
Bioactive Antimicrobial Peptides as Therapeutic Agents for Infected Diabetic Foot Ulcers



Abstract
:1. Introduction
2. Diabetic Foot Infection
3. Antimicrobial Peptides
4. Changes of Endogenous AMPs in DFUs
5. Endogenous and Synthetic AMPs as Promising Therapeutic Agents for Infected Wounds
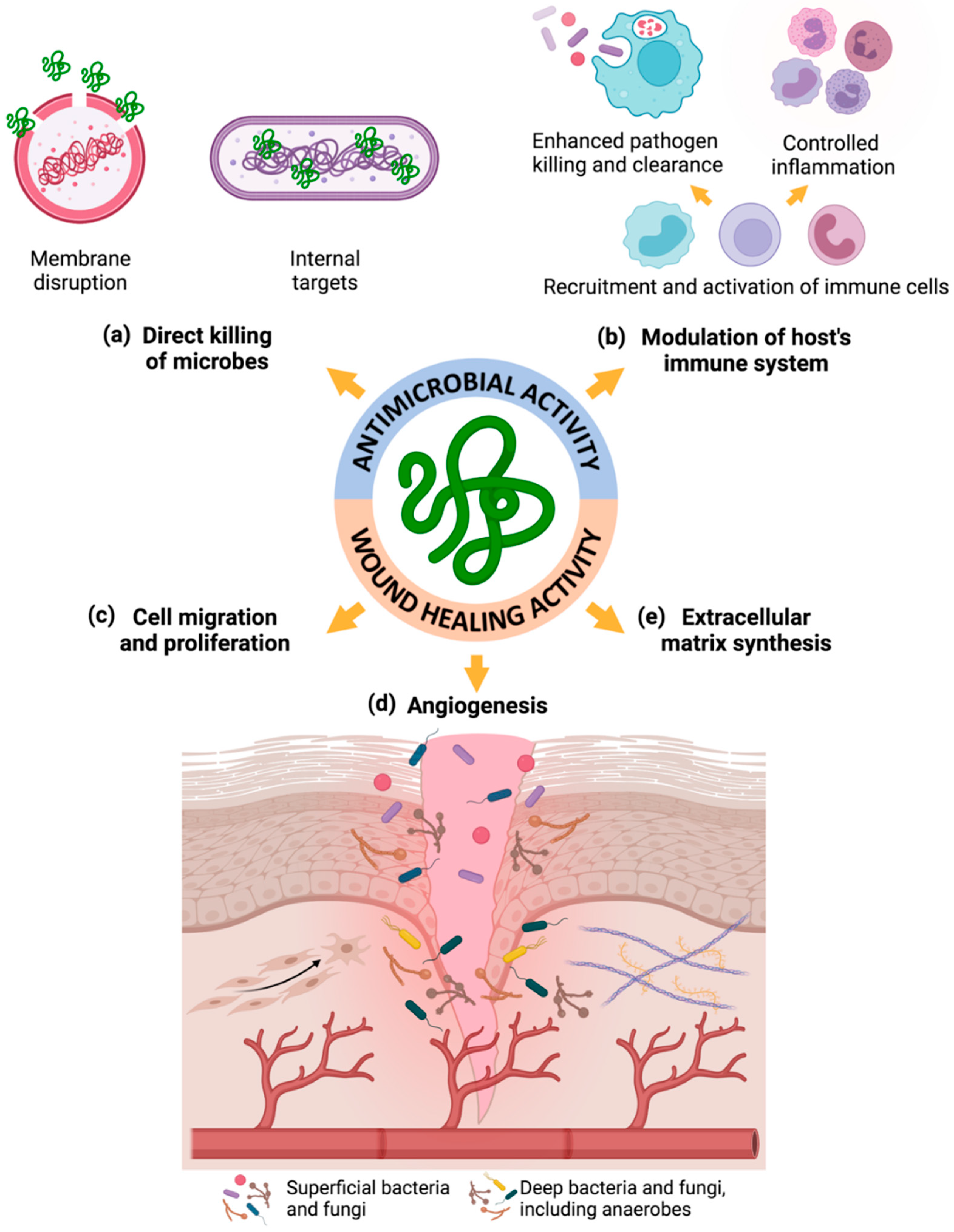
5.1. Free AMPs
5.2. Loaded AMPs on Delivery Systems
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019; ISBN 978-2-930229-87-4. [Google Scholar]
- Vibha, S.P.; Kulkarni, M.M.; Kirthinath Ballala, A.B.; Kamath, A.; Maiya, G.A. Community based study to assess the prevalence of diabetic foot syndrome and associated risk factors among people with diabetes mellitus. BMC Endocr. Disord. 2018, 18, 43. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Brocco, E.; Ninkovic, S.; Marin, M.; Whisstock, C.; Bruseghin, M.; Boschetti, G.; Viti, R.; Forlini, W.; Volpe, A. Diabetic foot management: Multidisciplinary approach for advanced lesion rescue. J. Cardiovasc. Surg. 2018, 59, 670–684. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A.F. Cutaneous Wound Healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Reardon, R.; Simring, D.; Kim, B.; Mortensen, J.; Williams, D.; Leslie, A. The diabetic foot ulcer. Aust. J. Gen. Pract. 2020, 49, 250–255. [Google Scholar] [CrossRef]
- Apelqvist, J. Diagnostics and treatment of the diabetic foot. Endocrine 2012, 41, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Uivaraseanu, B.; Bungau, S.; Tit, D.M.; Fratila, O.; Rus, M.; Maghiar, T.A.; Maghiar, O.; Pantis, C.; Vesa, C.M.; Zaha, D.C. Clinical, Pathological and Microbiological Evaluation of Diabetic Foot Syndrome. Medicina 2020, 56, 380. [Google Scholar] [CrossRef]
- Ramirez-Acuña, J.M.; Cardenas-Cadena, S.A.; Marquez-Salas, P.A.; Garza-Veloz, I.; Perez-Favila, A.; Cid-Baez, M.A.; Flores-Morales, V.; Martinez-Fierro, M.L. Diabetic foot ulcers: Current advances in antimicrobial therapies and emerging treatments. Antibiotics 2019, 8, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalan, L.R.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Uberoi, A.; Gardner, S.E.; Grice, E.A. Strain- and Species-Level Variation in the Microbiome of Diabetic Wounds Is Associated with Clinical Outcomes and Therapeutic Efficacy. Cell Host Microbe 2019, 25, 641–655. [Google Scholar] [CrossRef]
- Kareliya, H.; Bichile, L.; Bal, A.; Varaiya, A.; Bhalekar, P. Fungal Infection in Diabetic Foot a Clinicomicrobiological Study. Acta Sci. Mcrobiology 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Kalshetti, V.T.; Wadile, R.; Bothikar, S.; Ambade, V.; Bhate, V. Study of fungal infections in diabetic foot Ulcer. Indian J Microbiol Res 2017, 4, 87–89. [Google Scholar] [CrossRef]
