
Immunologic Roles of Hyaluronan in Dermal Wound Healing



Abstract
:1. Introduction
Overview of HA
2. Immune System
3. Innate Immunity
3.1. Mast Cells
3.2. Macrophages
3.3. Dendritic/Langerhans Cells
3.4. Other Cell Populations
4. Adaptive Immunity
4.1. T Lymphocytes
4.2. CD4+ T lymphocytes
4.3. Gamma Delta T (γδ T) Cells
4.4. B Cells
5. Concluding Thoughts
Funding
Conflicts of Interest
Glossary
| DAMP | damage-associated molecular pattern |
| DETC | dendritic epidermal T cells |
| ECM | extracellular matrix |
| γδ T | gamma delta T cell |
| HA | hyaluronic acid/hyaluronan |
| HMW-HA | high molecular-weight hyaluronic acid |
| IL | interleukin |
| LMW-HA | low molecular-weight hyaluronic acid |
| MW | molecular weight |
| NKT | natural killer T |
| PAMP | pathogen-associated molecular pattern |
| PRR | pattern recognition receptor |
| RHAMM | receptor for HA-mediated motility |
| TCR | T cell receptor |
| TGF-β | transforming growth factor beta |
| Th1/2 | T helper type 1/2 |
| TLR | toll-like receptor |
| TNF-α | tumor necrosis factor alpha |
References
- Bayat, A.; McGrouther, D.A.; Ferguson, M.W. Skin scarring. BMJ 2003, 326, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Reinke, J.; Sorg, H. Wound Repair and Regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singampalli, K.L.; Balaji, S.; Wang, X.; Parikh, U.M.; Kaul, A.; Gilley, J.; Birla, R.K.; Bollyky, P.L.; Keswani, S.G. The Role of an IL-10/Hyaluronan Axis in Dermal Wound Healing. Front. Cell Dev. Biol. 2020, 8, 636. [Google Scholar] [CrossRef] [PubMed]
- Litwiniuk, M.; Krejner, A.; Speyrer, M.S.; Gauto, A.R.; Grzela, T. Hyaluronic Acid in Inflammation and Tissue Regeneration. Wounds 2016, 28, 78–88. [Google Scholar]
- Inokoshi, Y.; Tanino, Y.; Wang, X.; Sato, S.; Fukuhara, N.; Nikaido, T.; Fukuhara, A.; Saito, J.; Frevert, C.W.; Munakata, M. Clinical significance of serum hyaluronan in chronic fibrotic interstitial pneumonia. Respirology 2013, 18, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Reed, R.K.; Lilja, K.; Laurent, T.C. Hyaluronan in the rat with special reference to the skin. Acta Physiol. Scand. 1988, 134, 405–411. [Google Scholar] [CrossRef]
- Tammi, R.; Ripellino, J.A.; Margolis, R.U.; Tammi, M. Localization of Epidermal Hyaluronic Acid Using the Hyaluronate Binding Region of Cartilage Proteoglycan as a Specific Probe. J. Investig. Dermatol. 1988, 90, 412–414. [Google Scholar] [CrossRef]
- Nagy, N.; Sunkari, V.; Kaber, G.; Hasbun, S.; Lam, D.N.; Speake, C.; Sanda, S.; McLaughlin, T.L.; Wight, T.N.; Long, S.R.; et al. Hyaluronan levels are increased systemically in human type 2 but not type 1 diabetes independently of glycemic control. Matrix Biol. 2019, 80, 46–58. [Google Scholar] [CrossRef]
- Ruppert, S.M.; Hawn, T.R.; Arrigoni, A.; Wight, T.N.; Bollyky, P.L. Tissue integrity signals communicated by high-molecular weight hyaluronan and the resolution of inflammation. Immunol. Res. 2014, 58, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.D.; Horton, M.R. Threat Matrix: Low-Molecular-Weight Hyaluronan (HA) as a Danger Signal. Immunol. Res. 2005, 31, 207–218. [Google Scholar] [CrossRef]
