

Metabolites of Geum aleppicum and Sibbaldianthe bifurca: Diversity and α-Glucosidase Inhibitory Potential



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals
2.3. Plant Extracts Preparation
2.4. High-Performance Liquid Chromatography with Photodiode Array Detection and Electrospray Ionization Triple Quadrupole Mass Spectrometric Detection (HPLC-PDA-ESI-tQ-MS/MS) Metabolite Profiling
2.5. HPLC-PDA-ESI-tQ-MS/MS Metabolite Quantification
2.6. HPLC Activity-Based Profiling
2.7. Statistical Analysis
3. Results and Discussion
3.1. Metabolites of Geum aleppicum Herb: HPLC-PDA-ESI-tQ-MS/MS Profile
3.1.1. Carbohydrates
3.1.2. Organic Acids
3.1.3. Benzoic Acid Derivatives
3.1.4. Ellagic Acid Derivatives and Ellagitannins
3.1.5. Flavonoids
3.1.6. Triterpenoids
3.2. Metabolites of Sibbaldianthe bifurca Herb: HPLC-PDA-ESI-tQ-MS/MS Profile
3.2.1. Carbohydrates and Organic Acids
3.2.2. Galloyl O-Glycosides
3.2.3. Benzoic Acid Derivatives
3.2.4. Ellagic Acid Derivatives and Ellagitannins
3.2.5. Flavonoids
3.3. Quantitative Content and Seasonal Variation of Profile of Geum aleppicum and Sibbaldianthe bifurca Herb
3.4. Chemotaxonomic Significance of G. aleppicum and S. bifurca Metabolites
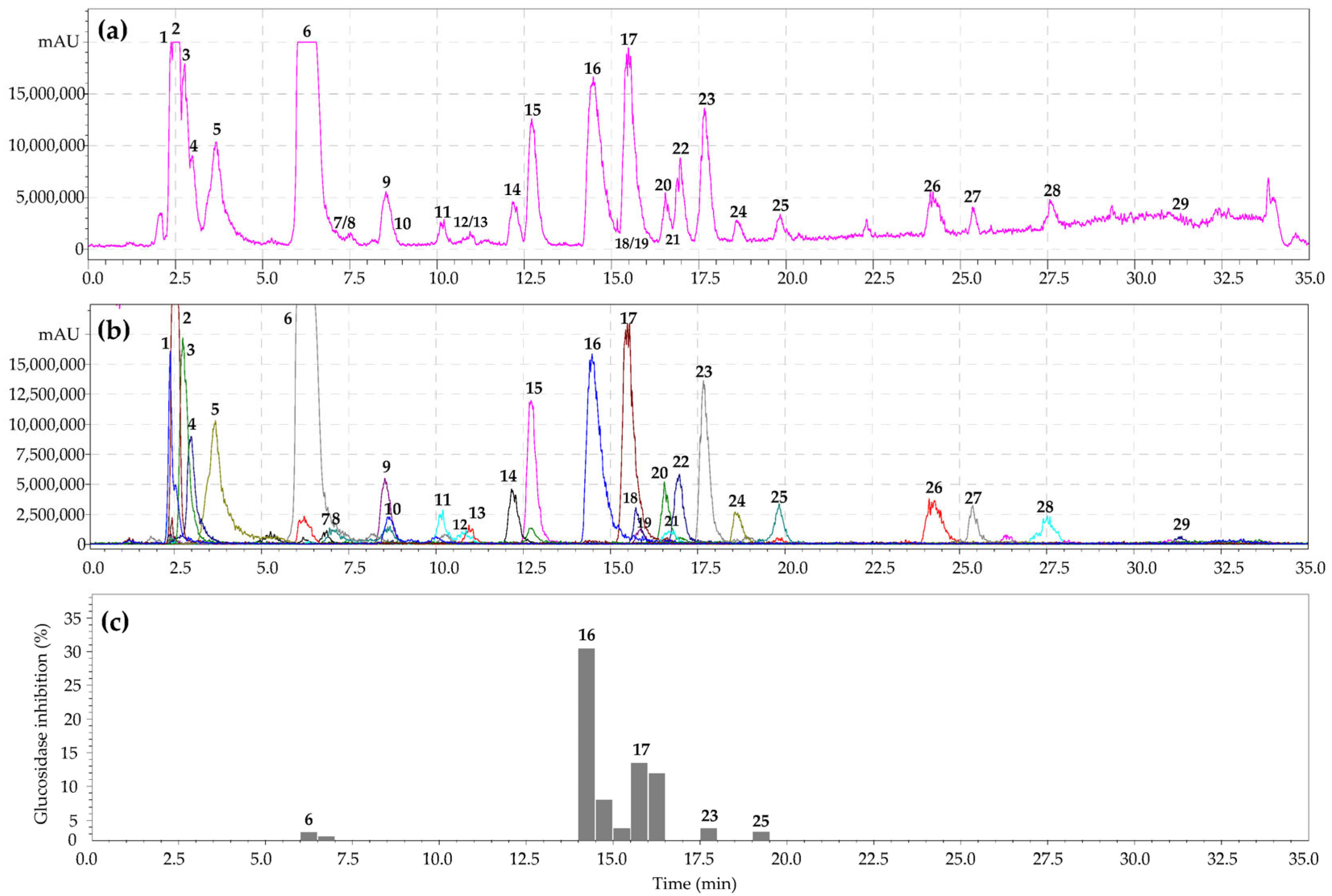
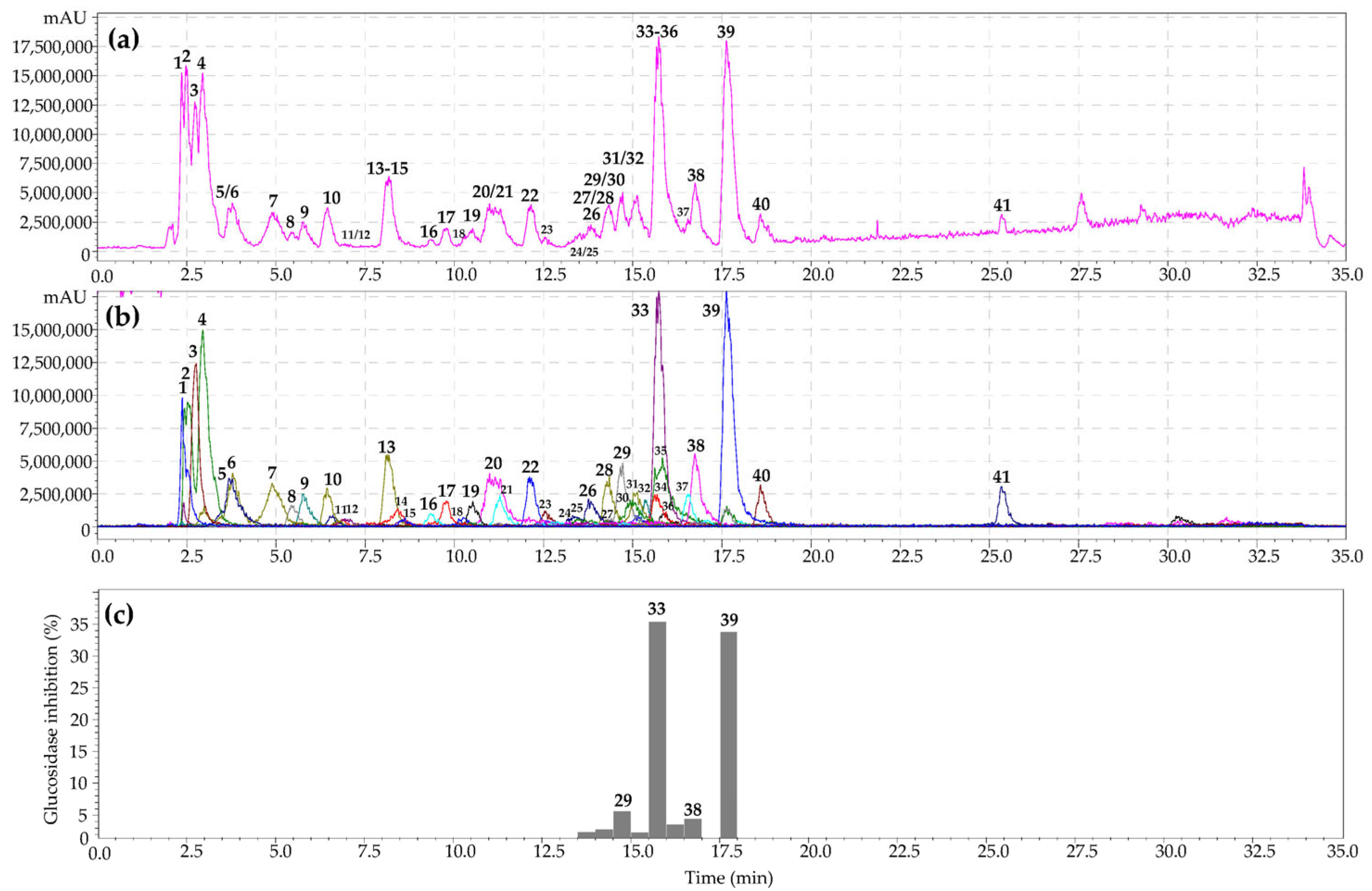
3.5. α-Glucosidase Inhibiting Activity of Geum aleppicum and Sibbaldianthe bifurca Herb Extract: HPLC Activity-Based Profiling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, J.S.; Nyberg, N.T.; Staerk, D. Assessment of constituents in Allium by multivariate data analysis, high-resolution α-glucosidase inhibition assay and HPLC-SPE-NMR. Food Chem. 2014, 161, 192–198. [Google Scholar] [CrossRef]
- Cardullo, N.; Muccilli, V.; Pulvirenti, L.; Cornu, A.; Pouységu, L.; Deffieux, D.; Quideau, S.; Tringali, C. C-glucosidic ellagitannins and galloylated glucoses as potential functional food ingredients with anti-diabetic properties: A study of α-glucosidase and α-amylase inhibition. Food Chem. 2020, 313, 126099. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Luo, A.; Zhong, K.; Huang, Y.; Gao, Y.; Zhang, J.; Gao, H.; Xu, Z.; Gao, X. α-Glucosidase inhibitory activity by the flower buds of Lonicera japonica Thunb. J. Func. Foods 2013, 5, 1253–1259. [Google Scholar] [CrossRef]
- Feng, J.; Yang, X.-W.; Wang, R.-F. Bio-assay guided isolation and identification of α-glucosidase inhibitors from the leaves of Aquilaria sinensis. Phytochemistry 2011, 72, 242–247. [Google Scholar] [CrossRef]
- Sudhir, R.; Mohan, V. Postprandial hyperglycemia in patients with type 2 diabetes mellitus. Treat. Endocrinol. 2002, 1, 105–116. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef]
- Kashtoh, H.; Baek, K.-H. Recent updates on phytoconstituent alpha-glucosidase inhibitors: An approach towards the treatment of type two diabetes. Plants 2022, 11, 2722. [Google Scholar] [CrossRef] [PubMed]