- Raiesi, O.; Shabandoust, H.; Dehghan, P.; Shamsaei, S.; Soleimani, A. Fungal infection in foot diabetic patients. J. Basic Res. Med. Sci. 2018, 5, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Chellan, G.; Shivaprakash, S.; Ramaiyar, S.K.; Varma, A.K.; Varma, N.; Sukumaran, M.T.; Vasukutty, J.R.; Bal, A.; Kumar, H. Spectrum and prevalence of fungi infecting deep tissues of lower-limb wounds in patients with type 2 diabetes. J. Clin. Microbiol. 2010, 48, 2097–2102. [Google Scholar] [CrossRef] [Green Version]
- Barwell, N.D.; Devers, M.C.; Kennon, B.; Hopkinson, H.E.; McDougall, C.; Young, M.J.; Robertson, H.M.A.; Stang, D.; Dancer, S.J.; Seaton, A.; et al. Diabetic foot infection: Antibiotic therapy and good practice recommendations. Int. J. Clin. Pract. 2017, 71, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobizal, K.B.; Wukich, D.K. Diabetic foot infections: Current concept review. Diabet. Foot Ankle 2012, 3, 18409. [Google Scholar] [CrossRef]
- Percival, S.L.; Malone, M.; Mayer, D.; Salisbury, A.M.; Schultz, G. Role of anaerobes in polymicrobial communities and biofilms complicating diabetic foot ulcers. Int. Wound J. 2018, 15, 776–782. [Google Scholar] [CrossRef]
- MacDonald, A.; Brodell, J.D.; Daiss, J.L.; Schwarz, E.M.; Oh, I. Evidence of differential microbiomes in healing versus non-healing diabetic foot ulcers prior to and following foot salvage therapy. J. Orthop. Res. 2019, 37, 1596–1603. [Google Scholar] [CrossRef]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the chronic-wound microbiome: Fungal communities are prevalent, dynamic, and associated with delayed healing. MBio 2016, 7, e01058-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalan, L.; Grice, E.A. Fungi in the wound microbiome. Adv. Wound Care 2018, 7, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Korting, H.C.; Schlölmann, C.; Stauss-Grabo, M.; Schäfer-Korting, M. Antimicrobial peptides and skin: A paradigm of translational medicine. Skin Pharmacol. Physiol. 2012, 25, 323–334. [Google Scholar] [CrossRef] [Green Version]
- da Costa, J.P.; Cova, M.; Ferreira, R.; Vitorino, R. Antimicrobial peptides: An alternative for innovative medicines? Appl. Microbiol. Biotechnol. 2015, 99, 2023–2040. [Google Scholar] [CrossRef]
- Gonzalez-Curiel, I.; Trujillo, V.; Montoya-Rosales, A.; Rincon, K.; Rivas-Calderon, B.; De Haro-Acosta, J.; Marin-Luevano, P.; Lozano-Lopez, D.; Enciso-Moreno, J.A.; Rivas-Santiago, B. 1,25-dihydroxyvitamin D3 induces LL-37 and HBD-2 production in keratinocytes from diabetic foot ulcers promoting wound healing: An in vitro model. PLoS ONE 2014, 9, e111355. [Google Scholar] [CrossRef]
- Meng, L.; Guo, X.; Yang, X.; Liu, H.; Yu, M.; Wu, Y.; Zhu, Z. Human α defensins promote the expression of the inflammatory cytokine interleukin-8 under high-glucose conditions: Novel insights into the poor healing of diabetic foot ulcers. J. Biochem. Mol. Toxicol. 2019, 33, e22351. [Google Scholar] [CrossRef]
