
Pinocembrin Ameliorates Skin Fibrosis via Inhibiting TGF-β1 Signaling Pathway

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Keloid Tissue Samples
2.2. Separation and Culture of Keloid Fibroblasts
2.3. Separation and Culture of Primary Dermal Fibroblasts
2.4. Cell Counting Kit-8 (CCK-8) Assay
2.5. EdU Incorporation Assay
2.6. Wound-Healing Assay
2.7. Transwell Assays
2.8. Ex Vivo Explant Culture of Human Keloid Tissues
2.9. Quantitative Real-Time PCR (qRT-PCR)
2.10. Western Blot Analysis
2.11. Immunofluorescence Staining
2.12. Animals
2.13. Bleomycin-Induced Skin Fibrosis Model
2.14. Keloid Xenograft Mouse Model
2.15. Histological Examination
2.16. Hydroxyproline Content Determination
2.17. Flow Cytometric Analysis
2.18. Molecular Docking
2.19. Statistical Analysis
3. Results
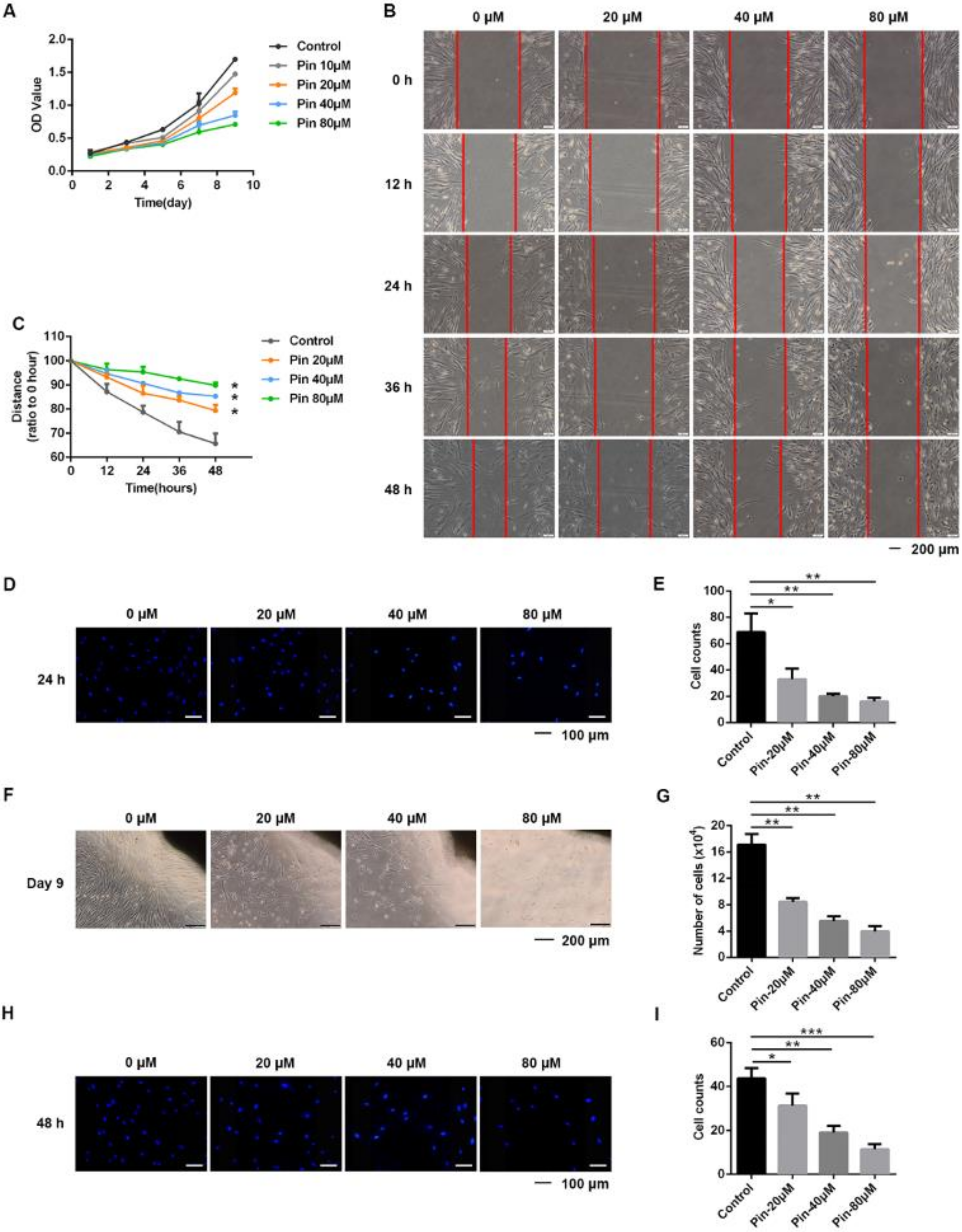
3.1. Pinocembrin Inhibits TGF-β1-Induced Proliferation and Migration of Mouse Primary Dermal Fibroblasts
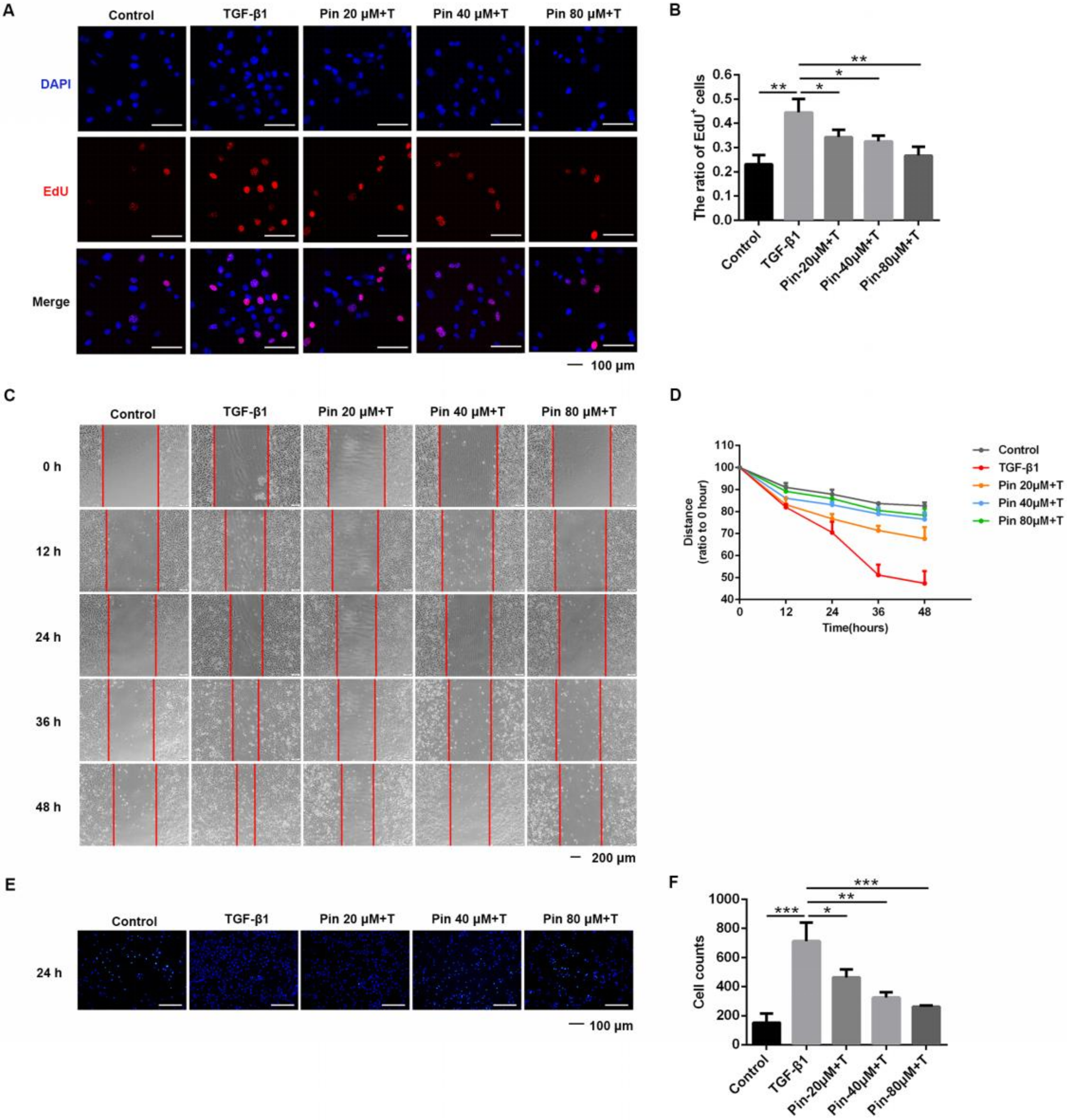
3.2. Pinocembrin Suppresses the Proliferation, Migration, and Invasion of Keloid Fibroblasts
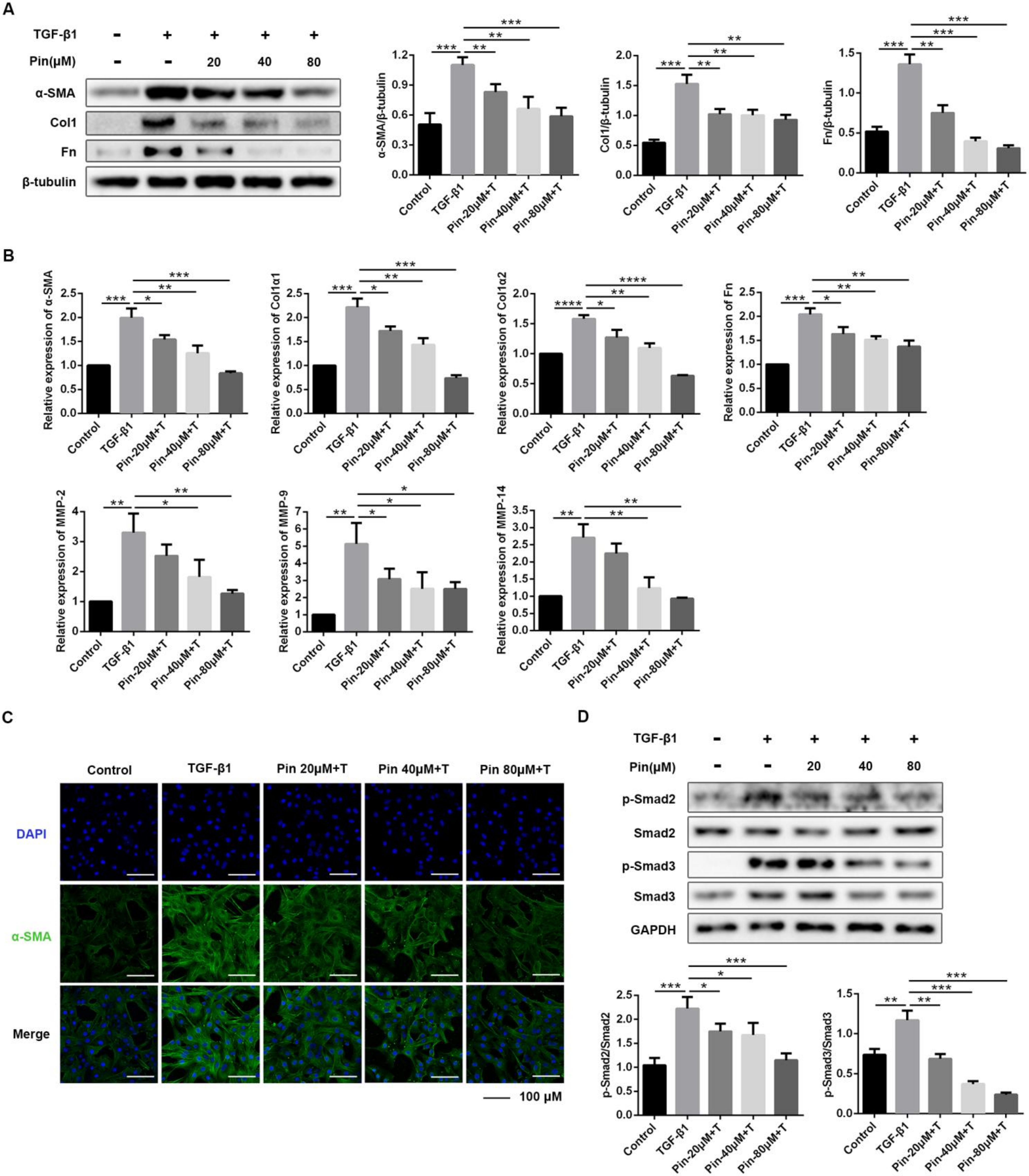
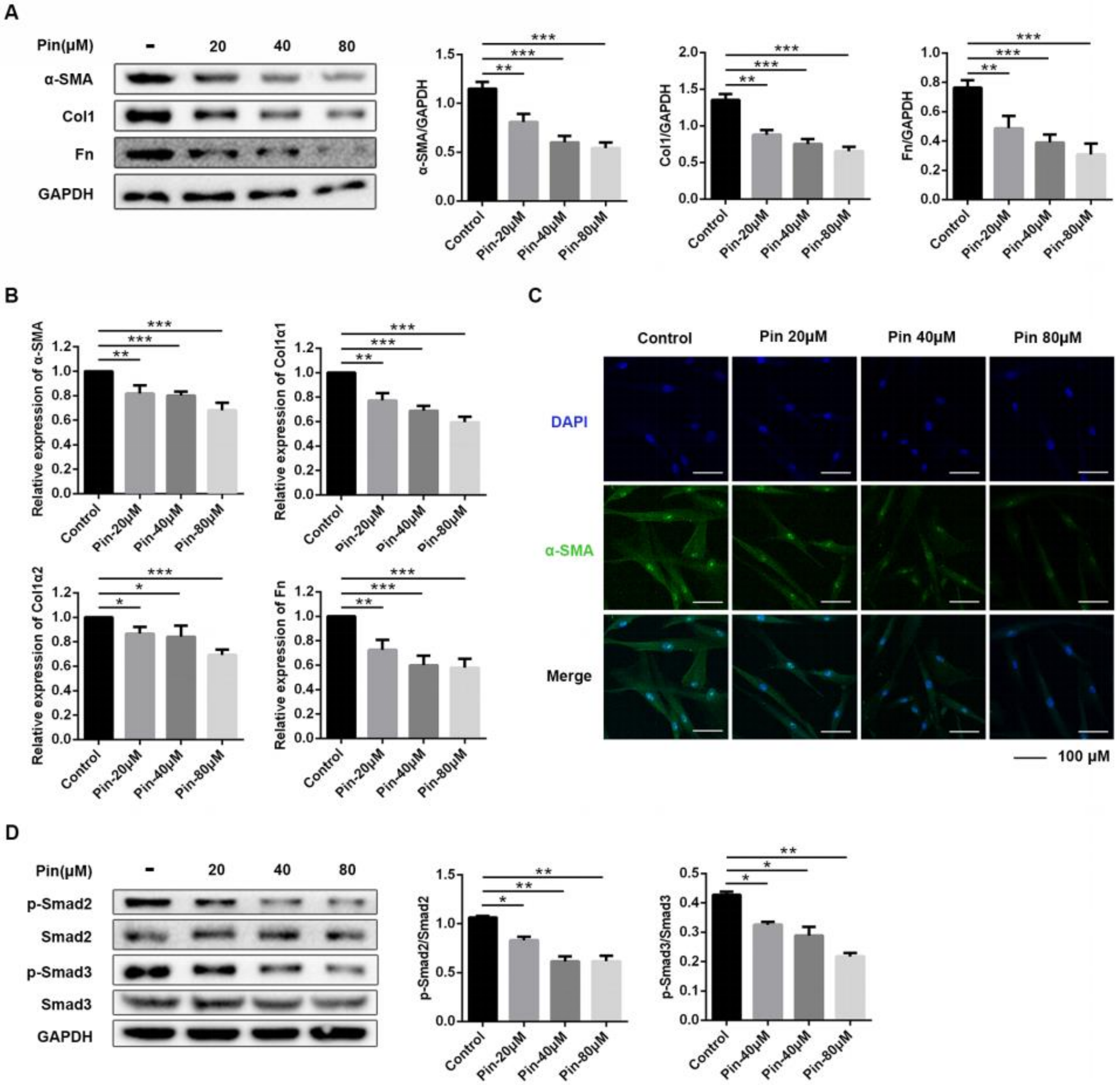
3.3. Pinocembrin Attenuates TGF-β1-Induced Activation of Mouse Primary Dermal Fibroblasts