- Liu, M.; Tolg, C.; Turley, E. Dissecting the Dual Nature of Hyaluronan in the Tumor Microenvironment. Front. Immunol. 2019, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Wolny, P.M.; Banerji, S.; Gounou, C.; Brisson, A.R.; Day, A.; Jackson, D.G.; Richter, R.P. Analysis of CD44-Hyaluronan Interactions in an Artificial Membrane System: Insights into the Distinct Binding Properties of High and Low Molecular Weight Hyaluronan. J. Biol. Chem. 2010, 285, 30170–30180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerji, S.; Wright, A.J.; Noble, M.; Mahoney, D.J.; Campbell, I.D.; Day, A.; Jackson, D.G. Structures of the Cd44–hyaluronan complex provide insight into a fundamental carbohydrate-protein interaction. Nat. Struct. Mol. Biol. 2007, 14, 234–239. [Google Scholar] [CrossRef]
- Sapudom, J.; Ullm, F.; Martin, S.; Kalbitzer, L.; Naab, J.; Möller, S.; Schnabelrauch, M.; Anderegg, U.; Schmidt, S.; Pompe, T. Molecular weight specific impact of soluble and immobilized hyaluronan on CD44 expressing melanoma cells in 3D collagen matrices. Acta Biomater. 2017, 50, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Sapudom, J.; Müller, C.D.; Nguyen, K.-T.; Martin, S.; Anderegg, U.; Pompe, T. Matrix Remodeling and Hyaluronan Production by Myofibroblasts and Cancer-Associated Fibroblasts in 3D Collagen Matrices. Gels 2020, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Kuschert, G.S.V.; Coulin, F.; Power, C.A.; Proudfoot, A.E.I.; Hubbard, R.E.; Hoogewerf, A.J.; Wells, T. Glycosaminoglycans Interact Selectively with Chemokines and Modulate Receptor Binding and Cellular Responses. Biochemistry 1999, 38, 12959–12968. [Google Scholar] [CrossRef] [PubMed]
- Webb, L.; Ehrengruber, M.U.; Clark-Lewis, I.; Baggiolini, M.; Rot, A. Binding to heparan sulfate or heparin enhances neutrophil responses to interleukin 8. Proc. Natl. Acad. Sci. USA 1993, 90, 7158–7162. [Google Scholar] [CrossRef] [Green Version]
- Pichert, A.; Samsonov, S.; Theisgen, S.; Thomas, L.H.; Baumann, L.; Schiller, J.; Beck-Sickinger, A.G.; Huster, D.; Pisabarro, M.T. Characterization of the interaction of interleukin-8 with hyaluronan, chondroitin sulfate, dermatan sulfate and their sulfated derivatives by spectroscopy and molecular modeling. Glycobiology 2011, 22, 134–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanko, S.P.; Tammi, M.I.; Tammi, R.H.; Wight, T.N. Hyaluronan-dependent pericellular matrix. Adv. Drug Deliv. Rev. 2007, 59, 1351–1365. [Google Scholar] [CrossRef] [Green Version]
- Abaskharoun, M.; Bellemare, M.; Lau, E.; Margolis, R.U. Expression of hyaluronan and the hyaluronan-binding proteoglycans neurocan, aggrecan, and versican by neural stem cells and neural cells derived from embryonic stem cells. Brain Res. 2010, 1327, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longaker, M.T.; Whitby, D.J.; Adzick, N.S.; Crombleholme, T.M.; Langer, J.C.; Duncan, B.W.; Bradley, S.M.; Stern, R.; Ferguson, M.W.; Harrison, M.R. Studies in fetal wound healing VI. Second and early third trimester fetal wounds demonstrate rapid collagen deposition without scar formation. J. Pediatr. Surg. 1990, 25, 63–69. [Google Scholar] [CrossRef]
