
Multiple Metabolic Engineering Strategies to Improve Shikimate Titer in Escherichia coli
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Reagents
2.2. Plasmid Construction
2.3. CRISPR/Cas9-Mediated Genome Editing
2.4. Shake Flask Fermentation
2.5. Fed-Batch Fermentation in a 5 L Fermenter
2.6. Analytical Methods
2.7. Statistical Analysis
3. Results and Discussion
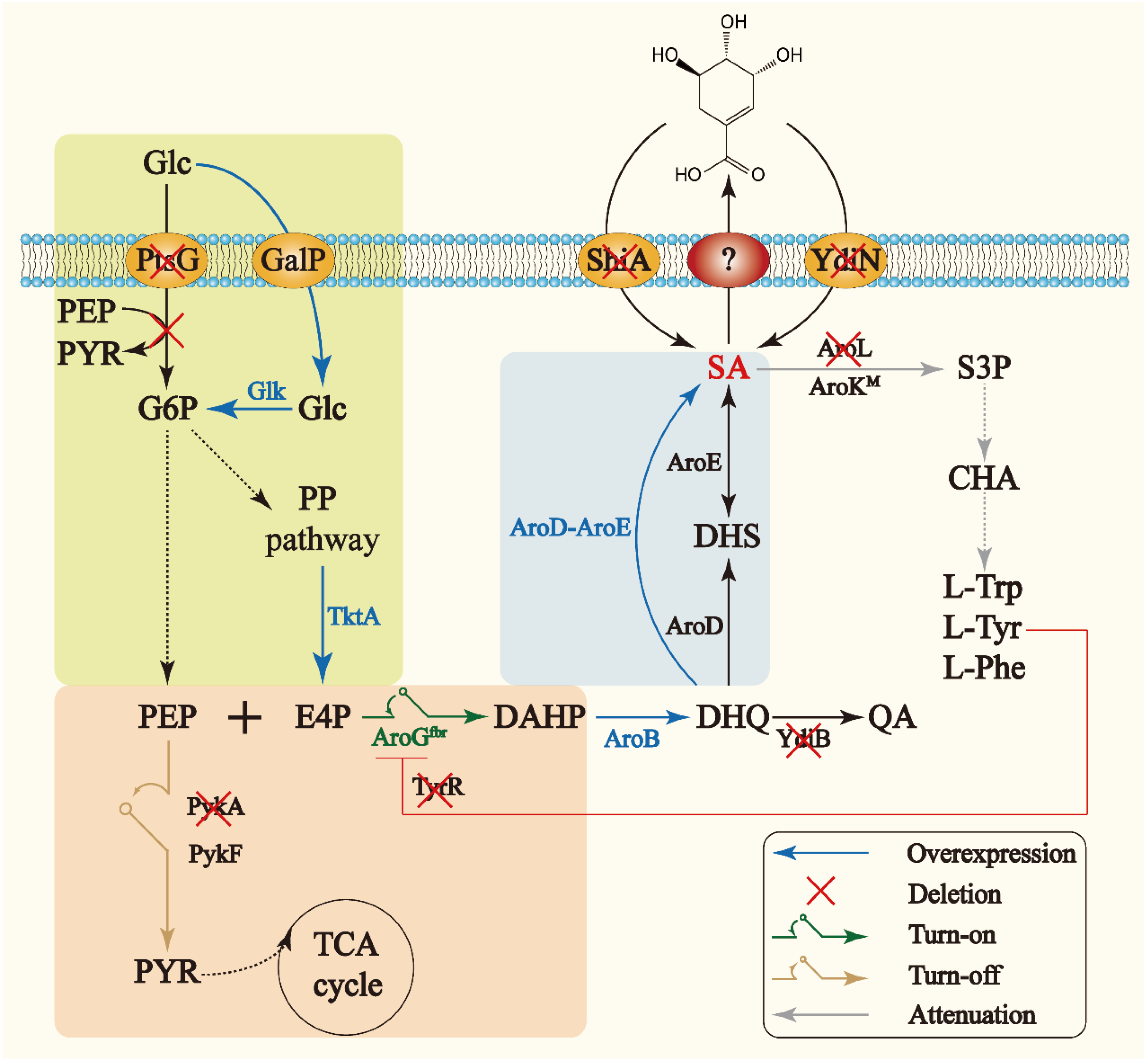
3.1. Strategies for Construction of a Basal Shikimate Production Strain
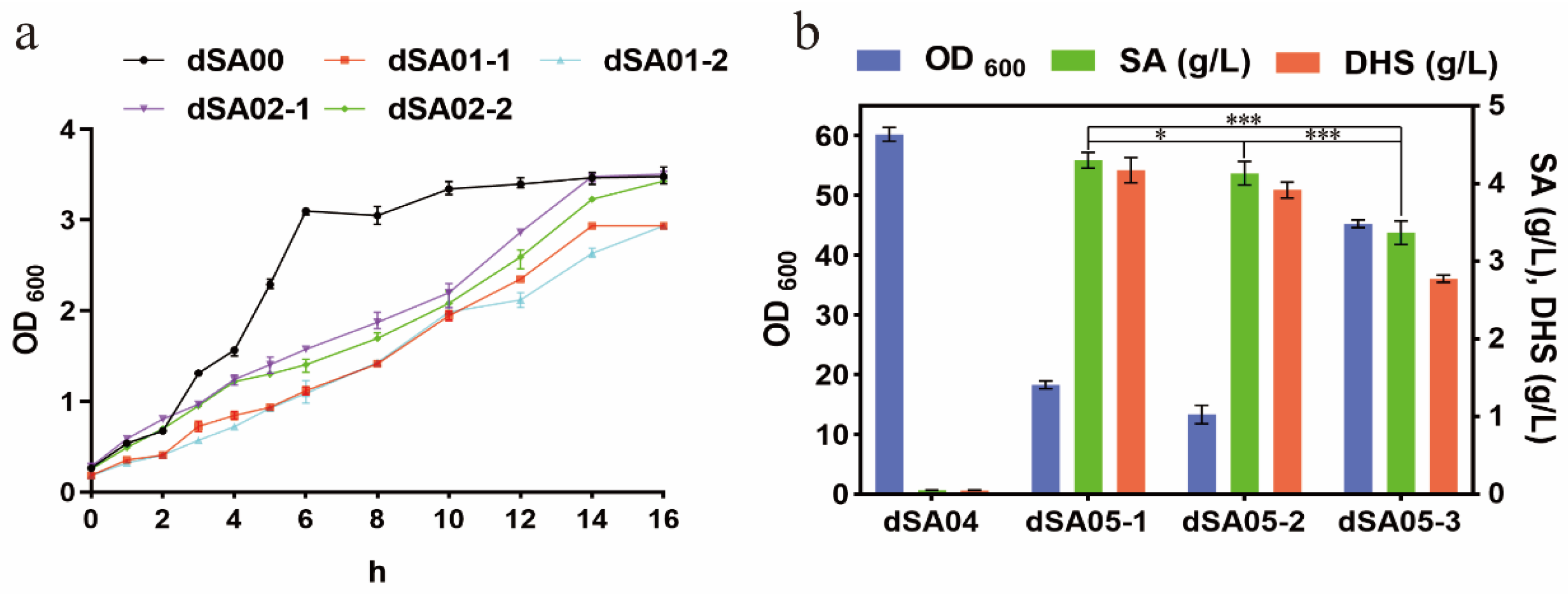
3.1.1. Modification of Glucose Uptake Pathway
3.1.2. Introduction of Feedback-Resistant Mutant aroGfbr
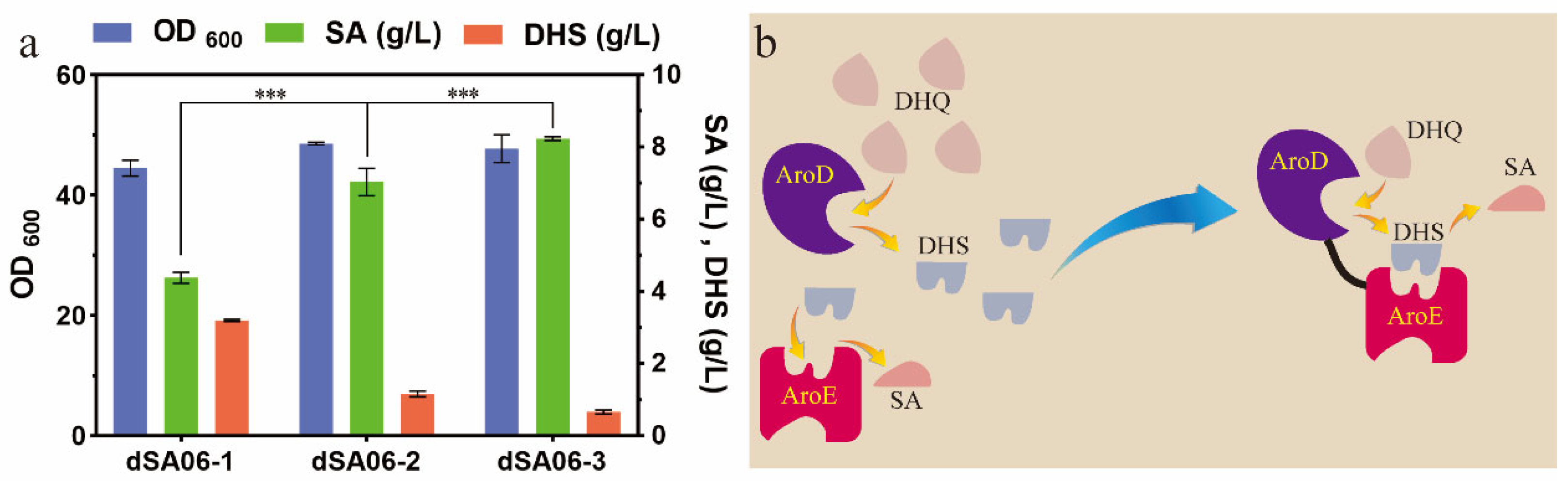
3.2. Strategy for Enhancing Shikimate Production and Reducing DHS Accumulation
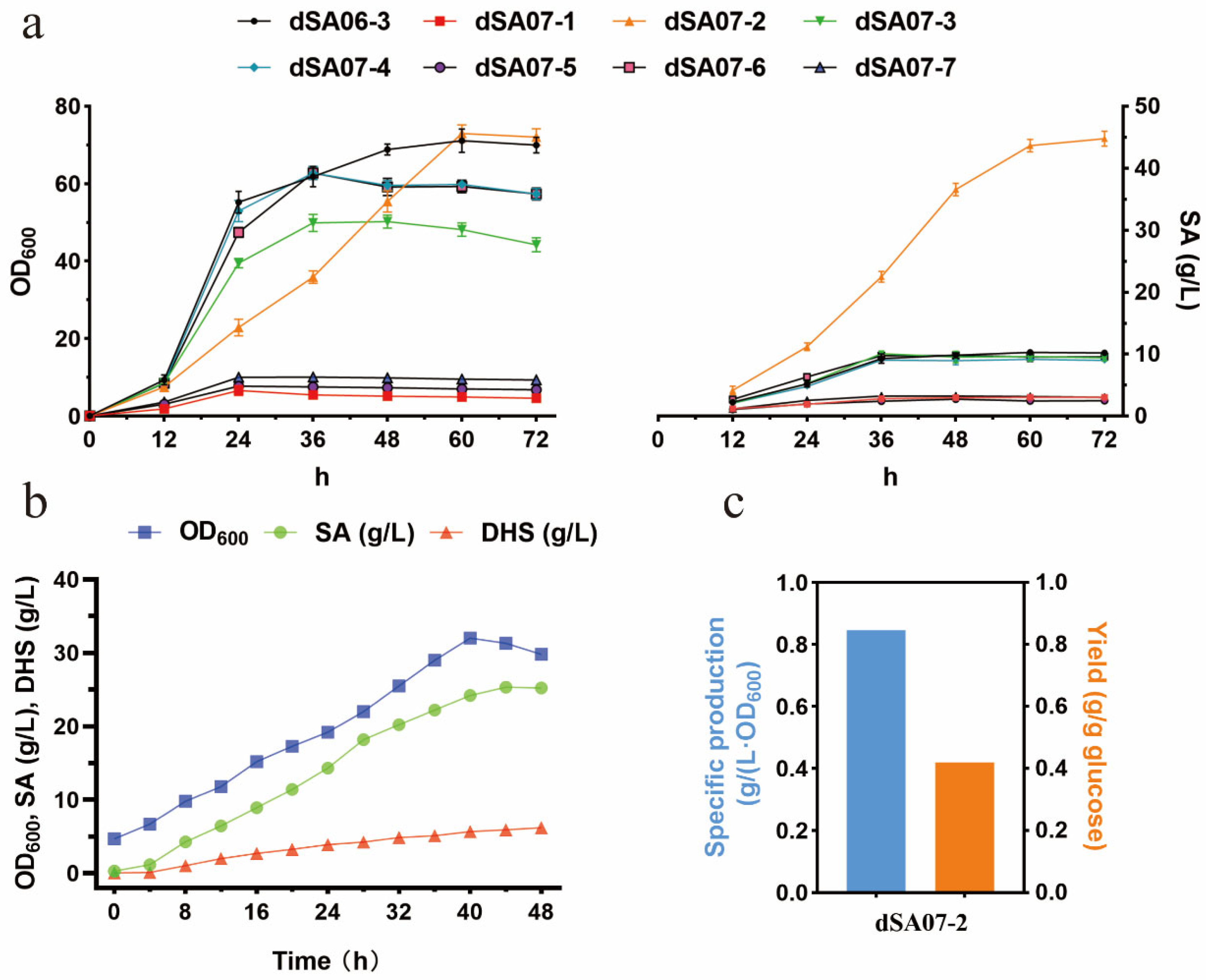
3.3. Strategy for Attenuation of aroK Expression
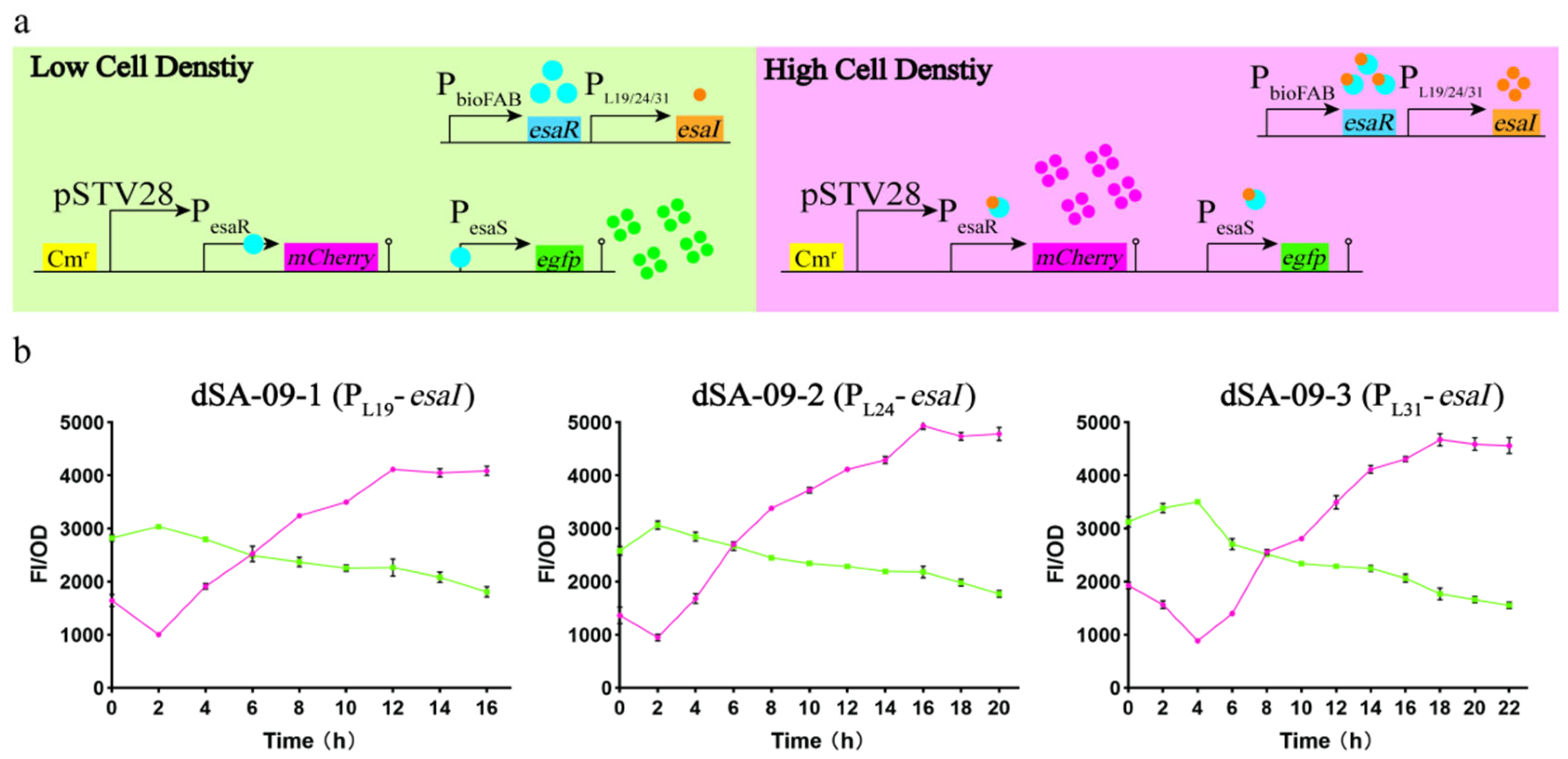
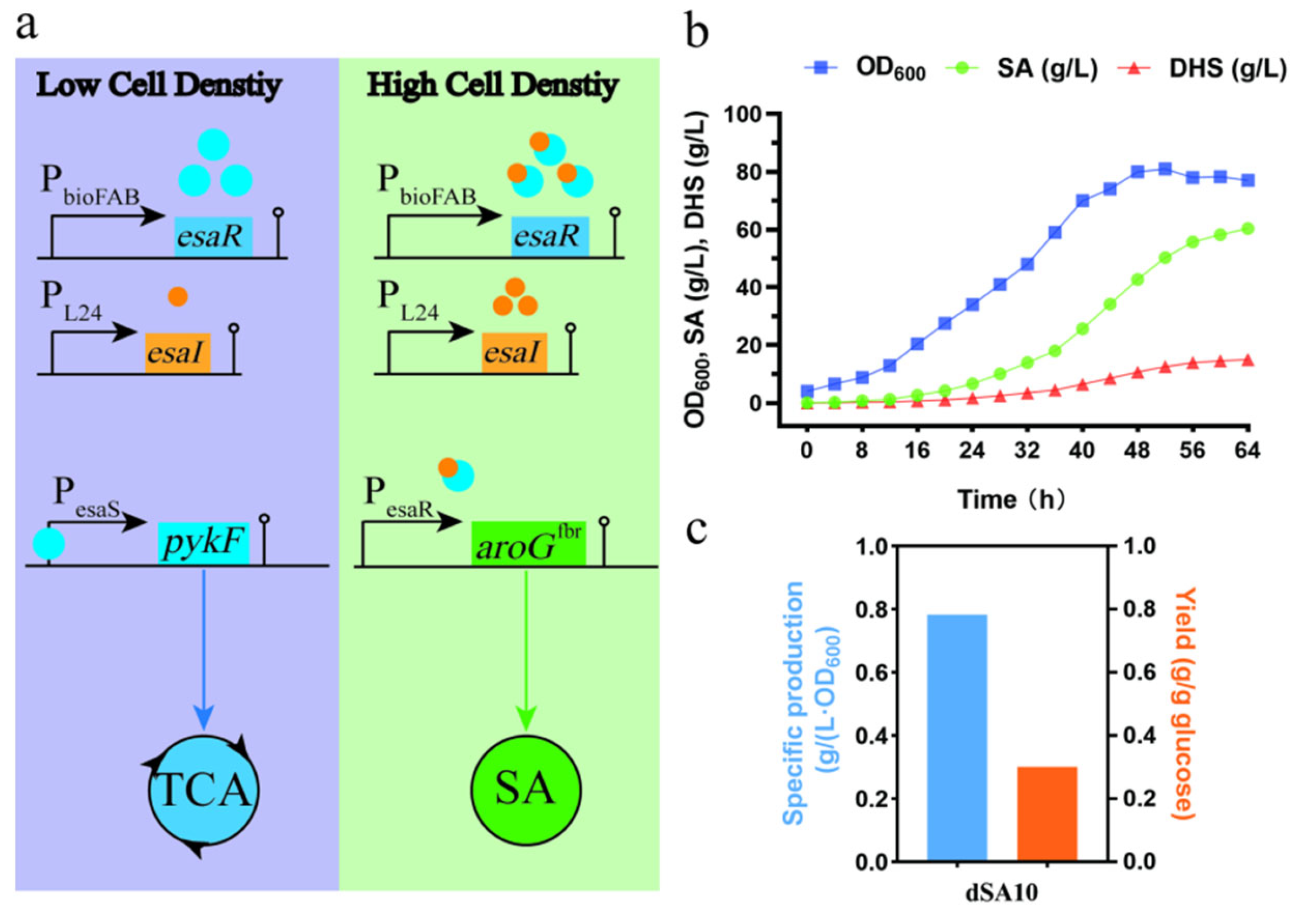
3.4. Strategy for Dynamic Regulation between the Shikimate Pathway and Central Metabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.S.; Wu, F.L.; Zhou, D.; Song, G.T.; Chen, W.J.; Zhang, C.Y.; Wang, Q.H. Cofactor self-sufficient whole-cell biocatalysts for the relay-race synthesis of shikimic acid. Fermentation 2022, 8, 229. [Google Scholar] [CrossRef]
- Rawat, G.; Tripathi, P.; Saxena, R.K. Expanding horizons of shikimic acid. Appl. Microbiol. Biotechnol. 2013, 97, 4277–4287. [Google Scholar] [CrossRef] [PubMed]
- Arceo, E.; Ellman, J.A.; Bergman, R.G. A direct, biomass-based synthesis of benzoic acid: Formic acid-mediated deoxygenation of the glucose-derived materials quinic acid and shikimic acid. ChemSusChem 2010, 3, 811–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.L.; Luo, Y.Q.; Wang, L.; Huang, Y.K.; He, Y.G.; Xie, W.J.; Liu, S.L.; Shi, X.X. Novel stereoselective syntheses of (+)-streptol and (−)-1-epi-streptol starting from naturally abundant (−)-shikimic acid. ACS Omega 2021, 6, 17103–17112. [Google Scholar] [CrossRef]
- Bochkov, D.V.; Sysolyatin, S.V.; Kalashnikov, A.I.; Surmacheva, I. Shikimic acid: Review of its analytical, isolation, and purification techniques from plant and microbial sources. J. Chem. Biol. 2012, 5, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Kogure, T.; Kubota, T.; Suda, M.; Hiraga, K.; Inui, M. Metabolic engineering of Corynebacterium glutamicum for shikimate overproduction by growth-arrested cell reaction. Metab. Eng. 2016, 38, 204–216. [Google Scholar] [CrossRef]