3.4. Pinocembrin Reduces Fibrosis-Associated Expression of Keloid Pathogenesis in Keloid Fibroblasts
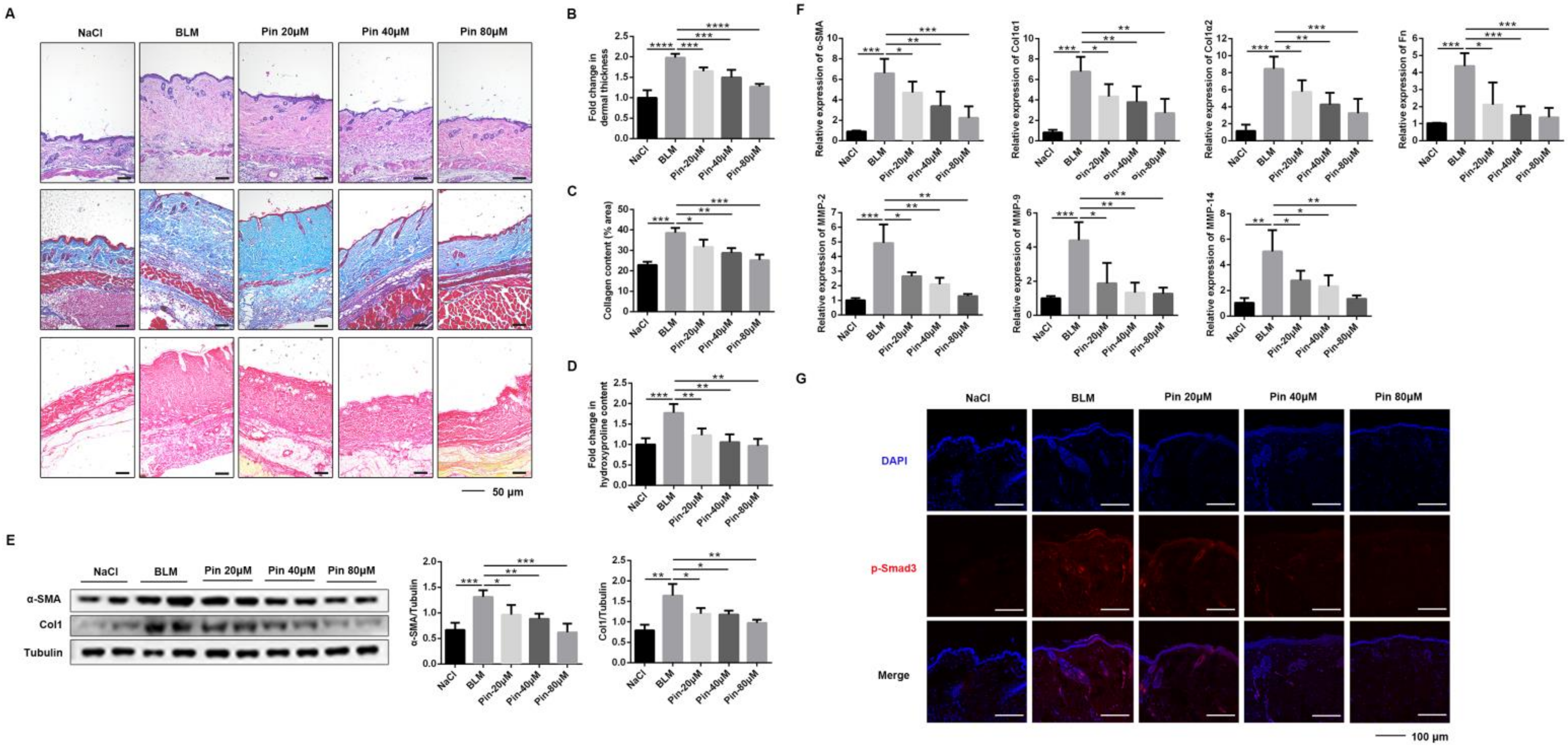
3.5. Pinocembrin Alleviates BLM-Induced Skin Fibrosis in Mice and Inhibits Fibrogenic Activation In Vivo
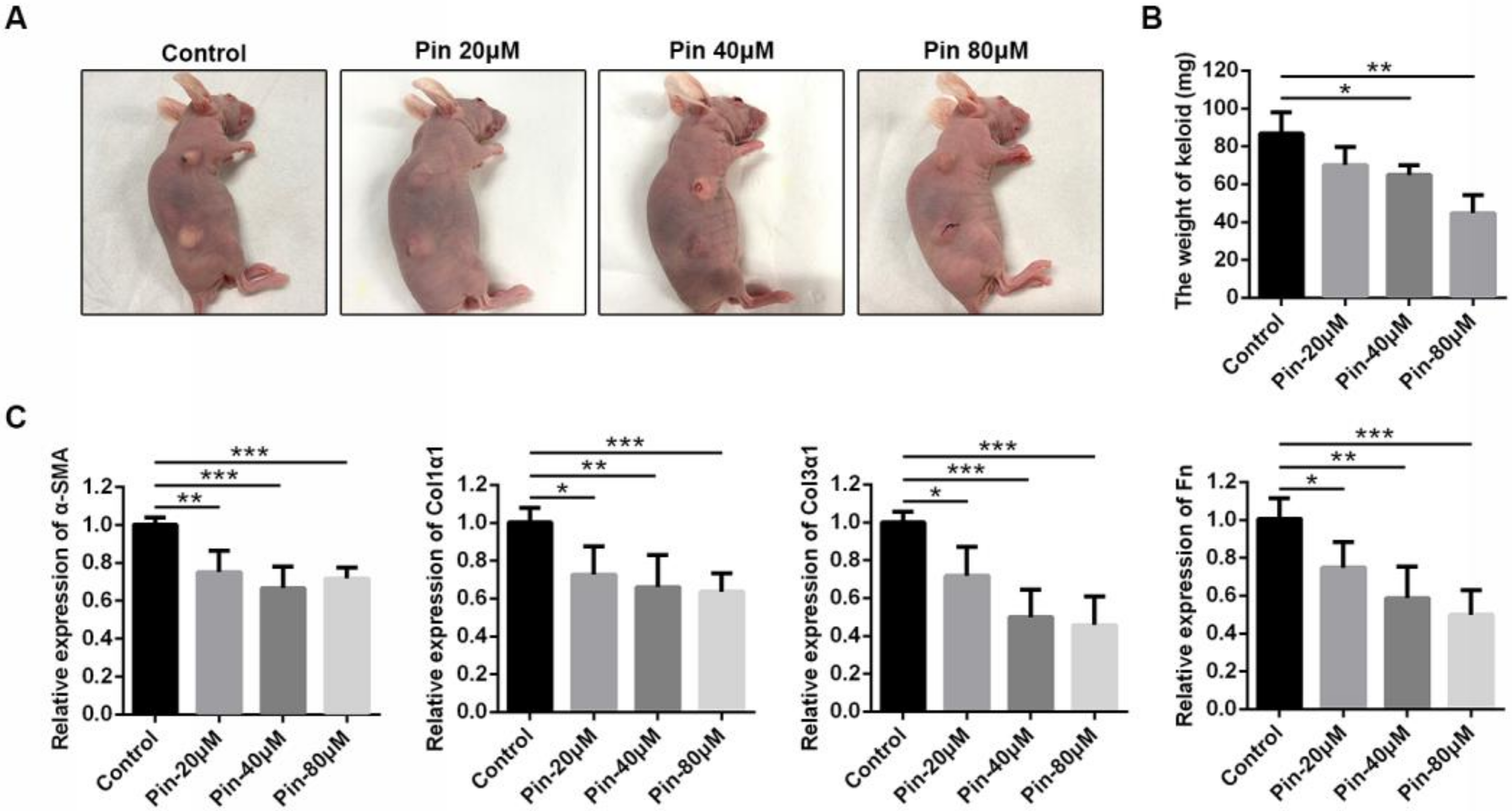
3.6. Intralesional Injection of Pinocembrin Reduces the Size and ECM Gene Expression of Xenografted Keloid Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abignano, G.; del Galdo, F. Quantitating skin fibrosis: Innovative strategies and their clinical implications. Curr. Rheumatol. Rep. 2014, 16, 404. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.Y.; Lagares, D.; Tager, A.M.; Kapoor, M. Fibrosis—A lethal component of systemic sclerosis. Nat. Rev. Rheumatol. 2014, 10, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Petit, A. History of keloid. Ann. Dermatol. Venereol. 2016, 143, 81–95. [Google Scholar] [CrossRef]
- Andrews, J.P.; Marttala, J.; Macarak, E.; Rosenbloom, J.; Uitto, J. Keloids: The paradigm of skin fibrosis—Pathomechanisms and treatment. Matrix Biol. 2016, 51, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Babalola, O.; Mamalis, A.; Lev-Tov, H.; Jagdeo, J. NADPH oxidase enzymes in skin fibrosis: Molecular targets and therapeutic agents. Arch. Dermatol. Res. 2014, 306, 313–330. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Sarrazy, V.; Billet, F.; Micallef, L.; Coulomb, B.; Desmoulière, A. Mechanisms of pathological scarring: Role of myofibroblasts and current developments. Wound Repair Regen. 2011, 19 (Suppl. 1), s10–s15. [Google Scholar] [CrossRef]
- Chalmers, R.L. The evidence for the role of transforming growth factor-beta in the formation of abnormal scarring. Int. Wound J. 2011, 8, 218–223. [Google Scholar] [CrossRef]
- Penn, J.W.; Grobbelaar, A.O.; Rolfe, K.J. The role of the TGF-beta family in wound healing, burns and scarring: A review. Int. J. Burn. Trauma 2012, 2, 18–28. [Google Scholar]
- Unahabhokha, T.; Sucontphunt, A.; Nimmannit, U.; Chanvorachote, P.; Yongsanguanchai, N.; Pongrakhananon, V. Molecular signalings in keloid disease and current therapeutic approaches from natural based compounds. Pharm. Biol. 2015, 53, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.J.; Han, G.; Owens, P.; Siddiqui, Y.; Li, A.G. Role of TGF beta-mediated inflammation in cutaneous wound healing. J. Investig. Dermatol. Symp. Proc. 2006, 11, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.C.; Boone, B.E.; Opalenik, S.R.; Williams, S.M.; Russell, S.B. Gene profiling of keloid fibroblasts shows altered expression in multiple fibrosis-associated pathways. J. Investig. Dermatol. 2008, 128, 1298–1310. [Google Scholar] [CrossRef] [Green Version]
- Pakyari, M.; Farrokhi, A.; Maharlooei, M.K.; Ghahary, A. Critical Role of Transforming Growth Factor Beta in Different Phases of Wound Healing. Adv. Wound Care 2013, 2, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Hanieh, H.; Islam, V.I.H.; Saravanan, S.; Chellappandian, M.; Ragul, K.; Durga, A.; Venugopal, K.; Senthilkumar, V.; Senthilkumar, P. Pinocembrin, a novel histidine decarboxylase inhibitor with anti-allergic potential in in vitro. Eur. J. Pharmacol. 2017, 814, 178–186. [Google Scholar] [CrossRef]
- Saad, M.A.; Salam, R.M.A.; Kenawy, S.A.; Attia, A.S. Pinocembrin attenuates hippocampal inflammation, oxidative perturbations and apoptosis in a rat model of global cerebral ischemia reperfusion. Pharmacol. Rep. 2015, 67, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Said, M.M.; Azab, S.S.; Saeed, N.M.; El-Demerdash, E. Antifibrotic Mechanism of Pinocembrin: Impact on Oxidative Stress, Inflammation and TGF-beta/Smad Inhibition in Rats. Ann. Hepatol. 2018, 17, 307–317. [Google Scholar] [CrossRef]