- Krummel, T.M.; Nelson, J.M.; Diegelmann, R.F.; Lindblad, W.J.; Salzberg, A.M.; Greenfield, L.J.; Cohen, I.K. Fetal response to injury in the rabbit. J. Pediatr. Surg. 1987, 22, 640–644. [Google Scholar] [CrossRef]
- King, A.; Balaji, S.; Marsh, E.; Le, L.D.; Shaaban, A.F.; Crombleholme, T.M.; Keswani, S.G. Interleukin-10 regulates the fetal hyaluronan-rich extracellular matrix via a STAT3-dependent mechanism. J. Surg. Res. 2013, 184, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaji, S.; Wang, X.; King, A.; Le, L.D.; Bhattacharya, S.S.; Moles, C.M.; Butte, M.J.; Perez, V.A.D.J.; Liechty, K.W.; Wight, T.N.; et al. Interleukin-10–mediated regenerative postnatal tissue repair is dependent on regulation of hyaluronan metabolism via fibroblast-specific STAT3 signaling. FASEB J. 2017, 31, 868–881. [Google Scholar] [CrossRef] [Green Version]
- Lesley, J.; Howes, N.; Perschl, A.; Hyman, R. Hyaluronan binding function of CD44 is transiently activated on T cells during an in vivo immune response. J. Exp. Med. 1994, 180, 383–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degrendele, H.C.; Kosfiszer, M.; Estess, P.; Siegelman, M.H. CD44 activation and associated primary adhesion is inducible via T cell receptor stimulation. J. Immunol. 1997, 159, 2549–2553. [Google Scholar]
- Lee-Sayer, S.S.M.; Dong, Y.; Arif, A.A.; Olsson, M.; Brown, K.L.; Johnson, P. The Where, When, How, and Why of Hyaluronan Binding by Immune Cells. Front. Immunol. 2015, 6, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R.; Janeway, C., Jr. Innate immunity. N. Engl. J. Med. 2000, 343, 338–344. [Google Scholar] [CrossRef]
- Kaisho, T.; Akira, S. Toll-like receptor function and signaling. J. Allergy Clin. Immunol. 2006, 117, 979–987. [Google Scholar] [CrossRef]
- Kaisho, T.; Akira, S. Regulation of dendritic cell function through Toll-like receptors. Curr. Mol. Med. 2003, 3, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Kluwe, J.; Mencin, A.; Schwabe, R.F. Toll-like receptors, wound healing, and carcinogenesis. J. Mol. Med. 2008, 87, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasu, M.R.; Isseroff, R.R. Toll-Like Receptors in Wound Healing: Location, Accessibility, and Timing. J. Investig. Dermatol. 2012, 132, 1955–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Guo, S.; Ranzer, M.J.; DiPietro, L.A. Toll-Like Receptor 4 Has an Essential Role in Early Skin Wound Healing. J. Investig. Dermatol. 2013, 133, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Pivarcsi, A.; Bodai, L.; Réthi, B.; Kenderessy-Szabó, A.; Koreck, A.; Széll, M.; Beer, Z.; Bata-Csörgoő, Z.; Magócsi, M.; Rajnavölgyi, É.; et al. Expression and function of Toll-like receptors 2 and 4 in human keratinocytes. Int. Immunol. 2003, 15, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Ku, J.K.; Kwon, H.J.; Kim, M.-Y.; Kang, H.; Song, P.; Armstrong, C.A.; Ansel, J.C.; Kim, H.O.; Park, Y.M. Expression of Toll-Like Receptors in Verruca and Molluscum Contagiosum. J. Korean Med. Sci. 2008, 23, 307–314. [Google Scholar] [CrossRef]