- Kashchenko, N.I.; Olennikov, D.N. Phenolome of Asian agrimony tea (Agrimonia asiatica Juz., Rosaceae): LC-MS profile, α-glucosidase inhibitory potential and stability. Foods 2020, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Potter, D.; Eriksson, T.; Evans, R.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Robertson, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Komarov, V.L. Flora of USSR; AN SSSR: Moscow, Russia, 1941; Volume X, pp. 81–254. [Google Scholar]
- Batorova, S.M.; Yakovlev, G.P.; Aseeva, T.A. Reference-Book of Traditional Tibetan Medicine Herbs; Nauka: Novosibirsk, Russia, 2013; pp. 67–73. [Google Scholar]
- Makarov, A.A. Plant Medical Remedies of Yakut Traditional Medicine; YaGU: Yakutsk, Russia, 1974; pp. 47–51. [Google Scholar]
- Tang, X.; Li, J.; Liu, L.; Jing, H.; Zuo, W.; Zeng, Y. Transcriptome analysis provides insights into Potentilla bifurca adaptation to high altitude. Life 2022, 12, 1337. [Google Scholar] [CrossRef]
- Liu, Y.; Xiang, S.; Fu, X. Characterization of the complete chloroplast genome sequence of medicinal plant: Potentilla bifurca (Rosaceae). Mitochondrial DNA B Resour. 2021, 6, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.R.; Jin, H.Z.; Qin, J.J.; Fu, J.J.; Zhang, W.D. Chemical constituents of plants from the genus Geum. Chem. Biodivers. 2011, 8, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Yoshida, T.; Hatano, T.; Iwasaki, M.; Kubo, M.; Orime, T.; Yoshizaki, M.; Naruhashi, N. Hydrolysable tannins as chemotaxonomic markers in the Rosaceae. Phytochemistry 1992, 31, 3091–3096. [Google Scholar] [CrossRef]
- Yang, W.; Zheng, M.S.; Lu, H.Z. 2013. Study the chemical constituents of Geum aleppicum Jacq. J. Med. Sci. Yanbian Univ. 2013, 36, 32–34. [Google Scholar]
- Piao, X.; Tian, Y.; Mi, X.; Cui, J. Tyrosinase inhibition of Potentilla bifurca. China J. Chin. Mater. Med. 2009, 34, 1952–1954. [Google Scholar]
- Wang, G.Y.; Yan, P.Y.; Liu, W.; Liu, L.K.; Li, J.P.; Zeng, Y. Potentilla bifurca flavonoids effectively improve insulin resistance. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 8358–8369. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Chirikova, N.K.; Olennikov, D.N. Agrimoniin, an active ellagitannin from Comarum palustre herb with anti-α-glucosidase and antidiabetic potential in streptozotocin-induced diabetic rats. Molecules 2017, 22, 73. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Olennikov, D.N.; Chirikova, N.K. Metabolites of Siberian raspberries: LC-MS profile, seasonal variation, antioxidant activity and, thermal stability of Rubus matsumuranus phenolome. Plants 2021, 10, 2317. [Google Scholar] [CrossRef]
- Olennikov, D.N. Ellagitannins and other phenolic compounds from Comarum palustre. Chem. Nat. Compd. 2016, 52, 721–723. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kruglova, M.Y. A new quercetin glucoside and other phenolic compounds from the genus Filipendula. Chem. Nat. Compd. 2013, 49, 610–616. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Kuz’mina, S.S. Phenolic profile of Potentilla anserina L. (Rosaceae) herb of Siberian origin and development of a rapid method for simultaneous determination of major phenolics in P. anserina pharmaceutical products by microcolumn RP-HPLC-UV. Molecules 2014, 20, 224–248. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Kashchenko, N.I. Componential profile and amylase inhibiting activity of phenolic compounds from Calendula officinalis L. leaves. Sci. World J. 2014, 2014, 654193. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Kashchenko, N.I. New flavonoids and turkesterone-2-O-cinnamate form leaves of Rhaponticum uniflorum. Chem. Nat. Compd 2019, 55, 256–264. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.-W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Zilfikarov, I.N.; Penzina, T.A. Use of microcolumn HPLC for analysis of aloenin in Aloe arborescens raw material and related drugs. Pharm. Chem. J. 2013, 47, 494–497. [Google Scholar] [CrossRef]
- Granica, S.; Kłębowska, A.; Kosiński, M.; Piwowarski, J.P.; Dudek, M.K.; Kaźmierski, S.; Kiss, A.K. Effects of Geum urbanum L. root extracts and its constituents on polymorphonuclear leucocytes functions. Significance in periodontal diseases. J. Ethnopharmacol. 2016, 188, 1–12. [Google Scholar] [CrossRef]
- Aubert, S.; Choler, P.; Pratt, J.; Douzet, R.; Gout, E.; Bligny, R. Methyl-β-d-glucopyranoside in higher plants: Accumulation and intracellular localization in Geum montanum L. leaves and in model systems studied by 13C nuclear magnetic resonance. J. Exp. Bot. 2004, 55, 2179–2189. [Google Scholar] [CrossRef]
- Ming, D.S.; Jiang, R.W.; But, P.P.; Towers, G.H.; Yu, D.Q. A new compound from Geum rivale L. J. Asian Nat. Prod. Res. 2002, 4, 217–220. [Google Scholar] [CrossRef]
- Shahani, S.; Gohari, A.R.; Monsef-Esfahani, H.R. Quantification of sucrose in the root of Geum iranicum Khatamsaz. Pharm. Biomed. Res. 2015, 1, 31–36. [Google Scholar] [CrossRef]
- Berkov, S.; Kasabova, N.; Pavlova, D.; Tonkov, S. Metabolic and chemotaxonomical studies in some Geum (Rosaceae) species. Phytol. Balc. 2017, 23, 7–16. [Google Scholar]
- Suh, S.J.; Cho, K.J.; Moon, T.C.; Chang, H.W.; Park, Y.G.; Kim, C.H. 3,4,5-trihydroxybenzaldehyde from Geum japonicum has dual inhibitory effect on matrix metalloproteinase 9; inhibition of gelatinoytic activity as well as MMP-9 expression in TNF-alpha induced HASMC. J. Cell Biochem. 2008, 105, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Yoshida, T.; Hatano, T. Correlation of oxidative transformation of hydrolysable tannins and plant evolution. Phytochemistry 2000, 55, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Panizzi, L.; Catalano, S.; Miarelli, C.; Cioni, P.L.; Campeol, E. In vitro antimicrobial activity of extracts and isolated constituents of Geum rivale. Phytother. Res. 2000, 14, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Gstirner, F.; Widenmann, H. Contents of the rhizome of Geum urbanum. Sci. Pharm. 1964, 32, 98–104. [Google Scholar]
- Yang, Z.; Yue, S.-J.; Gao, H.; Zhang, Q.; Xu, D.-Q.; Zhou, J.; Li, J.-J.; Tang, Y.-P. Natural deep eutectic solvent-ultrasound assisted extraction: A green approach for ellagic acid extraction from Geum japonicum. Front. Nutr. 2023, 9, 1079767. [Google Scholar] [CrossRef]
- Piwowarski, J.P.; Granica, S.; Kosiński, M.; Kiss, A.K. Secondary metabolites from roots of Geum urbanum L. Biochem. System. Ecol. 2014, 53, 46–50. [Google Scholar] [CrossRef]