- Ghosh, S.; Banerjee, U.C. Generation of aroE overexpression mutant of Bacillus megaterium for the production of shikimic acid. Microb. Cell Fact. 2015, 14, 69. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gao, C.; Ye, C.; Guo, L.; Liu, J.; Chen, X.; Song, W.; Wu, J.; Liu, L. Systems engineering of Escherichia coli for high-level shikimate production. Metab. Eng. 2023, 75, 1–11. [Google Scholar] [CrossRef]
- Gu, P.F.; Fan, X.Y.; Liang, Q.F.; Qi, Q.S.; Li, Q. Novel technologies combined with traditional metabolic engineering strategies facilitate the construction of shikimate-producing Escherichia coli. Microb. Cell Fact. 2017, 16, 167. [Google Scholar] [CrossRef]
- Lee, H.N.; Seo, S.Y.; Kim, H.J.; Park, J.H.; Park, E.; Choi, S.S.; Lee, S.J.; Kim, E.S. Artificial cell factory design for shikimate production in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab043. [Google Scholar] [CrossRef]
- Bilal, M.; Wang, S.; Iqbal, H.M.N.; Zhao, Y.; Hu, H.; Wang, W.; Zhang, X. Metabolic engineering strategies for enhanced shikimate biosynthesis: Current scenario and future developments. Appl. Microbiol. Biotechnol. 2018, 102, 7759–7773. [Google Scholar] [CrossRef]
- Pontrelli, S.; Chiu, T.Y.; Lan, E.I.; Chen, F.Y.H.; Chang, P.C.; Liao, J.C. Escherichia coli as a host for metabolic engineering. Metab. Eng. 2018, 50, 16–46. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Park, S.Y.; Park, Y.S.; Eun, H.; Lee, S.Y. Metabolic engineering of Escherichia coli for natural product biosynthesis. Trend. Biotechnol. 2020, 38, 745–765. [Google Scholar] [CrossRef]
- Niu, F.X.; He, X.; Huang, Y.B.; Liu, J.Z. Biosensor-guided atmospheric and room-temperature plasma mutagenesis and shuffling for high-level production of shikimic acid from Sucrose in Escherichia coli. J. Agric. Food Chem. 2020, 68, 11765–11773. [Google Scholar] [CrossRef]
- Li, Y.F.; Lin, Z.Q.; Huang, C.; Zhang, Y.; Wang, Z.W.; Tang, Y.J.; Chen, T.; Zhao, X.M. Metabolic engineering of Escherichia coli using CRISPR-Cas9 meditated genome editing. Metab. Eng. 2015, 31, 13–21. [Google Scholar] [CrossRef]
- Escalante, A.; Calderon, R.; Valdivia, A.; de Anda, R.; Hernandez, G.; Ramirez, O.T.; Gosset, G.; Bolivar, F. Metabolic engineering for the production of shikimic acid in an evolved Escherichia coli strain lacking the phosphoenolpyruvate: Carbohydrate phosphotransferase system. Microb. Cell Fact. 2010, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Martinez, J.A.; Baez-Viveros, J.L.; Flores, N.; Hernandez-Chavez, G.; Ramirez, O.T.; Gosset, G.; Bolivar, F. Constitutive expression of selected genes from the pentose phosphate and aromatic pathways increases the shikimic acid yield in high-glucose batch cultures of an Escherichia coli strain lacking PTS and pykF. Microb. Cell Fact. 