- Miller, L.; Sorensen, O.E.; Liu, P.T.; Jalian, H.R.; Eshtiaghpour, D.; Behmanesh, B.E.; Chung, W.; Starner, T.; Kim, J.; Sieling, P.A.; et al. TGF-α Regulates TLR Expression and Function on Epidermal Keratinocytes. J. Immunol. 2005, 174, 6137–6143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, M.; Whittlesey, K.; Metcalfe, D.D.; Dastych, J. Human Mast Cells Express the Hyaluronic-Acid-Binding Isoform of CD44 and Adhere to Hyaluronic Acid. Clin. Immunol. 2000, 94, 173–178. [Google Scholar] [CrossRef]
- Jiang, D.; Liang, J.; Fan, J.; Yu, S.; Chen, S.; Luo, Y.; Prestwich, G.D.; Mascarenhas, M.M.; Garg, H.G.; Quinn, D.; et al. Regulation of lung injury and repair by Toll-like receptors and hyaluronan. Nat. Med. 2005, 11, 1173–1179. [Google Scholar] [CrossRef]
- Hauck, S.; Zager, P.; Halfter, N.; Wandel, E.; Torregrossa, M.; Kakpenova, A.; Rother, S.; Ordieres, M.; Räthel, S.; Berg, A.; et al. Collagen/hyaluronan based hydrogels releasing sulfated hyaluronan improve dermal wound healing in diabetic mice via reducing inflammatory macrophage activity. Bioact. Mater. 2021, 6, 4342–4359. [Google Scholar] [CrossRef]
- Termeer, C.; Benedix, F.; Sleeman, J.; Fieber, C.; Voith, U.; Ahrens, T.; Miyake, K.; Freudenberg, M.; Galanos, C.; Simon, J.C. Oligosaccharides of Hyaluronan Activate Dendritic Cells via Toll-like Receptor 4. J. Exp. Med. 2002, 195, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, J.; Renukaradhya, G.J.; Sriram, V.; Du, W.; Gervay-Hague, J.; Brutkiewicz, R. CD44 Differentially Activates Mouse NK T Cells and Conventional T Cells. J. Immunol. 2006, 177, 268–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollyky, P.L.; Falk, B.A.; Long, S.A.; Preisinger, A.; Braun, K.R.; Wu, R.P.; Nepom, G.T. CD44 costimulation promotes FoxP3+ regulatory T cell persistence and function via production of IL-2, IL-10, and TGF-beta. J. Immunol. 2009, 183, 2232–2241. [Google Scholar] [CrossRef] [Green Version]
- Jameson, J.M.; Cauvi, G.; Sharp, L.L.; Witherden, D.A.; Havran, W.L. γδ T cell-induced hyaluronan production by epithelial cells regulates inflammation. J. Exp. Med. 2005, 201, 1269–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwata, Y.; Yoshizaki, A.; Komura, K.; Shimizu, K.; Ogawa, F.; Hara, T.; Muroi, E.; Bae, S.; Takenaka, M.; Yukami, T.; et al. CD19, a Response Regulator of B Lymphocytes, Regulates Wound Healing through Hyaluronan-Induced TLR4 Signaling. Am. J. Pathol. 2009, 175, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Eggli, P.S.; Graber, W. Cytochemical Localization of Hyaluronan in Rat and Human Skin Mast Cell Granules. J. Investig. Dermatol. 1993, 100, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Weller, K.; Foitzik, K.; Paus, R.; Syska, W.; Maurer, M. Mast cells are required for normal healing of skin wounds in mice. FASEB J. 2006, 20, 2366–2368. [Google Scholar] [CrossRef] [PubMed]
