

Expression Silencing of Mitogen-Activated Protein Kinase 8 Interacting Protein-1 Conferred Its Role in Pancreatic β-Cell Physiology and Insulin Secretion
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of RNA-seq Data in Human Pancreatic Islets
2.2. Screening for T2D-Associated Genetic Variants in MAPK8IP1
2.3. Culturing of INS-1 Cell Line and siRNA Transfection
2.4. RNA Extraction and qRT-PCR
2.5. Insulin Secretion and Insulin Content Assay
2.6. Glucose Uptake
2.7. Cell Viability Assay
2.8. Apoptosis Assay
2.9. Reactive Oxygen Species (ROS) Measurements
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
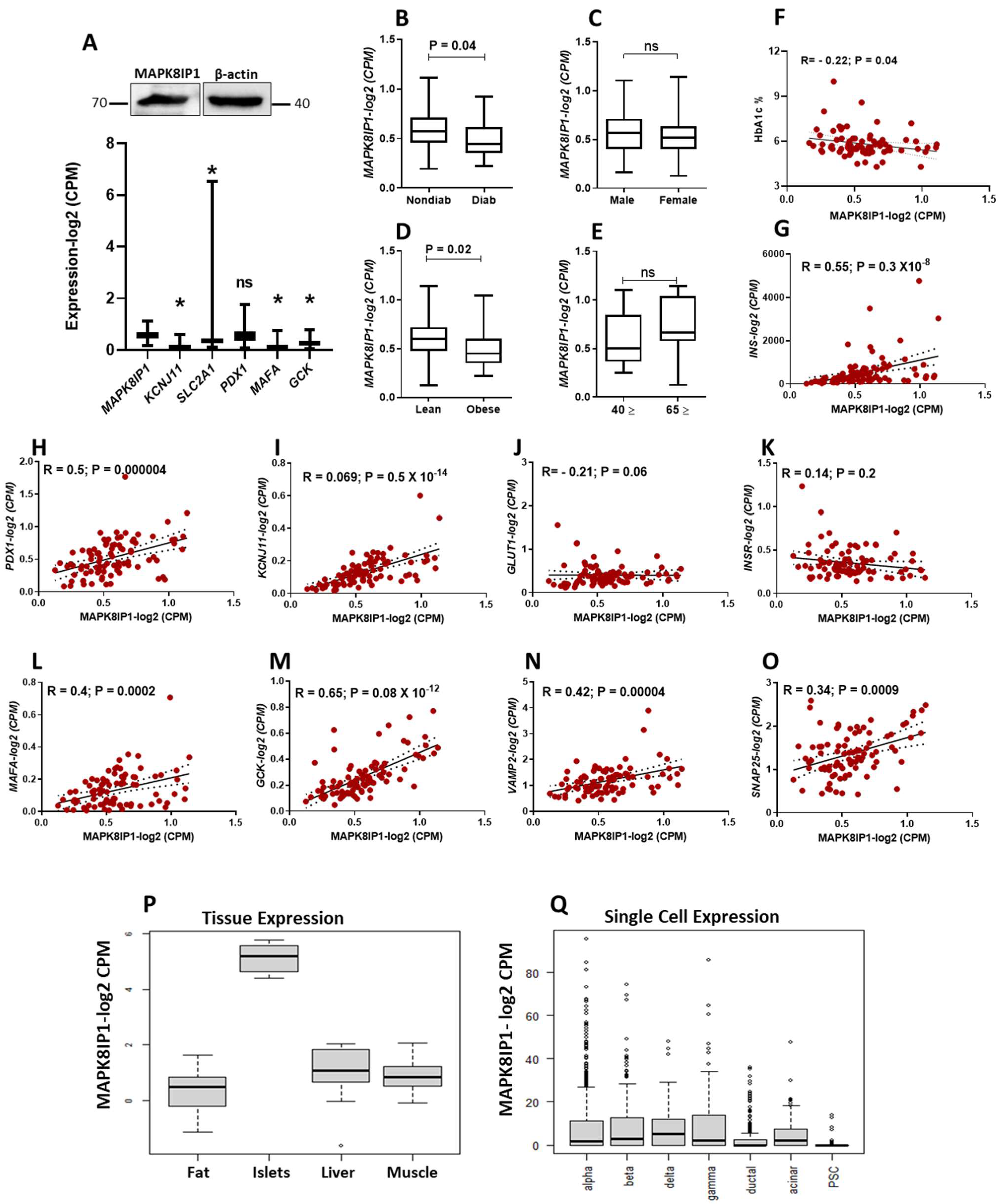
3.1. Expression Profile of MAPK8IP1 in Diabetic and Non-Diabetic Human Pancreatic Islets
3.2. Exploration of T2D Genetic Variants in MAPK8IP1
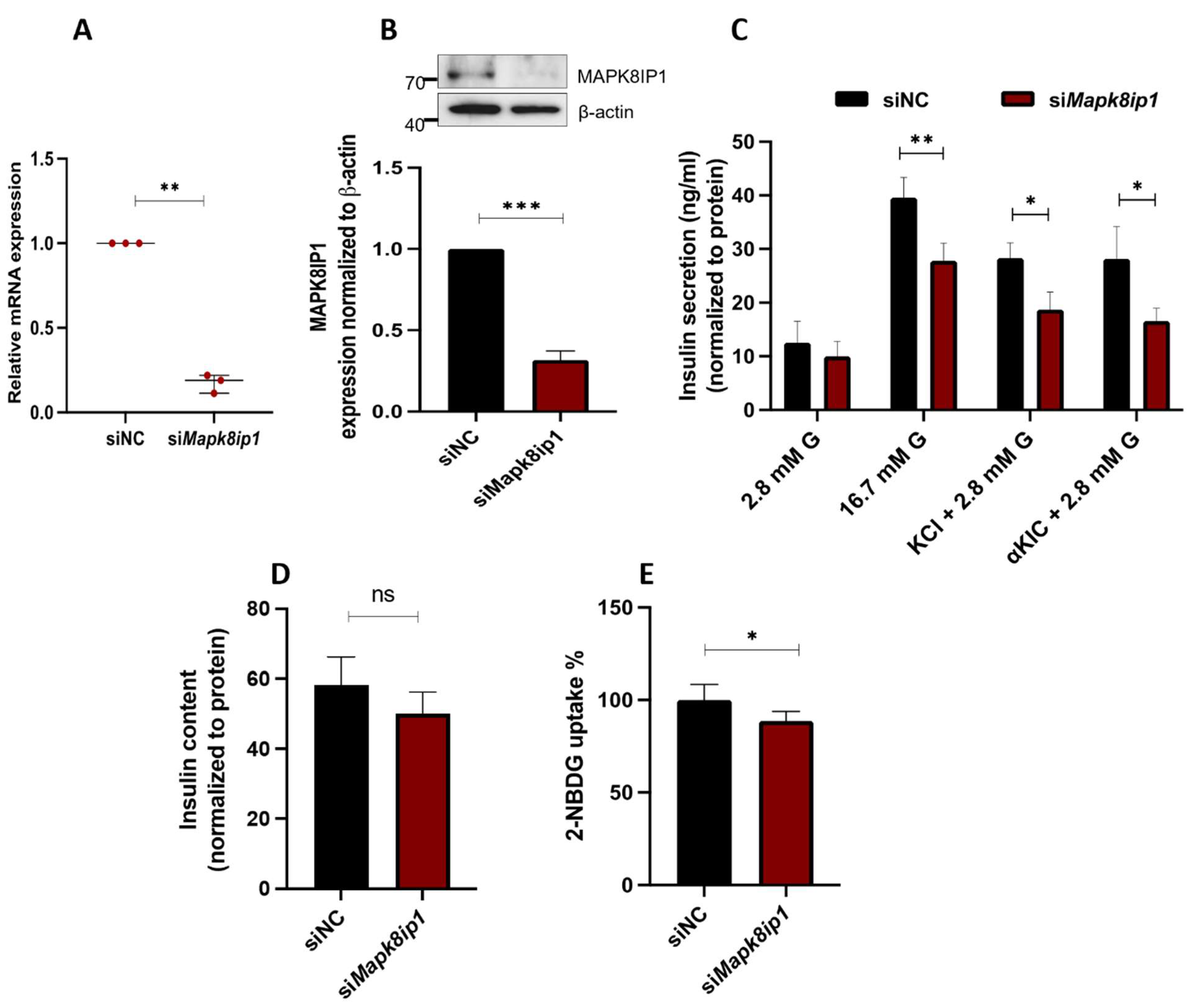
3.3. Silencing of Mapk8ip1 in INS-1 (832/13) Cells Affects Insulin Secretion and Glucose Uptake
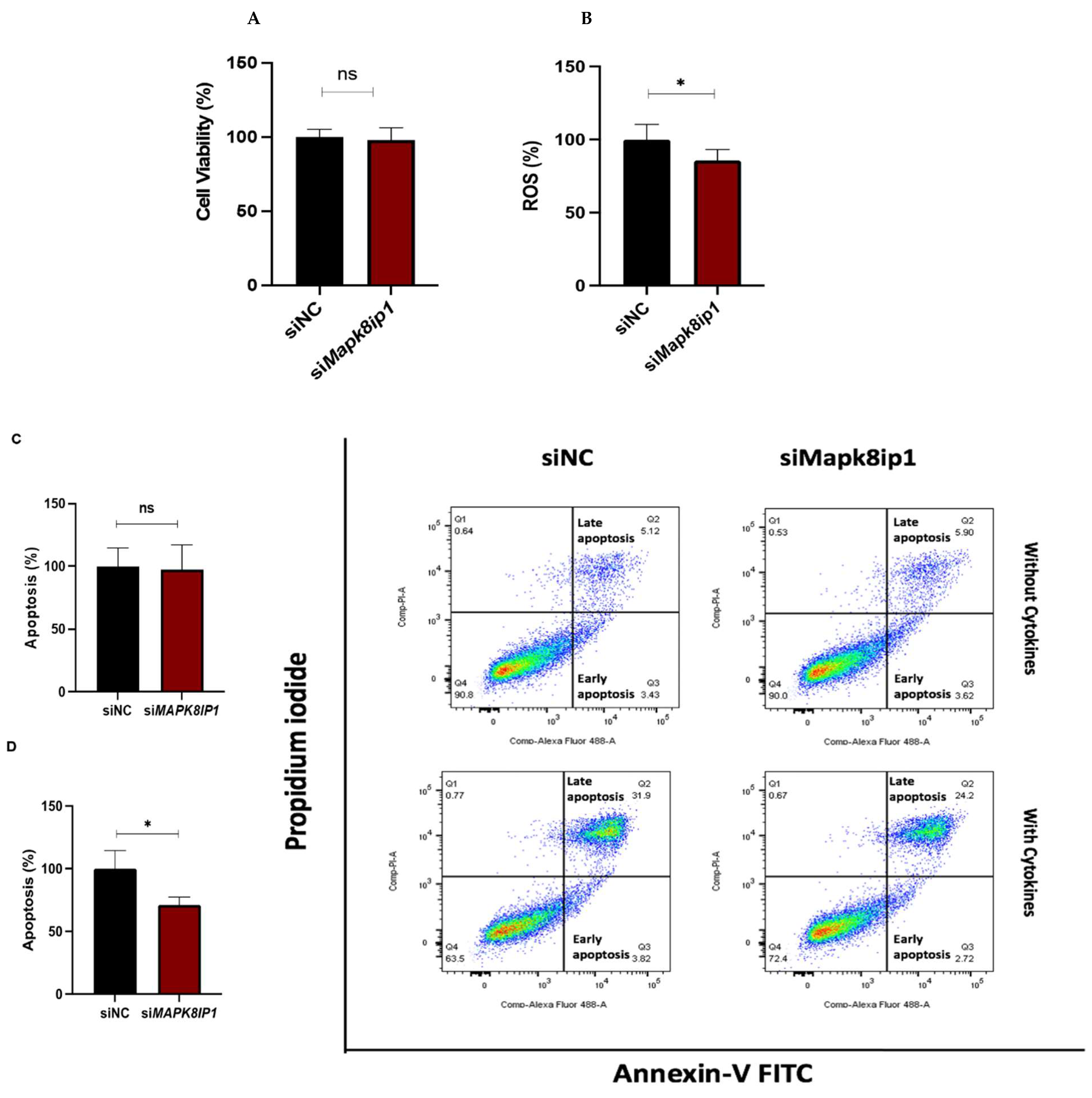
3.4. Effect of Mapk8ip1 Silencing on Cell Viability, Apoptosis, and ROS in INS-1 (832/13) Cells
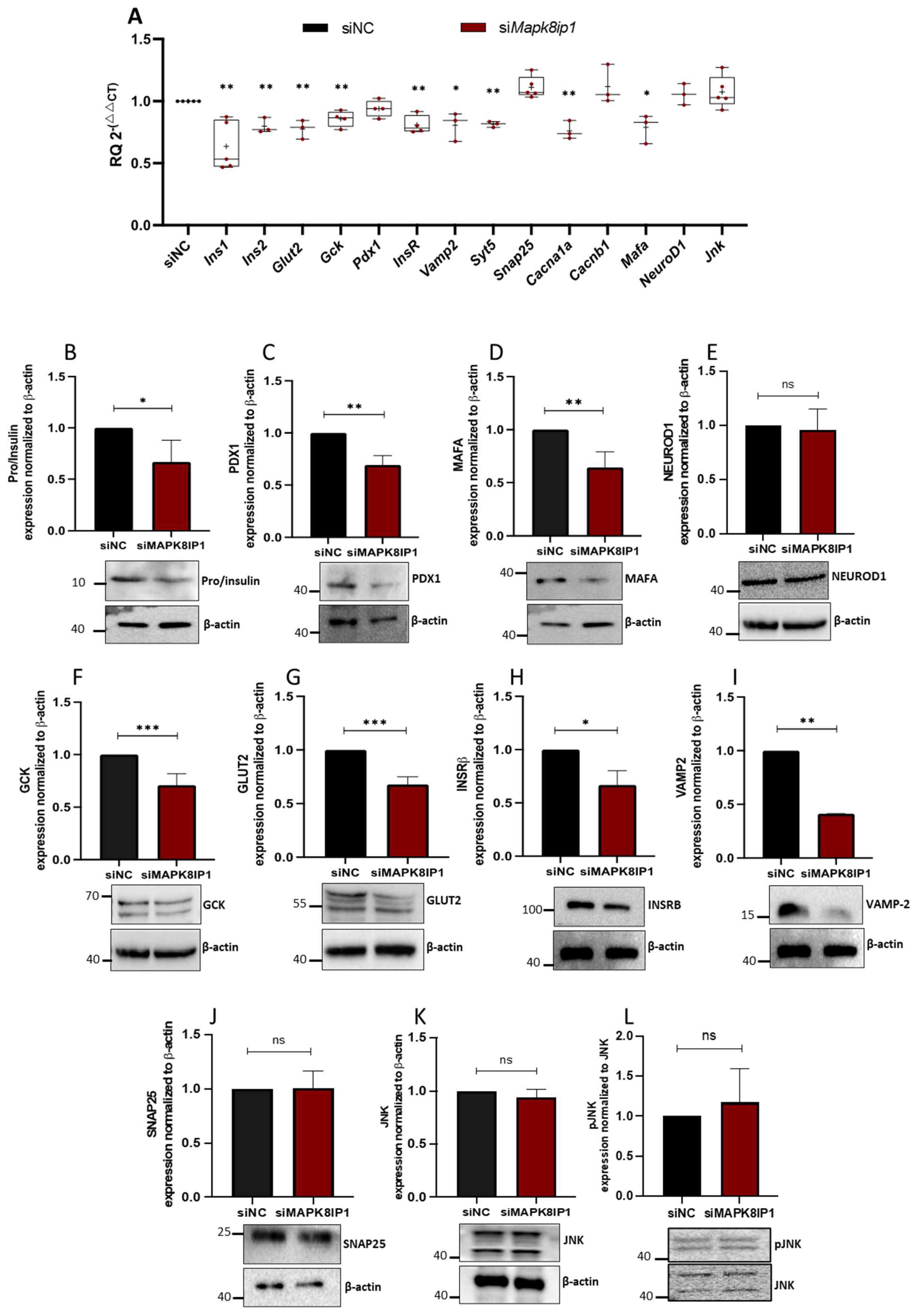
3.5. Mapk8ip1 Silencing Modifies the Expression of β-Cell Function-Related Genes
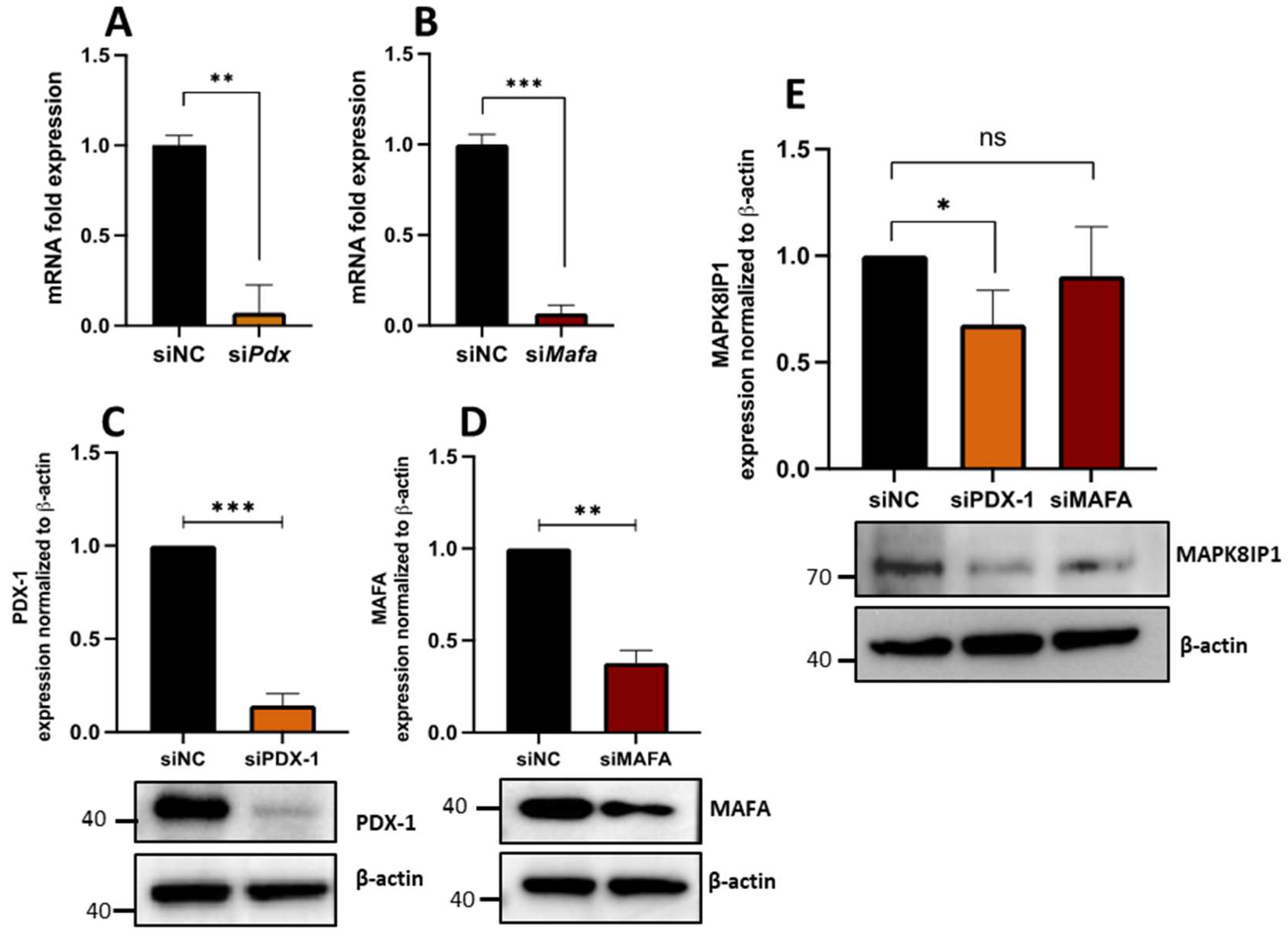
3.6. PDX1 Is Required for Proper MAPK8IP1 Expression in β-Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Ježek, P.; Jabůrek, M.; Plecitá-Hlavatá, L. Contribution of Oxidative Stress and Impaired Biogenesis of Pancreatic β-Cells to Type 2 Diabetes. Antioxid. Redox Signal. 2019, 31, 722–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khin, P.-P.; Lee, J.-H.; Jun, H.-S. A Brief Review of the Mechanisms of β-Cell Dedifferentiation in Type 2 Diabetes. Nutrients 2021, 13, 1593. [Google Scholar] [CrossRef] [PubMed]
- Yung, J.H.M.; Giacca, A. Role of c-Jun N-terminal Kinase (JNK) in Obesity and Type 2 Diabetes. Cells 2020, 9, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanuza-Masdeu, J.; Arévalo, M.I.; Vila, C.; Barberà, A.; Gomis, R.; Caelles, C. In vivo JNK activation in pancreatic β-cells leads to glucose intolerance caused by insulin resistance in pancreas. Diabetes 2013, 62, 2308–2317. [Google Scholar] [CrossRef] [Green Version]