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Tang, J.L.; Liu, Y.; Zhu, X.N.; Zhang, T.C.; Zhang, X.L. Combinatorial modulation of galP and glk gene expression for improved alternative glucose utilization. Appl. Microbiol. Biotechnol. 2012, 93, 2455–2462. [Google Scholar] [CrossRef]
- Zhang, P.C.; Gao, J.; Zhang, H.Y.; Wang, Y.Z.; Liu, Z.; Lee, S.Y.; Mao, X.Z. Metabolic engineering of Escherichia coli for the production of an antifouling agent zosteric acid. Metab. Eng. 2023, 76, 247–259. [Google Scholar] [CrossRef]
- Parker, C.; Barnell, W.O.; Snoep, J.L.; Ingram, L.O.; Conway, T. Characterization of the Zymomonas mobilis glucose facilitator gene product (glf) in recombinant Escherichia coli. Mol. Microbiol. 1995, 15, 795–802. [Google Scholar] [CrossRef]
- Ikeda, M.; Mizuno, Y.; Awane, S.; Hayashi, M.; Mitsuhashi, S.; Takeno, S. Identification and application of a different glucose uptake system that functions as an alternative to the phosphotransferase system in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2011, 90, 1443–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Montalvo, V.; Martinez, A.; Hernandez-Chavez, G.; Bolivar, F.; Valle, F.; Gosset, G. Expression of galP and glk in a Escherichia coli PTS mutant restores glucose transport and increases glycolytic flux to fermentation products. Biotechnol. Bioeng. 2003, 83, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.; Jallu, S.; Singh, T.P. The shikimate pathway: Review of amino acid sequence, function and three-dimensional structures of the enzymes. Crit. Rev. Microbiol. 2015, 41, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Z.; Li, M.M.; Zhou, L.; Shen, W.; Algasan, G.; Fan, Y.; Wang, Z.X. Metabolic engineering of Escherichia coli for improving shikimate synthesis from glucose. Bioresour. Technol. 2014, 166, 64–71. [Google Scholar] [CrossRef]