- Santos, R.; Gomes, D.; Macedo, H.; Barros, D.; Tibério, C.; Veiga, A.S.; Tavares, L.; Castanho, M.; Oliveira, M. Guar gum as a new antimicrobial peptide delivery system against diabetic foot ulcers Staphylococcus aureus isolates. J. Med. Microbiol. 2016, 65, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Mi, B.; Liu, J.; Liu, Y.; Hu, L.; Liu, Y.; Panayi, A.C.; Zhou, W.; Liu, G. The designer antimicrobial peptide A-hBD-2 facilitates skin wound healing by stimulating keratinocyte migration and proliferation. Cell. Physiol. Biochem. 2018, 51, 647–663. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound healing: A cellular perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Childs, D.R.; Murthy, A.S. Overview of Wound Healing and Management. Surg. Clin. North Am. 2017, 97, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Petkovic, M.; Sørensen, A.E.; Leal, E.C.; Carvalho, E.; Dalgaard, L.T. Mechanistic Actions of microRNAs in Diabetic Wound Healing. Cells 2020, 9, 2228. [Google Scholar] [CrossRef]
- Petkovic, M.; Vangmouritzen, M.; Mojsoska, B.; Jenssen, H. Immunomodulatory properties of host defence peptides in skin wound healing. Biomolecules 2021, 11, 952. [Google Scholar] [CrossRef]
- Hurlow, J.J.; Humphreys, G.J.; Bowling, F.L.; McBain, A.J. Diabetic foot infection: A critical complication. Int. Wound J. 2018, 15, 814–821. [Google Scholar] [CrossRef]
- Pouget, C.; Dunyach-Remy, C.; Pantel, A.; Schuldiner, S.; Sotto, A.; Lavigne, J.P. Biofilms in diabetic foot ulcers: Significance and clinical relevance. Microorganisms 2020, 8, 1580. [Google Scholar] [CrossRef]
- Han, S.H.; Lee, J.S.; Song, K.H.; Choe, Y.B.; Ahn, K.J.; Lee, Y.W. Differences in foot skin microbiomes between patients with type 2 diabetes and healthy individuals. Mycoses 2020, 63, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.G.; Moura, J.; Carvalho, E.; Empadinhas, N. Microbiota of chronic diabetic wounds: Ecology, impact, and potential for innovative treatment strategies. Front. Microbiol. 2017, 8, 1791. [Google Scholar] [CrossRef]
- Mottola, C.; Mendes, J.J.; Cristino, J.M.; Cavaco-Silva, P.; Tavares, L.; Oliveira, M. Polymicrobial biofilms by diabetic foot clinical isolates. Folia Microbiol. 2016, 61, 35–43. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, A.S. Bad “Staph” in the Wound Environment of Diabetic Foot Ulcers. Cell Host Microbe 2019, 25, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Dörr, S.; Freier, F.; Schlecht, M.; Lobmann, R. Bacterial diversity and inflammatory response at first-time visit in younger and older individuals with diabetic foot infection (DFI). Acta Diabetol. 2021, 58, 181–189. [Google Scholar] [CrossRef]
- Murali, T.S.; Kavitha, S.; Spoorthi, J.; Bhat, D.V.; Prasad, A.S.B.; Upton, Z.; Ramachandra, L.; Acharya, R.V.; Satyamoorthy, K. Characteristics of microbial drug resistance and its correlates in chronic diabetic foot ulcer infections. J. Med. Microbiol. 2014, 63, 1377–1385. [Google Scholar] [CrossRef]