- Kaneto, H.; Matsuoka, T.-A. Involvement of oxidative stress in suppression of insulin biosynthesis under diabetic conditions. Int. J. Mol. Sci. 2012, 13, 13680–13690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmarsh, A.J.; Cavanagh, J.; Tournier, C.; Yasuda, J.; Davis, R.J. A mammalian scaffold complex that selectively mediates MAP kinase activation. Science 1998, 281, 1671–1674. [Google Scholar] [CrossRef]
- Bonny, C.; Nicod, P.; Waeber, G. IB1, a JIP-1-related nuclear protein present in insulin-secreting cells. J. Biol. Chem. 1998, 273, 1843–1846. [Google Scholar] [CrossRef] [Green Version]
- Pellet, J.B.; Haefliger, J.A.; Staple, J.K.; Widmann, C.; Welker, E.; Hirling, H.; Bonny, C.; Nicod, P.; Catsicas, S.; Waeber, G.; et al. Spatial, temporal and subcellular localization of islet-brain 1 (IB1), a homologue of JIP-1, in mouse brain. Eur. J. Neurosci. 2000, 12, 621–632. [Google Scholar] [CrossRef]
- Tawadros, T.; Formenton, A.; Dudler, J.; Thompson, N.; Nicod, P.; Leisinger, H.J.; Waeber, G.; Haefliger, J.A. The scaffold protein IB1/JIP-1 controls the activation of JNK in rat stressed urothelium. J. Cell Sci. 2002, 115, 385–393. [Google Scholar] [CrossRef]
- Jaeschke, A.; Czech, M.P.; Davis, R.J. An essential role of the JIP1 scaffold protein for JNK activation in adipose tissue. Genes Dev. 2004, 18, 1976–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abderrahmani, A.; Steinmann, M.; Plaisance, V.; Niederhauser, G.; Haefliger, J.A.; Mooser, V.; Bonny, C.; Nicod, P.; Waeber, G. The transcriptional repressor REST determines the cell-specific expression of the human MAPK8IP1 gene encoding IB1 (JIP-1). Mol. Cell. Biol. 2001, 21, 7256–7267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nihalani, D.; Wong, H.; Verma, R.; Holzman, L.B. Src family kinases directly regulate JIP1 module dynamics and activation. Mol. Cell. Biol. 2007, 27, 2431–2441. [Google Scholar] [CrossRef] [Green Version]