- Yoshida, T.; Okuda, T.; Memon, M.U.; Shingu, T. Structure of gemin A, a new dimeric ellagitannin having α- and β-glucose cores. J. Chem. Soc. Chem. Commun. 1982, 6, 351–353. [Google Scholar] [CrossRef]
- Moilanen, J.; Salminen, J.-P. Ecologically neglected tannins and their biologically relevant activity: Chemical structures of plant ellagitannins reveal their in vitro oxidative activity at high pH. Chemoecology 2008, 18, 73–83. [Google Scholar] [CrossRef]
- Dong, H.; Chen, S.X.; Kini, R.M.; Xu, H.X. Effects of tannins from Geum japonicum on the catalytic activity of thrombin and factor Xa of blood coagulation cascade. J. Nat. Prod. 1998, 61, 1356–1360. [Google Scholar] [CrossRef]
- Kamińska, J.; Assenow, I. Phytochemical studies of Geum bulgaricum Panc. Acta Pol. Pharm. 1971, 28, 201–206. [Google Scholar]
- Murai, Y.; Iwashina, T. Flavonol glucuronides from Geum calthifolium var. nipponicum and Sieversia pentapetala (Rosaceae). Biochem. System. Ecol. 2010, 38, 1081–1082. [Google Scholar] [CrossRef]
- Xu, H.-X.; Kadota, S.; Wang, H.; Kurokawa, M.; Shiraki, K. A new hydrolyzable tannin from Geum japonicum and its antiviral activity. Heterocycles 1994, 38, 167–175. [Google Scholar]
- Zaharieva, M.M.; Dimitrova, L.L.; Philipov, S.; Nikolova, I.; Vilhelmova, N.; Grozdanov, P.; Nikolova, N.; Popova, M.; Bankova, V.; Konstantinov, S.M.; et al. In vitro antineoplastic and antiviral activity and in vivo toxicity of Geum urbanum L. extracts. Molecules 2022, 27, 245. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, S.; Kouno, I.; Kawano, N. Triterpenoids and glycosides from Geum japonicum. Phytochemistry 1985, 24, 115–118. [Google Scholar] [CrossRef]
- Dimitrova, L.; Zaharieva, M.M.; Popova, M.; Kostadinova, N.; Tsvetkova, I.; Bankova, V.; Najdenski, H. Antimicrobial and antioxidant potential of different solvent extracts of the medicinal plant Geum urbanum L. Chem. Cent. J. 2017, 11, 113. [Google Scholar] [CrossRef]
- Yean, M.-H.; Kim, J.-S.; Hyun, Y.-J.; Hyun, J.-W.; Bae, K.-H.; Kang, S.-S. Terpenoids and phenolics from Geum japonicum. Korean J. Pharmacogn. 2012, 43, 107–121. [Google Scholar]
- Xu, H.X.; Zeng, F.Q.; Wan, M.; Sim, K.Y. Anti-HIV triterpene acids from Geum japonicum. J. Nat. Prod. 1996, 59, 643–645. [Google Scholar] [CrossRef]
- Ton That, Q.; Nguyen Thien, T.V.; Dang, H.P.; Le Hoan, N.; Vo, L.K.T.; Nguyen, M.H.D.; Ngu, N.T.; Nguyen, T.S.; Hansen, P.E. Chemical constituents of Geum urbanum L. roots. Nat. Prod. Res. 2018, 32, 2529–2534. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Oligomeric hydrolyzable tannins, a new class of plant polyphenols. Heterocycles 1990, 30, 1195. [Google Scholar] [CrossRef]
- Moilanen, J.; Sinkkonen, J.; Salminen, J.P. Characterization of bioactive plant ellagitannins by chromatographic, spectroscopic and mass spectrometric methods. Chemoecology 2013, 23, 165–179. [Google Scholar] [CrossRef]
- Schulenburg, K.; Feller, A.; Hoffmann, T.; Schecker, J.H.; Martens, S.; Schwab, W. Formation of β-glucogallin, the precursor of ellagic acid in strawberry and raspberry. J. Exp. Bot. 2016, 67, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Zheng, W. Effect of plant growth temperature on antioxidant capacity in strawberry. J. Agric. Food Chem. 2001, 49, 4977–4982. [Google Scholar] [CrossRef] [PubMed]