- Kim, B.G.; Jung, W.D.; Mok, H.; Ahn, J.H. Production of hydroxycinnamoyl-shikimates and chlorogenic acid in Escherichia coli: Production of hydroxycinnamic acid conjugates. Microb. Cell Fact. 2013, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Garcia, S.; Flores, N.; De Anda, R.; Hernandez, G.; Gosset, G.; Bolivar, F.; Escalante, A. The role of the ydiB Gene, which encodes quinate/shikimate dehydrogenase, in the production of quinic, dehydroshikimic and shikimic acids in a PTS- strain of Escherichia coli. J. Mol. Microbiol. Biotechnol. 2017, 27, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Knop, D.R.; Draths, K.M.; Chandran, S.S.; Barker, J.L.; von Daeniken, R.; Weber, W.; Frost, J.W. Hydroaromatic equilibration during biosynthesis of shikimic acid. J. Am. Chem. Soc. 2001, 123, 10173–10182. [Google Scholar] [CrossRef]
- Kramer, M.; Bongaerts, J.; Bovenberg, R.; Kremer, S.; Muller, U.; Orf, S.; Wubbolts, M.; Raeven, L. Metabolic engineering for microbial production of shikimic acid. Metab. Eng. 2003, 5, 277–283. [Google Scholar] [CrossRef]
- Ikeda, M. Towards bacterial strains overproducing L-tryptophan and other aromatics by metabolic engineering. Appl. Microbiol. Biotechnol. 2006, 69, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Dell, A.; Frost, J.W. Identification and kinetic characterization of 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase from Escherichia coli. Curr. Opin. Plant Biol. 1993, 32, 7948–7953. [Google Scholar]
- Juminaga, D.; Baidoo, E.E.K.; Redding-Johanson, A.M.; Batth, T.S.; Burd, H.; Mukhopadhyay, A.; Petzold, C.J.; Keasling, J.D. Modular engineering of L-tyrosine production in Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zeng, A.P. Rational design and metabolic analysis of Escherichia coli for effective production of L-tryptophan at high concentration. Appl. Microbiol. Biotechnol. 2017, 101, 559–568. [Google Scholar] [CrossRef]
- Du, L.H.; Zhang, Z.; Xu, Q.Y.; Chen, N. Central metabolic pathway modification to improve L-tryptophan production in Escherichia coli. Bioengineered 2019, 10, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.Y.; Meng, X.J.; Jiang, J.; Pang, D.X.; Jones, G.; OuYang, H.S.; Ren, L.Z. Site-directed mutagenesis and over expression of aroG gene of Escherichia coli K-12. Int. J. Biol. Macromol. 2012, 51, 915–919. [Google Scholar] [CrossRef]
- Li, M.J.; Liu, C.Q.; Yang, J.M.; Nian, R.; Xian, M.; Li, F.L.; Zhang, H.B. Common problems associated with the microbial productions of aromatic compounds and corresponding metabolic engineering strategies. Biotechnol. Adv. 2020, 41, 107548. [Google Scholar] [CrossRef]
