initiation of meiotic recombination in fission yeast. Nucleic Acids Res. 2018, 46, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, A.J.; Wang, H.P.; Protacio, R.U.; Davidson, M.K.; Tackett, A.J.; Wahls, W.P. Chromatin-mediated regulators of meiotic recombination revealed by proteomics of a recombination hotspot. Epigenetics Chromatin 2018, 11, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meselson, M.S.; Radding, C.M. A general model for genetic recombination. Proc. Natl. Acad. Sci. USA 1975, 72, 358–361. [Google Scholar] [CrossRef] [Green Version]
- Szostak, J.W.; Orr-Weaver, T.L.; Rothstein, R.J.; Stahl, F.W. The double-strand-break repair model for recombination. Cell 1983, 33, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Treco, D.; Schultes, N.P.; Szostak, J.W. Double-strand breaks at an initiation site for meiotic gene conversion. Nature 1989, 338, 87–90. [Google Scholar] [CrossRef]
- Roeder, G.S. Meiotic chromosomes: It takes two to tango. Genes Dev. 1997, 11, 2600–2621. [Google Scholar] [CrossRef] [Green Version]
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Bergerat, A.; de Massy, B.; Gadelle, D.; Varoutas, P.C.; Nicolas, A.; Forterre, P. An atypical topoisomerase II from Archaea with implications for meiotic recombination. Nature 1997, 386, 414–417. [Google Scholar] [CrossRef]
- Cervantes, M.D.; Farah, J.A.; Smith, G.R. Meiotic DNA breaks associated with recombination in S. pombe. Mol. Cell 2000, 5, 883–888. [Google Scholar] [CrossRef]
- Alani, E.; Padmore, R.; Kleckner, N. Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell 1990, 61, 419–436. [Google Scholar] [CrossRef]
- Cao, L.; Alani, E.; Kleckner, N. A pathway for generation and processing of double-strand breaks during meiotic recombination in S. cerevisiae. Cell 1990, 61, 1089–1101. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S.; Kleckner, N. Covalent protein-DNA complexes at the 5′ strand termini of meiosis-specific double-strand breaks in yeast. Proc. Natl. Acad. Sci. USA 1995, 92, 11274–11278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.A.; Schreckhise, R.W.; Steiner, W.W.; Smith, G.R. Meiotic recombination remote from prominent DNA break sites in S. pombe. Mol. Cell 2002, 9, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Steiner, W.W.; Schreckhise, R.W.; Smith, G.R. Meiotic DNA breaks at the S. pombe recombination hot spot M26. Mol. Cell 2002, 9, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkipeddi, S.M.K.; Velleca, A.J.; Carone, D.M. Probing the function of long noncoding RNAs in the nucleus. Chromosome Res. 2020, 28, 87–110. [Google Scholar] [CrossRef] [PubMed]
- Sanchez Calle, A.; Kawamura, Y.; Yamamoto, Y.; Takeshita, F.; Ochiya, T. Emerging roles of long non-coding RNA in cancer. Cancer Sci. 2018, 109, 2093–2100. [Google Scholar] [CrossRef] [Green Version]
- Akbari, A.; Abbasi, S.; Borumandnia, N.; Eshkiki, Z.S.; Sedaghat, M.; Tabaeian, S.P.; Kashani, A.F.; Talebi, A. Epigenetic regulation of gastrointestinal cancers mediated by long non-coding RNA. Cancer Biomark. 2022; 1–19, Prepress. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, X.; Lai, K.; Zhou, R.; Chen, Z.; Yang, Z.; Gao, X. Discovery of the hidden coding information in cancers: Mechanisms and biological functions. Int. J. Cancer, 2022; Prepress. [Google Scholar] [CrossRef]
- Calin, G.A.; Liu, C.G.; Ferracin, M.; Hyslop, T.; Spizzo, R.; Sevignani, C.; Fabbri, M.; Cimmino, A.; Lee, E.J.; Wojcik, S.E.; et al. Ultraconserved regions encoding ncRNAs are altered in human leukemias and carcinomas. Cancer Cell 2007, 12, 215–229. [Google Scholar] [CrossRef]