- Dajas-Bailador, F.; Jones, E.V.; Whitmarsh, A.J. The JIP1 scaffold protein regulates axonal development in cortical neurons. Curr. Biol. 2008, 18, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Lee, Y.J. Dissociation of Akt1 from its negative regulator JIP1 is mediated through the ASK1-MEK-JNK signal transduction pathway during metabolic oxidative stress: A negative feedback loop. J. Cell Biol. 2005, 170, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Standen, C.L.; Kennedy, N.J.; Flavell, R.A.; Davis, R.J. Signal transduction cross talk mediated by Jun N-terminal kinase-interacting protein and insulin receptor substrate scaffold protein complexes. Mol. Cell Biol. 2009, 29, 4831–4840. [Google Scholar] [CrossRef] [Green Version]
- Fu, M.-m.; Holzbaur, E.L.F. JIP1 regulates the directionality of APP axonal transport by coordinating kinesin and dynein motors. J. Cell Biol. 2013, 202, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Waeber, G.; Delplanque, J.; Bonny, C.; Mooser, V.; Steinmann, M.; Widmann, C.; Maillard, A.; Miklossy, J.; Dina, C.; Hani, E.H.; et al. The gene MAPK8IP1, encoding islet-brain-1, is a candidate for type 2 diabetes. Nat. Genet. 2000, 24, 291–295. [Google Scholar] [CrossRef]
- Whitmarsh, A.J.; Kuan, C.Y.; Kennedy, N.J.; Kelkar, N.; Haydar, T.F.; Mordes, J.P.; Appel, M.; Rossini, A.A.; Jones, S.N.; Flavell, R.A.; et al. Requirement of the JIP1 scaffold protein for stress-induced JNK activation. Genes Dev. 2001, 15, 2421–2432. [Google Scholar] [CrossRef] [Green Version]
- Kant, S.; Standen, C.L.; Morel, C.; Jung, D.Y.; Kim, J.K.; Swat, W.; Flavell, R.A.; Davis, R.J. A Protein Scaffold Coordinates SRC-Mediated JNK Activation in Response to Metabolic Stress. Cell Rep. 2017, 20, 2775–2783. [Google Scholar] [CrossRef] [Green Version]
- Bonny, C.; Oberson, A.; Steinmann, M.; Schorderet, D.F.; Nicod, P.; Waeber, G. IB1 Reduces Cytokine-induced Apoptosis of Insulin-secreting Cells. J. Biol. Chem. 2000, 275, 16466–16472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Z.; Van de Casteele, M.; Dong, J.; Heimberg, H.; Haefliger, J.A.; Waeber, G.; Schuit, F.; Pipeleers, D. Variations in IB1/JIP1 expression regulate susceptibility of beta-cells to cytokine-induced apoptosis irrespective of C-Jun NH2-terminal