- Lindner, H.A.; Nadeau, G.; Matte, A.; Michel, G.; Menard, R.; Cygler, M. Site-directed mutagenesis of the active site region in the quinate/shikimate 5-dehydrogenase YdiB of Escherichia coli. J. Biol. Chem. 2005, 280, 7162–7169. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.A.; Christendat, D. Structure of Arabidopsis dehydroquinate dehydratase-shikimate dehydrogenase and implications for metabolic channeling in the shikimate pathway. Biochemistry 2006, 45, 7787–7796. [Google Scholar] [CrossRef]
- Chen, K.; Dou, J.; Tang, S.R.; Yang, Y.S.; Wang, H.; Fang, H.Q.; Zhou, C.L. Deletion of the aroK gene is essential for high shikimic acid accumulation through the shikimate pathway in E. coli. Bioresour. Technol. 2012, 119, 141–147. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Ling, C.; Zhang, Y.Y.; Huang, J.; Liu, J.Z. Production of shikimic acid from Escherichia coli through chemically inducible chromosomal evolution and cofactor metabolic engineering. Microb. Cell Fact. 2014, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.Y.; Hung, W.P.; Tsai, S.H. Improvement of shikimic acid production in Escherichia coli with growth phase-dependent regulation in the biosynthetic pathway from glycerol. World J. Microbiol. Biotechnol. 2017, 33, 25. [Google Scholar] [CrossRef]
- Komera, I.; Gao, C.; Guo, L.; Hu, G.P.; Chen, X.L.; Liu, L.M. Bifunctional optogenetic switch for improving shikimic acid production in E. coli. Biotechnol. Biofuels Bioprod. 2022, 15, 13. [Google Scholar] [CrossRef] [PubMed]
- Romanowski, M.J.; Burley, S.K. Crystal structure of the Escherichia coli shikimate kinase I (AroK) that confers sensitivity to mecillinam. Proteins 2002, 47, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.H.; Gu, Y.J.; Li, Y.; Yan, H.G.; Ji, X.H. Crystal structure of Mycobacterium tuberculosis shikimate kinase in complex with shikimic acid and an ATP analogue. Biochemistry 2006, 45, 8539–8545. [Google Scholar] [CrossRef] [PubMed]
- Sutton, K.A.; Breen, J.; MacDonald, U.; Beanan, J.M.; Olson, R.; Russo, T.A.; Schultz, L.W.; Umland, T.C. Structure of shikimate kinase, an in vivo essential metabolic enzyme in the nosocomial pathogen Acinetobacter baumannii, in complex with shikimate. Acta Crystallogr. D 2015, 71, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.C.; Chang, Y.N.; Wang, W.C. Structural basis for shikimate-binding specificity of Helicobacter pylori shikimate kinase. J. Bacteriol. 2005, 187, 8156–8163. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.C.; Chen, Y.F.; Wang, H.J.; Hsu, K.C.; Lin, S.C.; Chen, T.J.; Yang, J.M.; Wang, W.C. Structures of Helicobacter pylori shikimate kinase reveal a selective inhibitor-induced-fit mechanism. PLoS ONE 2012, 7, e33481. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Lee, D.E.; Lee, B.U.; Kim, H.S. Global analyses of transcriptomes and proteomes of a parent strain and an L-threonine-overproducing mutant strain. J. Bacteriol. 2003, 185, 5442–5451. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.; Zelder, O.; Hafner, S.; Schroder, H.; Wittmann, C. From zero to hero-design-based systems metabolic engineering of Corynebacterium glutamicum for L-lysine production. Metab. Eng. 2011, 13, 159–168. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Lu, T. Autonomous production of 1,4-butanediol via a de novo biosynthesis pathway in engineered Escherichia coli. Metab. Eng. 2015, 29, 135–141. [Google Scholar] [CrossRef]
- Kim, E.M.; Woo, H.M.; Tian, T.; Yilmaz, S.; Javidpour, P.; Keasling, J.D.; Lee, T.S. Autonomous control of metabolic state by a quorum sensing (QS)-mediated regulator for bisabolene production in engineered E. coli. Metab. Eng. 2017, 44, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Reizman, I.M.B.; Reisch, C.R.; Prather, K.L.J. Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit. Nat. Biotechnol. 2017, 35, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Doong, S.J.; Gupta, A.; Prather, K.L.J. Layered dynamic regulation for improving metabolic pathway productivity in Escherichia coli. Proc. Natl. Acad. Sci. USA 2018, 115, 2964–2969. [Google Scholar] [CrossRef] [Green Version]
- Dinh, C.V.; Prather, K.L.J. Development of an autonomous and bifunctional quorum-sensing circuit for metabolic flux control in engineered Escherichia coli. Proc. Natl. Acad. Sci. USA 2019, 116, 25562–25568. [Google Scholar] [CrossRef]
- Shen, Y.P.; Fong, L.S.; Yan, Z.B.; Liu, J.Z. Combining directed evolution of pathway enzymes and dynamic pathway regulation using a quorum-sensing circuit to improve the production of 4-hydroxyphenylacetic acid in Escherichia coli. Biotechnol. Biofuels 2019, 12, 94. [Google Scholar] [CrossRef]
- Gu, F.; Jiang, W.; Mu, Y.L.; Huang, H.; Su, T.Y.; Luo, Y.; Liang, Q.F.; Qi, Q.S. Quorum sensing-based dual-function switch and its application in solving two key metabolic engineering problems. ACS Synth. Biol. 2020, 9, 209–217. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, D.H.; Wang, R.R.; Zhao, C.G.; Ma, Q.; Wu, H.Y.; Xie, X.X. Reconstructing a recycling and nonauxotroph biosynthetic pathway in Escherichia coli toward highly efficient production of L-citrulline. Metab. Eng. 2021, 68, 220–231. [Google Scholar] [CrossRef]