- Dowd, S.E.; Delton Hanson, J.; Rees, E.; Wolcott, R.D.; Zischau, A.M.; Sun, Y.; White, J.; Smith, D.M.; Kennedy, J.; Jones, C.E. Survey of fungi and yeast in polymicrobial infections in chronic wounds. J. Wound Care 2011, 20, 40–47. [Google Scholar] [CrossRef]
- Mehra, B.K.; Singh, A.K.; Gupta, D.; Narang, R.; Patil, R. A Clinicomicrobiological Study on Incidence of Mycotic Infections in Diabetic Foot Ulcers. Int. J. Sci. Study 2017, 50, 12. [Google Scholar] [CrossRef]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Topical antimicrobial peptide formulations for wound healing: Current developments and future prospects. Acta Biomater. 2020, 103, 52–67. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y.M. Endogenous Antimicrobial Peptides and Skin Infections in Atopic Dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Vatan, A.; Saltoglu, N.; Yemisen, M.; Balkan, I.I.; Surme, S.; Demiray, T.; Mete, B.; Tabak, F. Association between biofilm and multi/extensive drug resistance in diabetic foot infection. Int. J. Clin. Pract. 2018, 72, e13060. [Google Scholar] [CrossRef]
- Duplantier, A.J.; van Hoek, M.L. The human cathelicidin antimicrobial peptide LL-37 as a potential treatment for polymicrobial infected wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Teixeira, C.; Ferraz, R.; Prudencio, C.; Gomes, P. Wound-healing peptides for treatment of chronic diabetic foot ulcers and other infected skin injuries. Molecules 2017, 22, 1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomioka, H.; Nakagami, H.; Tenma, A.; Saito, Y.; Kaga, T.; Kanamori, T.; Tamura, N.; Tomono, K.; Kaneda, Y.; Morishita, R. Novel anti-microbial peptide SR-0379 accelerates wound healing via the PI3 kinase/Akt/mTOR pathway. PLoS ONE 2014, 9, e92597. [Google Scholar] [CrossRef]
- Comune, M.; Rai, A.; Chereddy, K.K.; Pinto, S.; Aday, S.; Ferreira, A.F.; Zonari, A.; Blersch, J.; Cunha, R.; Rodrigues, R.; et al. Antimicrobial peptide-gold nanoscale therapeutic formulation with high skin regenerative potential. J. Control. Release 2017, 262, 58–71. [Google Scholar] [CrossRef]
- Pazgier, M.; Hoover, D.M.; Yang, D.; Lu, W.; Lubkowski, J. Human β-defensins. Cell. Mol. Life Sci. 2006, 63, 1294–1313. [Google Scholar] [CrossRef]
- Rivas-Santiago, B.; Trujillo, V.; Montoya, A.; Gonzalez-Curiel, I.; Castañeda-Delgado, J.; Cardenas, A.; Rincon, K.; Hernandez, M.L.; Hernández-Pando, R. Expression of antimicrobial peptides in diabetic foot ulcer. J. Dermatol. Sci. 2012, 65, 19–26. [Google Scholar] [CrossRef]
- Torrent, M.; Andreu, D.; Nogués, V.M.; Boix, E. Connecting peptide physicochemical and antimicrobial properties by a rational prediction model. PLoS ONE 2011, 6, e16968. [Google Scholar] [CrossRef]
- NIHR Horizon Scanning Research and Intelligence Centre. Pexiganan (Locilex) for Mild Foot Infection in Diabetic Foot Ulcers, University of Birmingham: Birmingham, UK, 2015.