- Grochowski, D.M.; Skalicka-Woźniak, K.; Orhan, I.E.; Xiao, J.; Locatelli, M.; Piwowarski, J.P.; Granica, S.; Tomczyk, M. A comprehensive review of agrimoniin. Ann. N. Y. Acad. Sci. 2017, 1401, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, A.; Salminen, J.-P. Hydrolyzable tannins, flavonol glycosides, and phenolic acids show seasonal and ontogenic variation in Geranium sylvaticum. J. Agric. Food Chem. 2017, 65, 6387–6403. [Google Scholar] [CrossRef]
- Feng, T.; Moore, M.J.; Yan, M.-H.; Sun, Y.-X.; Zhang, H.-J.; Meng, A.-P.; Li, X.-D.; Jian, S.-G.; Li, J.-Q.; Wang, H.-C. Phylogenetic study of the tribe Potentilleae (Rosaceae), with further insight into the disintegration of Sibbaldia. J. Syst. Evol. 2017, 55, 177–191. [Google Scholar] [CrossRef]
- Wilkes, S.; Glasl, H. Isolation, characterization, and systematic significance of 2-pyrone-4,6-dicarboxylic acid in Rosaceae. Phytochemistry 2001, 58, 441–449. [Google Scholar] [CrossRef]
- Okuda, T. Systematics and health effects of chemically distinct tannins in medicinal plants. Phytochemistry 2005, 66, 2012–2031. [Google Scholar] [CrossRef]
- Challice, J.S. Phenolic compounds of the subfamily Pomoideae: A chemotaxonomic survey. Phytochemistry 1972, 12, 1095–1101. [Google Scholar] [CrossRef]
- Hamburger, M. HPLC-based activity profiling for pharmacologically and toxicologically relevant natural products—Principles and recent examples. Pharm. Biol. 2019, 57, 328–334. [Google Scholar] [CrossRef]
- Mahmoud, A.B.; Danton, O.; Kaiser, M.; Khalid, S.; Hamburger, M.; Mäser, P. HPLC-based activity profiling for antiprotozoal compounds in Croton gratissimus and Cuscuta hyalina. Front. Pharmacol. 2020, 11, 1246. [Google Scholar] [CrossRef]
- Akaberi, M.; Danton, O.; Tayarani-Najaran, Z.; Asili, J.; Iranshahi, M.; Emami, S.A.; Hamburger, M. HPLC-based activity profiling for antiprotozoal compounds in the endemic Iranian medicinal plant Helichrysum oocephalum. J. Nat. Prod. 2019, 82, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Kongstad, K.T.; Özdemir, C.; Barzak, A.; Wubshet, S.G.; Staerk, D. Combined use of high-resolution α-glucosidase inhibition profiling and high-performance liquid chromatography-high-resolution mass spectrometry-solid-phase extraction-nuclear magnetic resonance spectroscopy for investigation of antidiabetic principles in crude plant extracts. J. Agric. Food Chem. 2015, 63, 2257–2263. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hu, M.; Hu, X.; Ding, H.; Gong, D.; Zhang, G. Inhibitory mechanism of epicatechin gallate on α-amylase and α-glucosidase and its combinational effect with acarbose or epigallocatechin gallate. J. Mol. Liq. 2019, 290, 111202. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, W.; Feng, F.; Zhang, Y.; Kang, W. α-Glucosidase inhibitors isolated from medicinal plants. Food Sci. Hum. Wellness 2014, 3, 136–174. [Google Scholar] [CrossRef]