- Zhou, B.Y.; Wang, W.B.; Wu, X.L.; Zhang, W.J.; Zhou, G.D.; Gao, Z.; Liu, W. Nintedanib inhibits keloid fibroblast functions by blocking the phosphorylation of multiple kinases and enhancing receptor internalization. Acta Pharmacol. Sin. 2020, 41, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, Y.; Jiang, D.; Dong, Y.; Liu, Y.; Zhang, S.; Guo, J.; Qi, C.; Zhao, C.; Jiang, F.; et al. Targeting FSTL1 for Multiple Fibrotic and Systemic Autoimmune Diseases. Mol. Ther. 2021, 29, 347–364. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Darmawan, C.C.; Montenegro, S.E.; Jo, G.; Kusumaningrum, N.; Lee, S.H.; Chung, J.H.; Mun, J.H. Adiponectin-Based Peptide (ADP355) Inhibits Transforming Growth Factor-beta1-Induced Fibrosis in Keloids. Int. J. Mol. Sci. 2020, 21, 2833. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Wang, A.; Li, D.; Wang, Y.; Lin, L. Pinocembrin from Penthorum chinense Pursh suppresses hepatic stellate cells activation through a unified SIRT3-TGF-beta-Smad signaling pathway. Toxicol. Appl. Pharmacol. 2018, 341, 38–50. [Google Scholar] [CrossRef]