kinase signaling. Diabetes 2003, 52, 2497–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadista, J.; Vikman, P.; Laakso, E.O.; Mollet, I.G.; Esguerra, J.L.; Taneera, J.; Storm, P.; Osmark, P.; Ladenvall, C.; Prasad, R.B.; et al. Global genomic and transcriptomic analysis of human pancreatic islets reveals novel genes influencing glucose metabolism. Proc. Natl. Acad. Sci. USA 2014, 111, 13924–13929. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Lang, S.; Sharma, A.; Fadista, J.; Zhou, Y.; Ahlqvist, E.; Jonsson, A.; Lyssenko, V.; Vikman, P.; Hansson, O.; et al. A systems genetics approach identifies genes and pathways for type 2 diabetes in human islets. Cell Metab. 2012, 16, 122–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, L.; Piron, A.; Moran, I.; Guindo-Martinez, M.; Bonas-Guarch, S.; Atla, G.; Miguel-Escalada, I.; Royo, R.; Puiggros, M.; Garcia-Hurtado, X. TIGER: The gene expression regulatory variation landscape of human pancreatic islets. bioRxiv 2021, 37, 109807. [Google Scholar] [CrossRef]
- Hohmeier, H.E.; Mulder, H.; Chen, G.; Henkel-Rieger, R.; Prentki, M.; Newgard, C.B. Isolation of INS-1-derived cell lines with robust ATP-sensitive K+ channel-dependent and -independent glucose-stimulated insulin secretion. Diabetes 2000, 49, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Dhaiban, S.; Hachim, M.; Mohammed, A.K.; Mukhopadhyay, D.; Bajbouj, K.; Hamoudi, R.; Salehi, A.; Hamad, M. Reduced expression of Chl1 gene impairs insulin secretion by down-regulating the expression of key molecules of β-cell function. Exp. Clin. Endocrinol. Diabetes 2019, 129, 864–872. [Google Scholar] [CrossRef]
- El-Huneidi, W.; Anjum, S.; Mohammed, A.K.; Unnikannan, H.; Saeed, R.; Bajbouj, K.; Abu-Gharbieh, E.; Taneera, J. Copine 3 “CPNE3” is a novel regulator for insulin secretion and glucose uptake in pancreatic β-cells. Sci. Rep. 2021, 11, 20692. [Google Scholar] [CrossRef]