- Toda, M.; Kawabata, J.; Kasai, T. Inhibitory effects of ellagi- and gallotannins on rat intestinal α-glucosidase complexes. Biosci. Biotechnol. Biochem. 2001, 65, 542–547. [Google Scholar] [CrossRef]
- Spencer, C.M.; Cai, Y.; Martin, R.; Gaffney, S.H.; Goulding, P.N.; Mangnolato, D.; Lilley, Y.; Haslam, E. Polyphenol complexation—Some thoughts and observatios. Phytochemistry 1988, 27, 2397–2409. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, G.; Liao, Y.; Gong, D. Inhibitory kinetics and mechanism of kaempferol on α-glucosidase. Food Chem. 2016, 190, 207–215. [Google Scholar] [CrossRef]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.T.; Sousa, J.L.C.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure-activity relationship study. J. Enzyme Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No | tr, min | Compound a | UV, nm | [M–H]−, MS/MS, m/z | Seasonal Content, mg/g DW ± SD | ||
|---|---|---|---|---|---|---|---|
| May | July | September | |||||
| 1 | 2.39 | Saccharose S | 205 | 341 [M–H]− | 2.39 ± 0.13 | 24.18 ± 1.23 | 41.68 ± 2.55 |
| 2 | 2.55 | Glucose S | 205 | 179 [M–H]− | 3.86 ± 0.23 | 48.52 ± 2.96 | 27.11 ± 1.63 |
| 3 | 2.78 | Malic acid S | 205 | 133 [M–H]− | 2.63 ± 0.14 | 5.69 ± 0.36 | 9.18 ± 0.53 |
| 4 | 3.04 | Citric acid S | 205 | 191 [M–H]− | 0.93 ± 0.05 | 3.11 ± 0.20 | 5.73 ± 0.36 |
| 5 | 3.66 | 2-Pyrone-4,6-dicarboxylic acid S | 314 | 183 [M–H]−; [183]→139 [(M–H)–CO2]−, 111 [(M–H)–COOH–CO]− | 0.83 ± 0.04 | 5.29 ± 0.32 | 5.33 ± 0.32 |
| 6 | 6.32 | 3,4-Dihydroxybenzoic acid 4-O-Glc S | 297 | 315 [M–H]−; [315]→153 [(M–H)–Glc]− | 2.44 ± 0.17 | 11.27 ± 0.62 | 10.86 ± 0.68 |
| 7 | 6.72 | 3,4,5-Trihydroxybenzaldehyde S | 275 | 153 [M–H]− | traces | 0.08 ± 0.01 | traces |
| 8 | 7.04 | Pedunculagin S | 270 | 783 [M–H]−; 391 [M–2H]2− | traces | 0.26 ± 0.02 | traces |
| 9 | 8.50 | Benzoic acid O-Hex L | 265 | 283 [M–H]−; [283]→121 [(M–H)–Hex]− | 0.83 ± 0.05 | 4.33 ± 0.28 | 4.08 ± 0.22 |
| 10 | 8.59 | Casuariin S | 270 | 783 [M–H]−; 391 [M–2H]2− | 1.26 ± 0.08 | 2.57 ± 0.15 | 2.03 ± 0.12 |
| 11 | 10.16 | Ellagic acid methyl ether O-Pent L | 254, 360 | 447 [M–H]−; [447]→315 [(M–H)–Pent]−; [315]→301 [(M–H)–Pent–CH2]− | traces | 0.54 ± 0.03 | 0.10 ± 0.01 |
| 12 | 10.82 | Ellagic acid methyl ether O-Pent L | 254, 360 | 447 [M–H]−; [447]→315 [(M–H)–Pent]−; [315]→301 [(M–H)–Pent–CH2]− | traces | 0.32 ± 0.02 | traces |
| 13 | 10.96 | Quercetin-O-HexA-O-HexA L | 254, 267, 351 | 653 [M–H]−; [653]→477 [(M–H)–HexA]−, 301 [(M–H)–2×HexA]− | traces | 0.82 ± 0.05 | traces |
| 14 | 12.29 | Kaempferol-O-HexA-O-HexA L | 265, 345 | 637 [M–H]−; [637]→461 [(M–H)–HexA]−, 285 [(M–H)–2×HexA]− | 0.26 ± 0.01 | 1.89 ± 0.13 | 0.29 ± 0.02 |
| 15 | 12.73 | Feruloyl tartronic acid L | 296, 327 | 295 [M–H]−; [295]→193 [(M–H)–102]− | 1.07 ± 0.07 | 3.97 ± 0.25 | 2.18 ± 0.13 |
| 16 | 14.47 | Gemin A S | 270 | 1871 [M–H]−; 935 [M–2H]2− | 10.18 ± 0.70 | 53.26 ± 3.25 | 42.11 ± 2.11 |
| 17 | 15.49 | Quercetin-3-O-GlcA S | 254, 268, 352 | 477 [M–H]−; [477]→301 [(M–H)–GlcA]− | 5.20 ± 0.30 | 26.83 ± 1.58 | 10.75 ± 0.59 |
| 18 | 15.84 | Quercetin-3-O-Glc S | 254, 285, 355 | 463 [M–H]−; [463]→301 [(M–H)–Glc]− | traces | 1.28 ± 0.07 | traces |