- Gao, J.; Lin, S.; Gao, Y.; Zou, X.; Zhu, J.; Chen, M.; Wan, H.; Zhu, H. Pinocembrin inhibits the proliferation and migration and promotes the apoptosis of ovarian cancer cells through down-regulating the mRNA levels of N-cadherin and GABAB receptor. Biomed. Pharmacother. 2019, 120, 109505. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [Green Version]
- Al-Attar, A.; Mess, S.; Thomassen, J.M.; Kauffman, C.L.; Davison, S.P. Keloid pathogenesis and treatment. Plast. Reconstr. Surg. 2006, 117, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, X.F.; Wang, Z.C.; Lou, D.; Fang, Q.Q.; Hu, Y.Y.; Zhao, W.Y.; Zhang, L.Y.; Wu, L.H.; Tan, W.Q. Current potential therapeutic strategies targeting the TGF-beta/Smad signaling pathway to attenuate keloid and hypertrophic scar formation. Biomed. Pharmacother. 2020, 129, 110287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Zhou, S.; Xie, Z.; Wang, C.; Gao, Y.; Zhou, J.; Zhang, X.; Li, Q. Flavones hydroxylated at 5, 7, 3’ and 4’ ameliorate skin fibrosis via inhibiting activin receptor-like kinase 5 kinase activity. Cell Death Dis. 2019, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.Y.; Li, S.R.; Wu, J.L.; Chen, Y.Q.; Li, G.; Bi, S.; Dai, X. Establishment of a hypertrophic scar model by transplanting full-thickness human skin grafts onto the backs of nude mice. Plast. Reconstr. Surg. 2007, 119, 104–109. [Google Scholar] [CrossRef]