- Segerstolpe, Å.; Palasantza, A.; Eliasson, P.; Andersson, E.-M.; Andréasson, A.-C.; Sun, X.; Picelli, S.; Sabirsh, A.; Clausen, M.; Bjursell, M.K.; et al. Single-Cell Transcriptome Profiling of Human Pancreatic Islets in Health and Type 2 Diabetes. Cell Metab. 2016, 24, 593–607. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Moriguchi, T.; Kajihara, M.; Esaki, R.; Harada, A.; Shimohata, H.; Oishi, H.; Hamada, M.; Morito, N.; Hasegawa, K. MafA is a key regulator of glucose-stimulated insulin secretion. Mol. Cell Biol. 2005, 25, 4969–4976. [Google Scholar] [CrossRef] [Green Version]
- Hang, Y.; Yamamoto, T.; Benninger, R.K.; Brissova, M.; Guo, M.; Bush, W.; Piston, D.W.; Powers, A.C.; Magnuson, M.; Thurmond, D.C. The MafA transcription factor becomes essential to islet β-cells soon after birth. Diabetes 2014, 63, 1994–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, K.; Polonsky, K.S. Pdx1 and other factors that regulate pancreatic β-cell survival. Diabetes Obes. Metab. 2009, 11, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, P.; Bugliani, M.; De Tata, V.; Suleiman, M.; Marselli, L. Pancreatic beta cell identity in humans and the role of type 2 diabetes. Front. Cell Dev. Biol. 2017, 5, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramswig, N.C.; Everett, L.J.; Schug, J.; Dorrell, C.; Liu, C.; Luo, Y.; Streeter, P.R.; Naji, A.; Grompe, M.; Kaestner, K.H. Epigenomic plasticity enables human pancreatic α to β cell reprogramming. J. Clin. Investig. 2013, 123, 1275–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottosson-Laakso, E.; Krus, U.; Storm, P.; Prasad, R.B.; Oskolkov, N.; Ahlqvist, E.; Fadista, J.; Hansson, O.; Groop, L.; Vikman, P. Glucose-induced changes in gene expression in human pancreatic islets: Causes or consequences of chronic hyperglycemia. Diabetes 2017, 66, 3013–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.S.; Zhu, D.L.; Zhou, X.R.; Rong, Y.; Zeng, M.; Chen, J.B.; Jiang, F.; Tuo, X.M.; Feng, Z.; Yang, T.L.; et al. An Intronic Risk SNP rs12454712 for Central Obesity Acts As an Allele-Specific Enhancer To Regulate BCL2 Expression. Diabetes 2021, 70, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haefliger, J.A.; Tawadros, T.; Meylan, L.; Gurun, S.L.; Roehrich, M.E.; Martin, D.; Thorens, B.; Waeber, G. The scaffold protein IB1/JIP-1 is a critical mediator of cytokine-induced apoptosis in pancreatic beta cells. J. Cell Sci. 2003, 116, 1463–1469. [Google Scholar] [CrossRef] [Green Version]