- Zhang, M.; Zhao, J.; Zheng, J. Molecular understanding of a potential functional link between antimicrobial and amyloid peptides. Soft Matter 2014, 10, 7425–7451. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Antimicrobial peptides activity in the skin. Ski. Res. Technol. 2019, 25, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönberg, A.; Mahlapuu, M.; Ståhle, M.; Whately-Smith, C.; Rollman, O. Treatment with LL-37 is safe and effective in enhancing healing of hard-to-heal venous leg ulcers: A randomized, placebo-controlled clinical trial. Wound Repair Regen. 2014, 22, 613–621. [Google Scholar] [CrossRef]
- Grek, C.L.; Prasad, G.M.; Viswanathan, V.; Armstrong, D.G.; Gourdie, R.G.; Ghatnekar, G.S. Topical administration of a connexin43-based peptide augments healing of chronic neuropathic diabetic foot ulcers: A multicenter, randomized trial. Wound Repair Regen. 2015, 23, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Marin-Luevano, P.; Trujillo, V.; Rodriguez-Carlos, A.; González-Curiel, I.; Enciso-Moreno, J.A.; Hancock, R.E.W.; Rivas-Santiago, B. Induction by innate defence regulator peptide 1018 of pro-angiogenic molecules and endothelial cell migration in a high glucose environment. Peptides 2018, 101, 135–144. [Google Scholar] [CrossRef]
- Flamm, R.K.; Rhomberg, P.R.; Simpson, K.M.; Farrell, D.J.; Sader, H.S.; Jones, R.N. In vitro spectrum of pexiganan activity when tested against pathogens from diabetic foot infections and with selected resistance mechanisms. Antimicrob. Agents Chemother. 2015, 59, 1751–1754. [Google Scholar] [CrossRef] [Green Version]
- Lan, C.C.E.; Wu, C.S.; Huang, S.M.; Kuo, H.Y.; Wu, I.H.; Liang, C.W.; Chen, G.S. High-glucose environment reduces human β-defensin-2 expression in human keratinocytes: Implications for poor diabetic wound healing. Br. J. Dermatol. 2012, 166, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Galkowska, H.; Olszewski, W.L.; Wojewodzka, U. Expression of natural antimicrobial peptide β-defensin-2 and Langerhans cell accumulation in epidermis from human non-healing leg ulcers. Folia Histochem. Cytobiol. 2005, 43, 133–136. [Google Scholar] [CrossRef]
- Liu, H.; Duan, Z.; Tang, J.; Lv, Q.; Rong, M.; Lai, R. A short peptide from frog skin accelerates diabetic wound healing. FEBS J. 2014, 281, 4633–4643. [Google Scholar] [CrossRef] [Green Version]
- Steinstraesser, L.; Hirsch, T.; Schulte, M.; Kueckelhaus, M.; Jacobsen, F.; Mersch, E.A.; Stricker, I.; Afacan, N.; Jenssen, H.; Hancock, R.E.W.; et al. Innate defense regulator peptide 1018 in wound healing and wound infection. PLoS ONE 2012, 7, e39373. [Google Scholar] [CrossRef] [Green Version]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Gomes, A.; Ferraz, R.; Teixeira, C. The Emerging Role of Ionic Liquid-Based Approaches for Enhanced Skin Permeation of Bioactive Molecules: A Snapshot of the Past Couple of Years. Int. J. Mol. Sci. 2021, 22, 11991. [Google Scholar] [CrossRef]
- Li, W.; Separovic, F.; O’Brien-Simpson, N.M.; Wade, J.D. Chemically modified and conjugated antimicrobial peptides against superbugs. Chem. Soc. Rev. 2021, 50, 4932–4973. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Bessa, L.J.; Fernandes, I.; Ferraz, R.; Mateus, N.; Gameiro, P.; Teixeira, C.; Gomes, P. Turning a collagenesis-inducing peptide into a potent antibacterial and antibiofilm agent against multidrug-resistant gram-negative bacteria. Front. Microbiol. 2019, 10, 1915. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.; Bessa, L.J.; Fernandes, I.; Ferraz, R.; Martins, M.C.L.; Mateus, N.; Gameiro, P.; Teixeira, C.; Gomes, P. Disclosure of a Promising Lead to Tackle Complicated Skin and Skin Structure Infections: Antimicrobial and Antibiofilm Actions of Peptide PP4-3. Pharmaceutics 2021, 13, 1962. [Google Scholar] [CrossRef]
- Mouritzen, M.V.; Petkovic, M.; Qvist, K.; Poulsen, S.S.; Alarico, S.; Leal, E.C.; Dalgaard, L.T.; Empadinhas, N.; Carvalho, E.; Jenssen, H. Improved diabetic wound healing by LFcinB is associated with relevant changes in the skin immune response and microbiota. Mol. Ther. Methods Clin. Dev. 2021, 20, 726–739. [Google Scholar] [CrossRef]