- Marttala, J.; Andrews, J.P.; Rosenbloom, J.; Uitto, J. Keloids: Animal models and pathologic equivalents to study tissue fibrosis. Matrix Biol. 2016, 51, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.F.; Lee, J.Y.; Jung, S.N. Models of abnormal scarring. Biomed. Res. Int. 2013, 2013, 423147. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′–3′) |
|---|---|---|
| M-GAPDH | Forward | TGGATTTGGACGCATTGGTC |
| Reverse | TTTGCACTGGTACGTGTTGAT | |
| M-α-SMA | Forward | GCTGGTGATGATGCTCCCA |
| Reverse | GCCCATTCCAACCATTACTCC | |
| M-Col1α1 | Forward | CCAAGAAGACATCCCTGAAGTCA |
| Reverse | TGCACGTCATCGCACACA | |
| M-Col1α2 | Forward | GCAGGTTCACCTACTCTGTCCT |
| Reverse | CTTGCCCCATTCATTTGTCT | |
| M-Fn | Forward | AAGGATGGAGTGATAGCAACCC |
| Reverse | TCTGCTTGAAATCTGGTGTGC | |
| M-MMP-2 | Forward | CAAGTTCCCCGGCGATGTC |
| Reverse | TTCTGGTCAAGGTCACCTGTC | |
| M-MMP-9 | Forward | CTGGACAGCCAGACACTAAAG |
| Reverse | CTCGCGGCAAGTCTTCAGAG | |
| M-MMP-14 | Forward | CAGTATGGCTACCTACCTCCAG |
| Reverse | GCCTTGCCTGTCACTTGTAAA | |