- Im, J.Y.; Lee, K.W.; Kim, M.H.; Lee, S.H.; Ha, H.Y.; Cho, I.H.; Kim, D.; Yu, M.S.; Kim, J.B.; Lee, J.K.; et al. Repression of phospho-JNK and infarct volume in ischemic brain of JIP1-deficient mice. J. Neurosci. Res. 2003, 74, 326–332. [Google Scholar] [CrossRef]
- Abdelli, S.; Puyal, J.; Bielmann, C.; Buchillier, V.; Abderrahmani, A.; Clarke, P.G.; Beckmann, J.S.; Bonny, C. JNK3 is abundant in insulin-secreting cells and protects against cytokine-induced apoptosis. Diabetologia 2009, 52, 1871–1880. [Google Scholar] [CrossRef] [Green Version]
- Prause, M.; Christensen, D.P.; Billestrup, N.; Mandrup-Poulsen, T. JNK1 protects against glucolipotoxicity-mediated beta-cell apoptosis. PLoS ONE 2014, 9, e87067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguchi, N.; Vaziri, N.D.; Dafoe, D.C.; Ichii, H. The Role of Oxidative Stress in Pancreatic β Cell Dysfunction in Diabetes. Int. J. Mol. Sci. 2021, 22, 1509. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Zhang, Q.; Andersen, M.E. Reactive Oxygen Species and Antioxidants in Pancreatic β-Cell Function—Yin and Yang. In Systems Biology of Free Radicals and Antioxidants; Laher, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3319–3337. [Google Scholar]
- Brissova, M.; Shiota, M.; Nicholson, W.E.; Gannon, M.; Knobel, S.M.; Piston, D.W.; Wright, C.V.; Powers, A.C. Reduction in pancreatic transcription factor PDX-1 impairs glucose-stimulated insulin secretion. J. Biol. Chem. 2002, 277, 11225–11232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchet, E.; Van de Velde, S.; Matsumura, S.; Hao, E.; LeLay, J.; Kaestner, K.; Montminy, M. Feedback inhibition of CREB signaling promotes beta cell dysfunction in insulin resistance. Cell Rep. 2015, 10, 1149–1157. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, W.; Takahashi, S.; Yasuda, K. MafA is critical for maintenance of the mature beta cell phenotype in mice. Diabetologia 2015, 58, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Thorens, B.; Guillam, M.