- Jung Kim, D.; Lee, Y.W.; Park, M.K.; Shin, J.R.; Lim, K.J.; Cho, J.H.; Kim, S.C. Efficacy of the designer antimicrobial peptide SHAP1 in wound healing and wound infection. Amino Acids 2014, 46, 2333–2343. [Google Scholar] [CrossRef] [PubMed]
- Bolatchiev, A.; Baturin, V.; Bazikov, I.; Maltsev, A.; Kunitsina, E. Effect of antimicrobial peptides HNP-1 and hBD-1 on Staphylococcus aureus strains in vitro and in vivo. Fundam. Clin. Pharmacol. 2020, 34, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Ruza, D.; Cunha, E.; Tavares, L.; Oliveira, M. Diabetic foot infections: Application of a nisin-biogel to complement the activity of conventional antibiotics and antiseptics against Staphylococcus aureus biofilms. PLoS ONE 2019, 14, e0220000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayramov, D.; Li, Z.; Patel, E.; Izadjoo, M.; Kim, H.; Neff, J. A Novel Peptide-Based Antimicrobial Wound Treatment is Effective Against Biofilms of Multi-Drug Resistant Wound Pathogens. Mil. Med. 2018, 183, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Du, X.; Jiang, L.; Luo, H.; Wang, F.; Wang, J.; Qiu, L.; Liu, L.; Liu, X.; Wang, X.; et al. Glucose Oxidase-Loaded Antimicrobial Peptide Hydrogels: Potential Dressings for Diabetic Wound. J. Nanosci. Nanotechnol. 2019, 20, 2087–2094. [Google Scholar] [CrossRef]
- Lipsky, B.A.; Holroyd, K.J.; Zasloff, M. Topical versus systemic antimicrobial therapy for treating mildly infected diabetic foot ulcers: A randomized, controlled, double-blinded, multicenter trial of pexiganan cream. Clin. Infect. Dis. 2008, 47, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, D.W.; Kim, S.H.; Kim, H.H.; Lee, K.H.; Ki, C.S.; Park, Y.H. Multi-biofunction of antimicrobial peptide-immobilized silk fibroin nanofiber membrane: Implications for wound healing. Acta Biomater. 2016, 39, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.S.; Vila, T.; Hefni, E.; Karlsson, A.J.; Jabra-Rizk, M.A. Evaluation of the antifungal and wound-healing properties of a novel peptide-based bioadhesive hydrogel formulation. Antimicrob. Agents Chemother. 2019, 63, e00888-19. [Google Scholar] [CrossRef] [PubMed]
- Gawande, P.V.; Leung, K.P.; Madhyastha, S. Antibiofilm and Antimicrobial Efficacy of DispersinB®-KSL-W Peptide-Based Wound Gel Against Chronic Wound Infection Associated Bacteria. Curr. Microbiol. 2014, 68, 635–641. [Google Scholar] [CrossRef]
- Riool, M.; de Breij, A.; Kwakman, P.H.S.; Schonkeren-Ravensbergen, E.; de Boer, L.; Cordfunke, R.A.; Malanovic, N.; Drijfhout, J.W.; Nibbering, P.H.; Zaat, S.A.J. Thrombocidin-1-derived antimicrobial peptide TC19 combats superficial multi-drug resistant bacterial wound infections. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183282. [Google Scholar] [CrossRef]
- Lin, Z.; Wu, T.; Wang, W.; Li, B.; Wang, M.; Chen, L.; Xia, H.; Zhang, T. Biofunctions of antimicrobial peptide-conjugated alginate/hyaluronic acid/collagen wound dressings promote wound healing of a mixed-bacteria-infected wound. Int. J. Biol. Macromol. 2019, 140, 330–342. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gram-Positive | BACTERIA Gram-Negative | Anaerobes |
|---|---|---|
| S. aureus (MSSA and MRSA) [8,9,10,14,15,17,32,34,35,36,37,38] C. striatum [10,32,34,43] Streptococcus β-hemolytic [8,9,17,18,32] | P. aeruginosa [8,9,10,14,17,35,36,37,38,44] Proteus spp. [8,9,17,37] Enterobacter spp. [8,17,37] Citrobacter spp. [8,17,42] E. coli [8,17,37] A. baumannii [8,17,38,43] | Bacteroides spp. [9,17,18] Prevotella spp. [9,10,17] Peptostreptococcus spp. [9] Clostridium spp. [9] |
| FUNGI | ||
| C. albicans [11,12,13,14,19,20,39,40] | C. tropicalis [12,14,20,39,40] | C. glabrata [12,39,40] |
| C. parapsilosis [14,19,20,39,40] | T. rubrum [12,13,44] | T. mentagrophytes [12,13,40] |
| A. fumigatus [12,14,20] | T. asahii [14,19,20] | C. herbarum [19,20,40] |
| AMP | Primary Structure | Length (aa) | PDB Code | Secondary Structure | Tertiary Structure | MW (Da) | pI | Net Charge | Hydrophobicity (kcal/mol) |