- Grimbaldeston, M.; Nakae, S.; Kalesnikoff, J.; Tsai, M.; Galli, S.J. Mast cell–derived interleukin 10 limits skin pathology in contact dermatitis and chronic irradiation with ultraviolet B. Nat. Immunol. 2007, 8, 1095–1104. [Google Scholar] [CrossRef]
- Gordon, A.; Kozin, E.; Keswani, S.G.; Vaikunth, S.S.; Katz, A.B.; Zoltick, P.W.; Favata, M.; Ms, A.P.R.; Soslowsky, L.J.; Herlyn, M.; et al. Permissive environment in postnatal wounds induced by adenoviral-mediated overexpression of the anti-inflammatory cytokine interleukin-10 prevents scar formation. Wound Repair Regen. 2007, 16, 70–79. [Google Scholar] [CrossRef]
- Wick, G.; Grundtman, C.; Mayerl, C.; Wimpissinger, T.-F.; Feichtinger, J.; Zelger, B.; Sgonc, R.; Wolfram, D. The Immunology of Fibrosis. Annu. Rev. Immunol. 2013, 31, 107–135. [Google Scholar] [CrossRef] [Green Version]
- Kuang, D.-M.; Wu, Y.; Chen, N.; Cheng, J.; Zhuang, S.-M.; Zheng, L. Tumor-derived hyaluronan induces formation of immunosuppressive macrophages through transient early activation of monocytes. Blood 2007, 110, 587–595. [Google Scholar] [CrossRef]
- Aya, K.L.; Stern, R. Hyaluronan in wound healing: Rediscovering a major player. Wound Repair Regen. 2014, 22, 579–593. [Google Scholar] [CrossRef]
- Stern, R.; Maibach, H.I. Hyaluronan in skin: Aspects of aging and its pharmacologic modulation. Clin. Dermatol. 2008, 26, 106–122. [Google Scholar] [CrossRef]
- Mummert, M.E.; Mohamadzadeh, M.; Mummert, D.I.; Mizumoto, N.; Takashima, A. Development of a Peptide Inhibitor of Hyaluronan-Mediated Leukocyte Trafficking. J. Exp. Med. 2000, 192, 769–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovi, J.V.; He, L.-K.; DiPietro, L.A. Accelerated wound closure in neutrophil-depleted mice. J. Leukoc. Biol. 2003, 73, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Devalaraja, R.M.; Nanney, L.B.; Qian, Q.; Du, J.; Yu, Y.; Devalaraja, M.N.; Richmond, A. Delayed Wound Healing in CXCR2 Knockout Mice. J. Investig. Dermatol. 2000, 115, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, P.; Maiti, A.; Brown, K.; Li, R. A role for the cell adhesion molecule CD44 and sulfation in leukocyte–endothelial cell adhesion during an inflammatory response? Biochem. Pharmacol. 2000, 59, 455–465. [Google Scholar] [CrossRef]
- Degrendele, H.C.; Estess, P.; Picker, L.J.; Siegelman, M.H. CD44 and its ligand hyaluronate mediate rolling under physiologic flow: A novel lymphocyte-endothelial cell primary adhesion pathway. J. Exp. Med. 1996, 183, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- DeGrendele, H.C.; Estess, P.; Siegelman, M.H. Requirement for CD44 in Activated T Cell Extravasation into an Inflammatory Site. Science 1997, 278, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Muller, R.; Borgmann, S.; Brocker, E.-B.; Friedl, P. Amoeboid shape change and contact guidance: T-lymphocyte crawling through fibrillar collagen is independent of matrix remodeling by MMPs and other proteases. Blood 2003, 102, 3262–3269. [Google Scholar] [CrossRef] [Green Version]