- Adelman, K. Nascent transcription as a predictor and driver of histone modifications. Nat. Genet. 2022, 54, 223–224. [Google Scholar] [CrossRef]
- Wang, Z.; Chivu, A.G.; Choate, L.A.; Rice, E.J.; Miller, D.C.; Chu, T.; Chou, S.P.; Kingsley, N.B.; Petersen, J.L.; Finno, C.J.; et al. Prediction of histone post-translational modification patterns based on nascent transcription data. Nat. Genet. 2022, 54, 295–305. [Google Scholar] [CrossRef]
- An, B.; Kameda, T.; Imamura, T. The evolutionary acquisition and mode of functions of promoter-associated non-coding RNAs (pancRNAs) for mammalian development. Essays Biochem. 2021, 65, 697–708. [Google Scholar] [CrossRef]
- Asada, R.; Hirota, K. Multi-Layered Regulations on the Chromatin Architectures: Establishing the Tight and Specific Responses of Fission Yeast fbp1 Gene Transcription. Biomolecules 2022, 12, 1642. [Google Scholar] [CrossRef]
- Vassarotti, A.; Friesen, J.D. Isolation of the fructose-1,6-bisphosphatase gene of the yeast Schizosaccharomyces pombe. Evidence for transcriptional regulation. J. Biol. Chem. 1985, 260, 6348–6353. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Ohta, K. Transcription of mRNA-type long non-coding RNAs (mlonRNAs) disrupts chromatin array. Commun. Integr. Biol. 2009, 2, 25–26. [Google Scholar] [CrossRef] [PubMed]
- Oda, A.; Takemata, N.; Hirata, Y.; Miyoshi, T.; Suzuki, Y.; Sugano, S.; Ohta, K. Dynamic transition of transcription and chromatin landscape during fission yeast adaptation to glucose starvation. Genes Cells 2015, 20, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Senmatsu, S.; Asada, R.; Abe, T.; Hoffman, C.S.; Ohta, K.; Hirota, K. lncRNA transcriptional initiation induces chromatin remodeling within a limited range in the fission yeast fbp1 promoter. Sci. Rep. 2019, 9, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cromie, G.A.; Rubio, C.A.; Hyppa, R.W.; Smith, G.R. A natural meiotic DNA break site in Schizosaccharomyces pombe is a hotspot of gene conversion, highly associated with crossing over. Genetics 2005, 169, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Steiner, W.W.; Shibata, T.; Ohta, K. Multiple modes of chromatin configuration at natural meiotic recombination hot spots in fission yeast. Eukaryot. Cell 2007, 6, 2072–2080. [Google Scholar] [CrossRef] [PubMed]
- Plough, H.H. The Effect of Temperature on Linkage in the Second Chromosome of Drosophila. Proc. Natl. Acad. Sci. USA 1917, 3, 553–555. [Google Scholar] [CrossRef] [Green Version]
- White, M.A.; Dominska, M.; Petes, T.D. Transcription factors are required for the meiotic recombination hotspot at the HIS4 locus in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1993, 90, 6621–6625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieczkowski, P.A.; Dominska, M.; Buck, M.J.; Gerton, J.L.; Lieb, J.D.; Petes, T.D. Global analysis of the relationship between the binding of the Bas1p transcription factor and meiosis-specific double-strand DNA breaks in Saccharomyces cerevisiae. Mol. Cell Biol. 2006, 26, 1014–1027. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirota, K. Regulation Mechanisms of Meiotic Recombination Revealed from the Analysis of a Fission Yeast Recombination Hotspot ade6-M26. Biomolecules 2022, 12, 1761. https://doi.org/10.3390/biom12121761
Hirota K. Regulation Mechanisms of Meiotic Recombination Revealed from the Analysis of a Fission Yeast Recombination Hotspot ade6-M26. Biomolecules. 2022; 12(12):1761. https://doi.org/10.3390/biom12121761
Chicago/Turabian StyleHirota, Kouji. 2022. "Regulation Mechanisms of Meiotic Recombination Revealed from the Analysis of a Fission Yeast Recombination Hotspot ade6-M26" Biomolecules 12, no. 12: 1761. https://doi.org/10.3390/biom12121761