- Ma, Q.; Xia, L.; Wu, H.Y.; Zhuo, M.Y.; Yang, M.Y.; Zhang, Y.; Tan, M.; Zhao, K.X.; Sun, Q.W.; Xu, Q.Y.; et al. Metabolic engineering of Escherichia coli for efficient osmotic stress-free production of compatible solute hydroxyectoine. Biotechnol. Bioeng. 2022, 119, 89–101. [Google Scholar] [CrossRef]
- Shong, J.; Collins, C.H. Engineering the esaR promoter for tunable quorum sensing-dependent gene expression. ACS Synth. Biol. 2013, 2, 568–575. [Google Scholar] [CrossRef]
- Chandran, S.S.; Yi, J.; Draths, K.M.; von Daeniken, R.; Weber, W.; Frost, J.W. Phosphoenolpyruvate availability and the biosynthesis of shikimic acid. Biotechnol. Prog. 2003, 19, 808–814. [Google Scholar] [CrossRef]
- Gu, P.F.; Su, T.Y.; Wang, Q.; Liang, Q.F.; Qi, Q.S. Tunable switch mediated shikimate biosynthesis in an engineered non-auxotrophic Escherichia coli. Sci. Rep. 2016, 6, 29745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.L.; Lin, J.; Hu, H.F.; Zhou, B.; Zhu, B.Q. Site-specific integration and constitutive expression of key genes intov Escherichia coli chromosome increases shikimic acid yields. Enzyme Microb. Technol. 2016, 82, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Hou, J.S.; Xu, D.P.; Guo, L.; Chen, X.L.; Hu, G.P.; Ye, C.; Edwards, H.; Chen, J.; Chen, W.; et al. Programmable biomolecular switches for rewiring flux in Escherichia coli. Nat. Commun. 2019, 10, 3751. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Engineered Strains | Ec AroK Mutants | Corresponding HpAroK Mutants | Relative Activity of HpAroK Mutants (%) [46] | Characteristics [46] |
|---|---|---|---|---|
| dSA06-3 | WT | WT | 100 | - |
| dSA07-1 | none (∆aroK) | - | - | - |
| dSA07-2 | M13A | M10A | 38 | OCORE: contacts with a hydroxyl group of shikimate; relatively conserved |
| dSA07-3 | V47I | V44I | N.A. | OLID: interacts with a trans hydroxyl group of shikimate; relatively conserved |
| dSA07-4 | F51W | F48W | N.A. | OLID: interacts with a trans hydroxyl group of shikimate; relatively conserved |
| dSA07-5 | R60K | R57K | 2 | CX: contacts with a carboxyl of shikimate; strictly conserved |
| dSA07-6 | K118A | E114A | 82 | OCORE: contacts with a hydroxyl group of shikimate OLID: interacts with a trans hydroxyl group of shikimate; relatively conserved |