- Serrador, J.M.; Nieto, M.; Alonso-Lebrero, J.L.; Del Pozo, M.; Calvo, J.; Furthmayr, H.; Schwartz-Albiez, R.; Lozano, F.; González-Amaro, R.; Sánchez-Mateos, P.; et al. CD43 interacts with moesin and ezrin and regulates its redistribution to the uropods of T lymphocytes at the cell-cell contacts. Blood 1998, 91, 4632–4644. [Google Scholar] [CrossRef] [PubMed]
- Serrador, J.M.; Alonso-Lebrero, J.L.; del Pozo, M.A.; Furthmayr, H.; Schwartz-Albiez, R.; Calvo, J.; Lozano, F.; Sánchez-Madrid, F. Moesin Interacts with the Cytoplasmic Region of Intercellular Adhesion Molecule-3 and Is Redistributed to the Uropod of T Lymphocytes during Cell Polarization. J. Cell Biol. 1997, 138, 1409–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrass, P.; Kinjyo, I.; Ng, L.G.; Reiner, S.L.; Puré, E.; Weninger, W. CD44 Mediates Successful Interstitial Navigation by Killer T Cells and Enables Efficient Antitumor Immunity. Immunity 2008, 29, 971–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sackstein, R.; Schatton, T.; Barthel, S.R. T-lymphocyte homing: An underappreciated yet critical hurdle for successful cancer immunotherapy. Lab. Investig. 2017, 97, 669–697. [Google Scholar] [CrossRef] [Green Version]
- Walling, B.L.; Kim, M. LFA-1 in T Cell Migration and Differentiation. Front. Immunol. 2018, 9, 952. [Google Scholar] [CrossRef] [Green Version]
- Kulidjian, A.A.; Issekutz, A.C.; Issekutz, T.B. Differential role of E-selectin and P-selectin in T lymphocyte migration to cutaneous inflammatory reactions induced by cytokines. Int. Immunol. 2002, 14, 751–760. [Google Scholar] [CrossRef]
- Hsieh, C.; Macatonia, S.; Tripp, C.; Wolf, S.; O’Garra, A.; Murphy, K. Development of TH1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science 1993, 260, 547–549. [Google Scholar] [CrossRef] [PubMed]
- del Pozo, M.A.; Sanchez-Mateos, P.; Nieto, M.; Sanchez-Madrid, F. Chemokines regulate cellular polarization and adhesion receptor redistribution during lymphocyte interaction with endothelium and extracellular matrix. Involvement of cAMP signaling pathway. J. Cell Biol. 1995, 131, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Oliphant, C.J.; Hwang, Y.Y.; Walker, J.A.; Salimi, M.; Wong, S.H.; Brewer, J.; Englezakis, A.; Barlow, J.L.; Hams, E.; Scanlon, S.T.; et al. MHCII-Mediated Dialog between Group 2 Innate Lymphoid Cells and CD4+ T Cells Potentiates Type 2 Immunity and Promotes Parasitic Helminth Expulsion. Immunity 2014, 41, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasteiger, G.; Fan, X.; Dikiy, S.; Lee, S.Y.; Rudensky, A.Y. Tissue residency of innate lymphoid cells in lymphoid and nonlymphoid organs. Science 2015, 350, 981–985. [Google Scholar] [CrossRef] [Green Version]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of Regulatory T Cell Development by the Transcription Factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firan, M.; Dhillon, S.; Estess, P.; Siegelman, M.H. Suppressor activity and potency among regulatory T cells is discriminated by functionally active CD44. Blood 2006, 107, 619–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonder, C.S.; Clark, S.R.; Norman, M.U.; Johnson, P.; Kubes, P. Use of CD44 by CD4+ Th1 and Th2 lymphocytes to roll and adhere. Blood 2006, 107, 4798–4806. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wu, W.; Wong, W.M.; Ward, E.; Thrasher, A.J.; Goldblatt, D.; Gustafsson, K. Human gamma delta T cells: A lymphoid lineage cell capable of professional phagocytosis. J. Immunol. 2009, 183, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Pitard, V.; Roumanes, D.; Lafarge, X.; Couzi, L.; Garrigue, I.; Lafon, M.E.; Déchanet-Merville, J. Long-term expansion of effector/memory Vδ2− γδ T cells is a specific blood signature of CMV infection. Blood 2008, 112, 1317–1324. [Google Scholar] [CrossRef] [Green Version]
- Sîrbulescu, R.F.; Boehm, C.K.; Soon, E.; Wilks, M.Q.; Ilies, I.; Yuan, H.; Maxner, B.; Chronos, N.; Kaittanis, C.; Normandin, M.; et al. Mature B cells accelerate wound healing after acute and chronic diabetic skin lesions. Wound Repair Regen. 2017, 25, 774–791. [Google Scholar] [CrossRef]


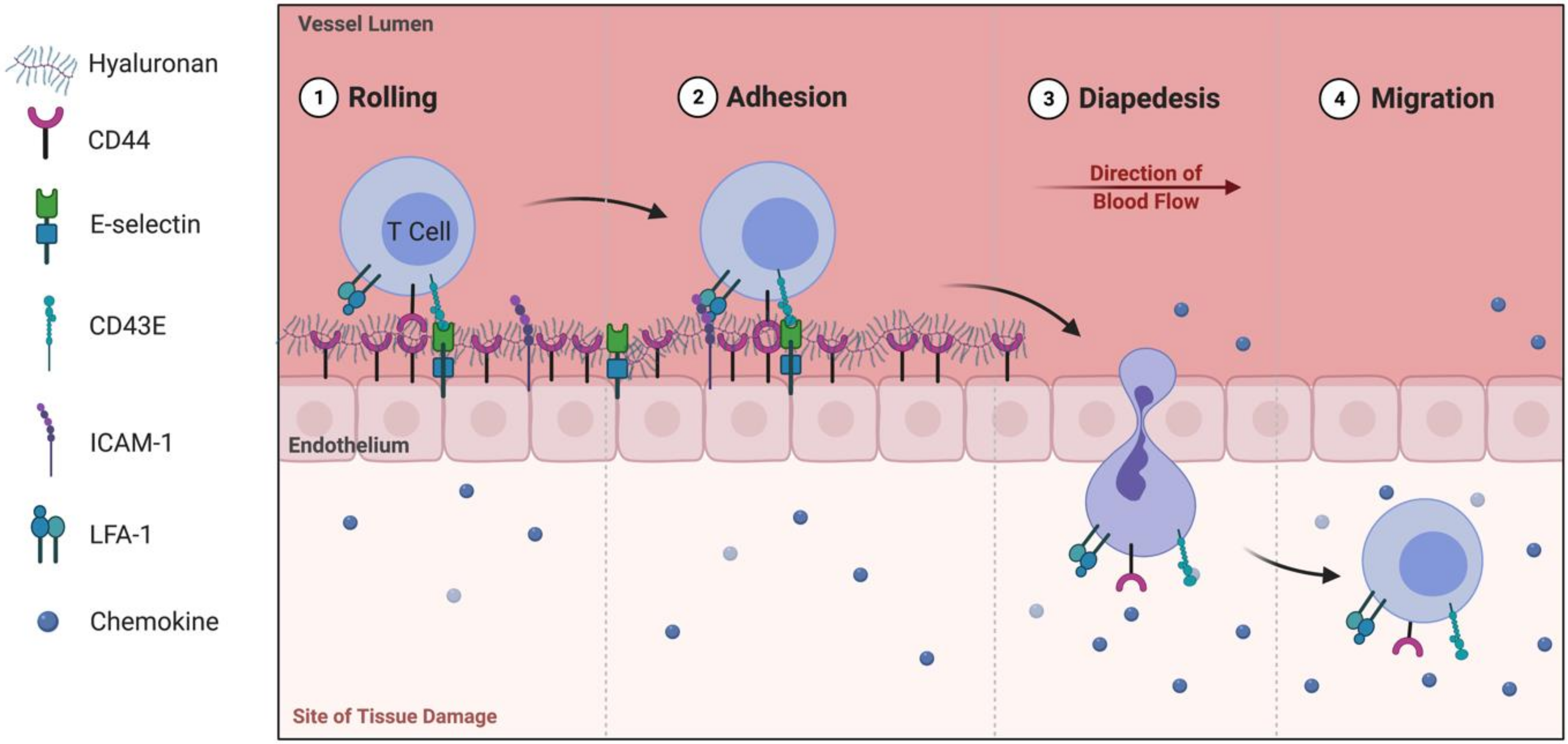{kind=link}
| Immune Subsystem | Cell Type | Subtype | Role with HA in Wound Healing |
|---|---|---|---|
| Innate | Mast cells | Facilitate attachment to HA [38] and promote anti-inflammatory wound milieu in concert with HA by way of IL-10 [4] | |
| Macrophages | M1 | Induced by LMW-HA during inflammatory stage of wound healing [39] | |
| M2 | Sulfated HA/collagen hydrogel improved murine diabetic wound healing by promoting M2 macrophages [40] | ||
| Dendritic/Langerhans cells | LMW-HA promotes maturation of dendritic cells via TLR 4 pathway during inflammatory phase of wound healing [41] | ||
| Natural killer T (NKT) cells | HA binds to CD44 receptors on the surface of NKT cells although downstream effects are unknown [42] | ||
| Adaptive | T lymphocytes | Antigen activation of T lymphocytes induces HA binding via CD44 which enables lymphocyte infiltration into inflamed tissue [28] | |
| CD4+ | HMW-HA amplifies Foxp3 expression of Tregs which stimulates production of IL-2 and immunosuppressive IL-10 and TGF-β [43] | ||
| γδ | Subpopulation known as DETCs secrete keratinocyte growth factors 1 and 2 which stimulate HA production and macrophage recruitment [44] | ||
| B cells | HA application to murine wounds attracts B cells to wound bed, stimulating production of IL-6 and TGF-β [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaul, A.; Short, W.D.; Keswani, S.G.; Wang, X. Immunologic Roles of Hyaluronan in Dermal Wound Healing. Biomolecules 2021, 11, 1234. https://doi.org/10.3390/biom11081234
Kaul A, Short WD, Keswani SG, Wang X. Immunologic Roles of Hyaluronan in Dermal Wound Healing. Biomolecules. 2021; 11(8):1234. https://doi.org/10.3390/biom11081234
Chicago/Turabian StyleKaul, Aditya, Walker D. Short, Sundeep G. Keswani, and Xinyi Wang. 2021. "Immunologic Roles of Hyaluronan in Dermal Wound Healing" Biomolecules 11, no. 8: 1234. https://doi.org/10.3390/biom11081234