|---|---|---|---|---|---|---|---|---|---|
| hBD-1 | DHYNCVSSGGQCLYSACPIFTKIQGTCYRGKAKCCK | 36 | 1IJU | α-helix + β-strand | three antiparallel β-sheets stabilized by three disulfide bridges and flanked by an α-helix segment, together stabilized by a disulfide bridge | 3931.77 | 8.55 | +4 | +28.98 |
| hBD-2 | GIGDPVTCLKSGAICHPVFCPRRYKQIGTCGLPGTKCCKKP | 41 | 1FD4 | 4331.17 | 9.26 | +6 | +32.25 | ||
| hBD-3 | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK | 45 | Not found | 5157.70 | 10.47 | +11 | +45.26 | ||
| LL-37 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | 37 | 2K6O | α-helix | one α-helical conformation | 4490.57 | 11.15 | +6 | +41.03 |
| AMP | Sequence | Source | Delivery Method | Role in Antimicrobial and Wound-Healing Activities | Reference |
|---|---|---|---|---|---|
| hBD-2 LL-37 | GIGDPVTCLKSGAICHPV FCPRRYKQIGTCGLPGTK CCKKP LLGDFFRKSKEKIGKEFK RIVQRIKDFLRNLVPRTES | Endogenous (human) Endogenous (human) | Free Free | ↑ antimicrobial activity (E. coli) ↑ keratinocyte migration | [23] |
| CW49 | APFRMGICTTN | Synthetic (frog skin) | Free | ↑ angiogenic ability ↑ anti-inflammatory effect little effect on re-epithelialization | [61] |
| IDR-1018 | VRLIVAVRIWRR-NH2 | Synthetic | Free | ↓ in vitro toxicity compared to LL-37 ↑ wound healing in S. aureus infected porcine and non-diabetic but not in diabetic murine wounds | [62] |
| IDR-1018 | VRLIVAVRIWRR-NH2 | Synthetic | Free | ↑ angiogenic ability ↑ anti-inflammatory effect ↑ migration of endothelial cells | [57] |
| Pexiganan | GIGKFLKKAKKFGKAFV KILKK | Synthetic (analogue of magainin II—frog skin) | Free | ↑ antimicrobial activity (E. coli, E. cloacae, Citrobacter spp., P. vulgaris, M. morganii, K. pneumoniae, S. marcescens, P. aeruginosa, A. baumannii, S. agalactiae, S. pyogenes, E. faecium, MSSA and MRSA) | [58] |
| 3.1-PP4 | KKLLKWLLKLLKTTKS | Synthetic | Free (chemically modified) | ↓ toxicity to HFF-1 human fibroblasts ↑ antimicrobial activity (E. coli, P. aeruginosa, and K. pneumoniae, including MDR isolates) ↓ formation of K. pneumoniae biofilms | [66] |
| PP4-3.11 | KTTKSKKLLKWLLKLL | Synthetic | Free (chemically modified) | ↑ antimicrobial activity (Gram-positive and Gram-negative bacteria, including MDR isolates, as well as against relevant Candida spp.) | [67] |
| A-hBD-2 | APKAMVTCLKSGAICHP VFCPRRYKQIGTCGLPGT KCCKKP | Synthetic | Free (chemically modified) | ↑ structural stability ↓ toxicity to keratinocytes ↑ antimicrobial activity (S. aureus) ↑ migration and proliferation of keratinocytes ↓ terminal differentiation of keratinocytes ↑ mobilization of intracellular Ca2+ ↑ wound healing in vivo | [26] |
| LFcinB | FKCRRWQWRMKKLGAP SITCVRRAF | Synthetic (derived from bLF) | Free (chemically modified) | ↑ keratinocyte migration in vitro and ex vivo ↑ wound healing ↑ antimicrobial activity (B. pumilus and S. aureus) ↑ angiogenesis and collagen deposition ↓ inflammation | [68] |
| SHAP1 | APKAMKLLKKLLKLQK KGI | Synthetic | Free (chemically modified) | ↓ toxicity to human erythrocytes and keratinocytes ↑ stability to proteases exposure ↑ wound closure compared to LL- 37 in vitro ↑ healing in vivo full-thickness excisional wounds ↑ antimicrobial activity (S. aureus) ↑ healing in S. aureus-infected murine wounds | [69] |
| SR-03791 | MLKLIFLHRLKRMRKRL DLysRK | Synthetic | Free (chemically modified) | ↑ proliferation of human dermal fibroblasts ↑ antimicrobial activity (bacteria, including drug-resistant, and also fungi, namely: E. coli, P. aeruginosa, S. aureus, C. krusei, T. mentagrophytes, T. rubrum, MRSA and A. baumannii (MDR)) ↑ accelerated wound healing in two different wound-healing rat models | [47] |
| AMP | Sequence | Source | Delivery Method | Role in Antimicrobial and Wound-Healing Activities | Reference |
|---|---|---|---|---|---|
| hBD-1 HNP-1 | GNFLTGLGHRSDHYNCV SSGGQCLYSACPIFTKIQ GTCYRGKAKCCK EPLQARADEVAAAPEQIAADIPEVVVSLAWDESL APKHPGSRKNMACYCRI PACIAGERRYGTCIYQGR LWAFCC | Endogenous (human) Endogenous (human) | Niosomal gel Niosomal gel | ↑ antimicrobial activity (MRSA-infected wound in rats and MSSA and MRSA isolated from patients with DFIs) | [70] |