| dSA07-7 | R140A | R132A | 5 | CX: contacts with a carboxyl of shikimate; strictly conserved |
| E. coli Strains | Strategies | Shikimate Titer | Ref. |
|---|---|---|---|
| SP1.1pts/pSC6.090B | inactivation of aroL, aroK, ptsH, ptsI, and crr; overexpression of aroB, glf, glk, aroFfbr, tktA, aroE, and serA | 87.00 | [60] |
| PB12.SA22 | inactivation of aroK and aroL; coexpression of aroGfbr, aroB, tktA, and aroE; inactivation of PTS | 7.05 | [16] |
| AR36 | inactivation of PTS aroK, aroL, and pykF; overexpression of aroB, tktA, aroGfbr, aroE, aroD, and zwf | 43.00 | [17] |
| SA116 | inactivation of aroK and aroL; integration of aroGfbr, aroB, aroE, and tktA; overexpression of csrB, pps, and pntAB | 3.12 | [40] |
| P9 | switch regulation of aroK; inactivation of araC, pta, ptsG, aroL, trpR, and pykF | 13.15 | [61] |
| SA5/pGBAE | inactivation of ptsH, ptsI, crr, aroK and aroL; integration of glk, galP, aroG, aroB, tktA and aroE; integration of ppsA at tyrR | 27.41 | [62] |
| DS7 | PTS system was genomically replaced by a glucose facilitator protein gene, Zmglf, from Z. mobilis; inactivation of aroK and aroL; overexpression of tktA, aroG, aroB, and aroE | 12.63 | [63] |
| Inha 224 | inactivation of aroK, aroL, tyrR, ptsG, pykA, and shiA; overexpression of aroB, aroD, aroG, aroF, ppsA, aroE, galP, and tktA | 101.00 | [10] |
| SA09 | inactivation of aroK and aroL; overexpression of glyA, talB, aroG, aroD, and aroEL241I/T61W, tktA; deletion of ptsH, ptsI, pykF, dhal, and ydiB | 126.40 | [8] |
| dSA10 | ptsG was genomically replaced by galP; inactivation of aroL, tyrR, ydiB, shiA, ydiN and pykA; mutation of aroKM13A; overexpression of glk, tktA, aroB, and aroD-aroE; dynamic regulation of pykF and aroGS211F | 60.31 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bo, T.; Wu, C.; Wang, Z.; Jiang, H.; Wang, F.; Chen, N.; Li, Y. Multiple Metabolic Engineering Strategies to Improve Shikimate Titer in Escherichia coli. Metabolites 2023, 13, 747. https://doi.org/10.3390/metabo13060747
Bo T, Wu C, Wang Z, Jiang H, Wang F, Chen N, Li Y. Multiple Metabolic Engineering Strategies to Improve Shikimate Titer in Escherichia coli. Metabolites. 2023; 13(6):747. https://doi.org/10.3390/metabo13060747
Chicago/Turabian StyleBo, Taidong, Chen Wu, Zeting Wang, Hao Jiang, Feiao Wang, Ning Chen, and Yanjun Li. 2023. "Multiple Metabolic Engineering Strategies to Improve Shikimate Titer in Escherichia coli" Metabolites 13, no. 6: 747. https://doi.org/10.3390/metabo13060747