| H-β-actin | Forward | AGGCCAACCGTGAAAAGATG |
| Reverse | AGAGCATAGCCCTCGTAGATGG | |
| H-α-SMA | Forward | TGGGTGAACTCCATCGCTGTA |
| Reverse | GTCGAATGCAACAAGGAAGCC | |
| H-Col1α1 | Forward | AAGCCGGAGGACAACCTTTTA |
| Reverse | GCGAAGAGAATGACCAGATCC | |
| H-Col1α2 | Forward | GATGTTGAACTTGTTGCTGAGG |
| Reverse | TCTTTCCCCATTCATTTGTCTT | |
| H-Col3α1 | Forward | TGGTGTTGGAGCCGCTGCCA |
| Reverse | CTCAGCACTAGAATCTGTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhai, Y.; Xi, B.; Ma, W.; Zhang, J.; Ma, X.; Miao, Y.; Zhao, Y.; Ning, W.; Zhou, H.; et al. Pinocembrin Ameliorates Skin Fibrosis via Inhibiting TGF-β1 Signaling Pathway. Biomolecules 2021, 11, 1240. https://doi.org/10.3390/biom11081240
Li X, Zhai Y, Xi B, Ma W, Zhang J, Ma X, Miao Y, Zhao Y, Ning W, Zhou H, et al. Pinocembrin Ameliorates Skin Fibrosis via Inhibiting TGF-β1 Signaling Pathway. Biomolecules. 2021; 11(8):1240. https://doi.org/10.3390/biom11081240
Chicago/Turabian StyleLi, Xiaohe, Yunqian Zhai, Buri Xi, Wei Ma, Jianwei Zhang, Xiaoyang Ma, Yang Miao, Yongjian Zhao, Wen Ning, Honggang Zhou, and et al. 2021. "Pinocembrin Ameliorates Skin Fibrosis via Inhibiting TGF-β1 Signaling Pathway" Biomolecules 11, no. 8: 1240. https://doi.org/10.3390/biom11081240