| Nisin | ITSISLCTPGCKTGALMGCNMKTATCH(or N)CSIHVSK | Endogenous (bacteria) | Guar gum gel | ↑ antimicrobial activity against S. aureus DFU biofilm-producing isolates, including some multidrug-resistant clinical isolates | [25] |
| Nisin | ITSISLCTPGCKTGALMGCNMKTATCH(or N)CSIHVSK | Endogenous (bacteria) | Guar gum gel | ↑ antibacterial activity against biofilms formed by DFI S. aureus | [71] |
| aCT1 2 | RQPKIWFPNRRKPWKKRPRPDDLEI-acid | Synthetic (analogue of Cx43) | Hydroxyethyl cellulose gel | ↓ ulcer area in DFU patients ↑ ulcer re-epithelialization in DFU patients ↓ time-to-complete-ulcer closure in DFU patients | [56] |
| ASP-1 ASP-2 | RRWVRRVRRWVRRVVRVVRRWVRR RWWRWWRRWWRR | Synthetic | Gel, Stratex or PU-based dressings | ↑ eradication of mono- and polymicrobial biofilms of MDR pathogens: S. aureus, A. baumannii, K. pneumoniae, P. aeruginosa, and MRSA ↑ BI compared to free ASP-1 and ASP-2 | [72] |
| IKYLSVN | IKYLSVN | Synthetic | GOx-loaded hydrogel | ↑ antimicrobial activity (S. aureus) ↓ blood glucose concentration of diabetic patients | [73] |
| LL-37 | LLGDFFRKSKEKIGKEFK RIVQRIKDFLRNLVPRTE SC | Synthetic | Gold-nanoscale formulation | ↑ phosphorylation of EGFR and ERK1/2 ↑ migratory properties of keratinocytes ↑ wound-healing activity in vivo ↑ expression of collagen, IL6 and VEGF | [48] |
| Pexiganan 2 | GIGKFLKKAKKFGKAFV KILKK | Synthetic (analogue of magainin II—frog skin) | Cream | =clinical outcome, microbiological eradication (S. aureus, E. coli, E. cloacae, S. marcescens, P. aeruginosa, Enterococcus spp., MSSA and MRSA), and wound healing as ofloxacin ↓ bacterial resistance in vivo | [74] |
| Cys-KR12 | CKRIVKRIKKWLR | Synthetic (originated from LL37) | SF nanofiber membrane (chemically modified) | ↑ antimicrobial activity (S. aureus, S. epidermidis, E. coli, and P. aeruginosa) ↑ proliferation of keratinocytes and fibroblasts ↑ differentiation of keratinocytes ↓ LPS-induced TNF-α expression of monocytes | [75] |
| K11R-K17R1 | DSHAKRHHGYRRKFHERHHSHRGY | Synthetic (analogue of Hst-5 peptide) | HPMC-based bioadhesive hydrogel (chemically modified) | ↑ antimicrobial activity (C. albicans strains resistant to traditional antifungals) ↑ cell proliferation and migration in human oral keratinocytes | [76] |
| KSL-W | KKVVFWVKFK | Synthetic (analogue of KSL peptide) | Pluronic F-127 gel (chemically modified) | ↑ antibiofilm and antimicrobial activity (chronic wound infection biofilm-embedded bacteria, including MRSA, S. epidermidis, CoNS, and A. baumannii) | [77] |
| TC19 | LRCMCIKWWSGKHPK | Synthetic (derived from human TC-1-derived peptide L3) | HPMC gel (chemically modified) | ↓ toxicity to human fibroblasts ↑ antimicrobial activity (ESKAPE panel in vitro, and MRSA and A. baumannii (MDR) in a murine superficial wound infection model) ↓ bacterial resistance inflammation in vitro | [78] |
| Tet213 | KRWWKWWRRC | Synthetic (cysteinylated HHC36 peptide) | Alg/HA/Col dressing (chemically modified) | ↑ antimicrobial activity (E. coli, S. aureus, MRSA) ↑ proliferation of NIH 3T3 fibroblast cells ↑ wound healing, re-epithelialization, collagen deposition, and angiogenesis in vivo rat model of partial-thickness mixed-bacterial infected wounds | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Silva, J.; Leal, E.C.; Carvalho, E. Bioactive Antimicrobial Peptides as Therapeutic Agents for Infected Diabetic Foot Ulcers. Biomolecules 2021, 11, 1894. https://doi.org/10.3390/biom11121894
Da Silva J, Leal EC, Carvalho E. Bioactive Antimicrobial Peptides as Therapeutic Agents for Infected Diabetic Foot Ulcers. Biomolecules. 2021; 11(12):1894. https://doi.org/10.3390/biom11121894
Chicago/Turabian StyleDa Silva, Jessica, Ermelindo C. Leal, and Eugénia Carvalho. 2021. "Bioactive Antimicrobial Peptides as Therapeutic Agents for Infected Diabetic Foot Ulcers" Biomolecules 11, no. 12: 1894. https://doi.org/10.3390/biom11121894