-T.r.s.; Beermann, F.; Burcelin, R.m.; Jaquet, M. Transgenic reexpression of GLUT1 or GLUT2 in pancreatic β cells rescues GLUT2-null mice from early death and restores normal glucose-stimulated insulin secretion. J. Biol. Chem. 2000, 275, 23751–23758. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Mussa, B.; Saber-Ayad, M.; Dhaiban, S.; Aljaibeji, H.; Sulaiman, N. Maturity-onset diabetes of the young: An overview with focus on the middle east. Curr. Mol. Med. 2017, 17, 549–562. [Google Scholar] [CrossRef]
- Wang, J.; Gu, W.; Chen, C. Knocking down insulin receptor in pancreatic beta cell lines with lentiviral-small hairpin RNA reduces glucose-stimulated insulin secretion via decreasing the gene expression of insulin, GLUT2 and Pdx1. Int. J. Mol. Sci. 2018, 19, 985. [Google Scholar] [CrossRef] [Green Version]
- Regazzi, R.; Wollheim, C.B.; Lang, J.; Theler, J.M.; Rossetto, O.; Montecucco, C.; Sadoul, K.; Weller, U.; Palmer, M.; Thorens, B. VAMP-2 and cellubrevin are expressed in pancreatic beta-cells and are essential for Ca(2+)-but not for GTP gamma S-induced insulin secretion. EMBO J. 1995, 14, 2723–2730. [Google Scholar] [CrossRef]
- Iezzi, M.; Kouri, G.; Fukuda, M.; Wollheim, C.B. Synaptotagmin V and IX isoforms control Ca2+ -dependent insulin exocytosis. J. Cell Sci. 2004, 117, 3119–3127. [Google Scholar] [CrossRef] [Green Version]
- Tuluc, P.; Theiner, T.; Jacobo-Piqueras, N.; Geisler, S.M. Role of High Voltage-Gated Ca2+ Channel Subunits in Pancreatic β-Cell Insulin Release. From Structure to Function. Cells 2021, 10, 2004. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Symbol | Accession Numbers | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
|---|---|---|---|
| MafA | NM_001399773.1 | GAGGAGGAGCGCAAGATCGG | AGCAAAAGTTTCGTGCTGTCAA |
| NeuroD1 | NM_019218.3 | CCCTAACTGATTGCACCAGC | TGCAGGGTAGTGCATGGTAA |
| Syt5 | NM_019350.2 | CACCTGACCCCAGATCCTTT | GAGTGGTACTGGAAGTCGGA |
| Snap25 |
NM_001270576.1 NM_001270575.1 NM_030991.3 | GGCGTTTGCTGAATGACAAC | CAGAGCCTGACACCCTAAGA |
| Cacna1a | NM_012918.5 | CTAGCCCTGCCAAGATCGG | ACGATAAGGCTGTTCTCGG |
| Cacnb1 | NM_017346.2 | CTTTACCCCAGCAACCACCC | GTCCACACACGAGTCTCCTG |