| 19 | 15.88 | Ellagic acid S | 255, 367 | 301 [M–H]− | traces | 0.89 ± 0.05 | 5.63 ± 0.38 |
| 20 | 16.55 | Kaempferol-3-O-GlcA S | 265, 344 | 461 [M–H]−; [461]→285 [(M–H)–GlcA]− | 1.03 ± 0.06 | 2.97 ± 0.18 | 2.08 ± 0.11 |
| 21 | 16.66 | Kaempferol-3-O-Glc S | 265, 343 | 447 [M–H]−; [447]→285 [(M–H)–Glc]− | traces | 0.52 ± 0.03 | traces |
| 22 | 16.92 | Niga-ichigoside F1 isomer L | 210 | 665 [M–H]−; [665]→503 [(M–H)–Glc]− | 1.22 ± 0.07 | 2.01 ± 0.12 | 2.09 ± 0.13 |
| 23 | 17.69 | Niga-ichigoside F1 S | 210 | 665 [M–H]−; [665]→503 [(M–H)–Glc]− | 4.27 ± 0.23 | 10.82 ± 0.66 | 8.12 ± 0.44 |
| 24 | 18.62 | Gallocatechin gallate O-gallate L | 250 | 593 [M–H]−; [593]→441 [(M–H)–GallA]− | traces | 2.93 ± 0.17 | 1.14 ± 0.06 |
| 25 | 19.84 | Rosamultin (tormentic acid O-Glc) S | 210 | 649 [M–H]−; [649]→487 [(M–H)–Glc]− | 0.74 ± 0.05 | 1.73 ± 0.10 | 1.93 ± 0.11 |
| 26 | 24.27 | Tormentic acid S | 210 | 487 [M–H]− | 0.52 ± 0.04 | 1.14 ± 0.07 | 1.37 ± 0.08 |
| 27 | 25.42 | Ellagic acid methyl ether L | 254, 362 | 315 [M–H]−; [315]→301 [(M–H)–CH2]− | 0.14 ± 0.01 | 0.63 ± 0.04 | 1.83 ± 0.11 |
| 28 | 27.50 | Corosolic acid S | 210 | 471 [M–H]− | 0.10 ± 0.01 | 0.43 ± 0.02 | 0.69 ± 0.04 |
| 29 | 31.31 | Ursolic acid S | 210 | 455 [M–H]− | traces | 0.05 ± 0.00 | 0.67 ± 0.04 |
| No | tr, min | Compound a | UV, nm | [M–H]−, MS/MS, m/z | Seasonal Content, mg/g DW ± SD | ||
|---|---|---|---|---|---|---|---|
| May | July | September | |||||
| 1 | 2.39 | Saccharose S | 205 | 341 [M–H]− | 1.86 ± 0.10 | 2.14 ± 0.12 | 26.18 ± 1.57 |
| 2 | 2.55 | Glucose S | 205 | 179 [M–H]− | 3.96 ± 0.22 | 6.29 ± 0.42 | 3.35 ± 0.17 |
| 3 | 2.78 | Malic acid S | 205 | 133 [M–H]− | 2.52 ± 0.15 | 4.27 ± 0.30 | 5.83 ± 0.36 |
| 4 | 3.04 | Citric acid S | 205 | 191 [M–H]− | 2.69 ± 0.18 | 4.83 ± 0.30 | 8.59 ± 0.50 |
| 5 | 3.66 | 2-Pyrone-4,6-dicarboxylic acid S | 314 | 183 [M–H]−; [183]→139 [(M–H)–CO2]−, 111 [(M–H)–COOH–CO]− | 0.50 ± 0.03 | 1.29 ± 0.07 | 1.14 ± 0.06 |
| 6 | 3.72 | Monogalloyl hexose L | 268 | 331 [M–H]−; [331]→169 [(M–H)–Hex]− | 2.93 ± 0.16 | 1.54 ± 0.11 | 3.22 ± 0.19 |
| 7 | 4.93 | Monogalloyl hexose L | 268 | 331 [M–H]−; [331]→169 [(M–H)–Hex]− | 2.97 ± 0.18 | 2.83 ± 0.17 | 4.69 ± 0.28 |
| 8 | 5.42 | Gallic acid S | 272 | 169 [M–H]− | 0.29 ± 0.02 | 0.82 ± 0.06 | 0.80 ± 0.05 |
| 9 | 6.32 | 3,4-Dihydroxybenzoic acid 4-O-Glc S | 273 | 343 [M–H]−; [343]→181 [(M–H)–Glc]− | 0.92 ± 0.06 | 1.63 ± 0.11 | 1.29 ± 0.08 |
| 10 | 6.64 | p-Hydroxybenzoic acid O-hexoside isomer L | 274 | 299 [M–H]−; [299]→137 [(M–H)–Glc]− | 0.83 ± 0.05 | 1.52 ± 0.09 | 1.16 ± 0.06 |
| 11 | 6.85 | Digalloyl hexose L | 272 | 483 [M–H]−; [483]→331 [(M–H)–GalA]− | traces | traces | traces |
| 12 | 7.04 | Pedunculagin S | 270 | 783 [M–H]−; 391 [M–2H]2− | traces | traces | traces |
| 13 | 8.14 | 1-O-p-Hydroxybenzoic acid O-Glc S | 274 | 299 [M–H]−; [299]→137 [(M–H)–Glc]− | 1.16 ± 0.07 | 2.32 ± 0.15 | 2.09 ± 0.13 |
| 14 | 8.41 | Digalloyl hexose L | 272 | 483 [M–H]−; [483]→331 [(M–H)–GalA]− | 0.42 ± 0.02 | 0.29 ± 0.02 | traces |
| 15 | 8.59 | Casuariin S | 270 | 783 [M–H]−; 391 [M–2H]2− | traces | traces | traces |