| Vamp2 | NM_012663.2 | TGGTGGACATCATGAGGGTG | GCTTGGCTGCACTTGTTTCA |
| Jnk | NM_053829.2 | TCCAGTTCTCGTACCCGCTA | AGCATGGCGTGACACAGTAA |
| Hprt1 | NM_012583.2 | TTGTGTCATCAGCGAAAGTGG | CACAGGACTAGAACGTCTGCT |
| ID | Reference Allele | Alternate Allele | Sample Size | p-Value |
|---|---|---|---|---|
| Diagram Diamante | ||||
| rs7115753 | A | G | 231,420 | 4.08 × 10−9 |
| Diagram 1000G | ||||
| rs11038677 | T | C | - | 3.70 × 10−5 |
| 70K for T2D | ||||
| rs553011963 | CGTT | AGTT | 66,940 | 0.0013 |
| Phenotype | p-Value | Beta | Odds Ratio | Sample Size |
|---|---|---|---|---|
| rs7115753 | ||||
| Fasting glucose adj BMI | 9.76 × 10−51 | ▼−0.0246 | 287,195 | |
| Fasting glucose | 2.75 × 10−13 | ▼−0.0298 | 263,726 | |
| Type 2 diabetes adj BMI | 1.37 × 10−7 | ▼0.9635 | 266,778 | |
| Type 2 diabetes | 1.62 × 10−7 | ▼0.9635 | 1,436,100 | |
| rs7115753 | ||||
| Fasting glucose adj BMI | 2.45 × 10−24 | ▼−0.0185 | 287,195 | |
| Fasting glucose | 2.371 × 10−8 | ▼−0.0212 | 277,462 | |
| Type 2 diabetes adj BMI | 3.59 × 10−7 | ▼0.9570 | 266,778 | |
| Type 2 diabetes | 0.00004 | ▼0.9813 | 1,436,100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, R.; Mohammed, A.K.; Saleh, S.E.; Aboshanab, K.M.; Aboulwafa, M.M.; Taneera, J. Expression Silencing of Mitogen-Activated Protein Kinase 8 Interacting Protein-1 Conferred Its Role in Pancreatic β-Cell Physiology and Insulin Secretion. Metabolites 2023, 13, 307. https://doi.org/10.3390/metabo13020307
Saeed R, Mohammed AK, Saleh SE, Aboshanab KM, Aboulwafa MM, Taneera J. Expression Silencing of Mitogen-Activated Protein Kinase 8 Interacting Protein-1 Conferred Its Role in Pancreatic β-Cell Physiology and Insulin Secretion. Metabolites. 2023; 13(2):307. https://doi.org/10.3390/metabo13020307
Chicago/Turabian StyleSaeed, Rania, Abdul Khader Mohammed, Sarra E. Saleh, Khaled M. Aboshanab, Mohammad M. Aboulwafa, and Jalal Taneera. 2023. "Expression Silencing of Mitogen-Activated Protein Kinase 8 Interacting Protein-1 Conferred Its Role in Pancreatic β-Cell Physiology and Insulin Secretion" Metabolites 13, no. 2: 307. https://doi.org/10.3390/metabo13020307