| 16 | 9.34 | Tellimagrandin I1 S | 272 | 785 [M–H]−; 392 [M–2H]2−; 1571 [2M–H]− | traces | 0.53 ± 0.03 | traces |
| 17 | 9.77 | Digalloyl hexose L | 272 | 483 [M–H]−; [483]→331 [(M–H)–GalA]− | 0.51 ± 0.03 | 0.39 ± 0.03 | 0.10 ± 0.01 |
| 18 | 10.26 | Trigalloyl hexose L | 274 | 635 [M–H]−; [635]→483 [(M–H)–GalA]− | 0.59 ± 0.04 | 0.42 ± 0.02 | 0.24 ± 0.01 |
| 19 | 10.51 | Trigalloyl hexose L | 274 | 635 [M–H]−; [635]→483 [(M–H)–GalA]− | 0.78 ± 0.04 | 0.58 ± 0.04 | 0.43 ± 0.02 |
| 20 | 10.95 | Trigalloyl hexose L | 274 | 635 [M–H]−; [635]→483 [(M–H)–GalA]− | 2.58 ± 0.16 | 2.14 ± 0.14 | 2.03 ± 0.12 |
| 21 | 11.28 | Tellimagrandin I2 S | 272 | 785 [M–H]−; 392 [M–2H]2−; 1571 [2M–H]− | 0.52 ± 0.03 | 1.16 ± 0.07 | 1.10 ± 0.07 |
| 22 | 12.16 | Trigalloyl hexose L | 274 | 635 [M–H]−; [635]→483 [(M–H)–Hex]− | 3.56 ± 0.21 | 2.89 ± 0.20 | 2.72 ± 0.18 |
| 23 | 12.55 | Trigalloyl-HHDP-Hex L | 273 | 951 [M–H]−; 475 [M–2H]2− | 0.37 ± 0.02 | 0.26 ± 0.1 | traces |
| 24 | 13.38 | Rugosin E1 S | 272 | 1721 [M–H]−; 860 [M–2H]2− | traces | 0.50 ± 0.03 | traces |
| 25 | 13.47 | Casuarinin S | 272 | 935 [M–H]−; 467 [M–2H]2− | traces | 0.93 ± 0.06 | traces |
| 26 | 13.82 | Rugosin E2 S | 272 | 1721 [M–H]−; 860 [M–2H]2− | traces | traces | traces |
| 27 | 14.12 | Potentillin S | 272 | 935 [M–H]−; 467 [M–2H]2− | traces | 0.09 ± 0.01 | traces |
| 28 | 14.31 | Casuarictin S | 276 | 937 [M–H]−; 468 [M–2H]2− | 1.60 ± 0.09 | 5.28 ± 0.31 | 4.16 ± 0.28 |
| 29 | 14.66 | Tetragalloyl hexose L | 271 | 787 [M–H]−; [787]→635 [(M–H)–GalA]− | 3.14 ± 0.16 | 2.83 ± 0.15 | 0.83 ± 0.05 |
| 30 | 14.94 | Agrimoniin S | 271 | 935 [M–H]−; 467 [M–2H]2− | traces | 0.26 ± 0.02 | traces |
| 31 | 15.07 | Tellimagrandin II2 S | 276 | 937 [M–H]−; 468 [M–2H]2− | 3.62 ± 0.21 | 7.83 ± 0.51 | 5.16 ± 0.33 |
| 32 | 15.38 | Kaempferol-3-O-Hex-Hex L | 265, 344 | 609 [M–H]−; [609]→447 [(M–H)–Glc]−, 285 [(M–H)–2×Glc]− | 0.14 ± 0.01 | 0.52 ± 0.03 | 0.50 ± 0.03 |
| 33 | 15.49 | Quercetin-3-O-GlcA S | 254, 268, 352 | 477 [M–H]−; [477]→301 [(M–H)–GlcA]− | 7.88 ± 0.49 | 19.62 ± 1.20 | 12.84 ± 0.73 |
| 34 | 15.67 | Pentagalloyl hexose L | 272 | 939 [M–H]−; [939]→787 [(M–H)–GalA]− | 0.84 ± 0.05 | 0.52 ± 0.03 | traces |
| 35 | 15.88 | Ellagic acid S | 255, 367 | 301 [M–H]− | traces | 1.26 ± 0.08 | 3.18 ± 0.19 |
| 36 | 15.93 | Digalloyl-bis-HHDP-Hex L | 271 | 1087 [M–H]−; 543 [M–2H]2− | 0.17 ± 0.01 | 0.10 ± 0.01 | traces |
| 37 | 16.55 | Kaempferol-3-O-GlcA S | 265, 344 | 461 [M–H]−; [461]→285 [(M–H)–GlcA]− | traces | 3.22 ± 0.19 | 3.35 ± 0.20 |
| 38 | 16.76 | Quercetin-7-O-Glc S | 254, 268, 364 | 463 [M–H]−; [463]→301 [(M–H)–Glc]− | 0.26 ± 0.01 | 4.29 ± 0.26 | 4.35 ± 0.29 |
| 39 | 17.64 | Quercetin-3-O-Ara S | 253, 268, 365 | 433 [M–H]−; [433]→301 [(M–H)–Ara]− | 5.63 ± 0.33 | 21.59 ± 1.45 | 18.26 ± 1.06 |
| 40 | 18.59 | Quercetin-3-O-(6″-O-Cin)-Glc S | 270, 285, 364 | 593 [M–H]−; [593]→463 [(M–H)–Cin]−, 301 [(M–H)–Glc]− | 0.08 ± 0.01 | 1.53 ± 0.10 | 0.16 ± 0.01 |
| 41 | 25.42 | Ellagic acid methyl ether L | 254, 362 | 315 [M–H]−; [315]→301 [(M–H)–CH2]− | 0.18 ± 0.01 | 2.53 ± 0.15 | 3.19 ± 0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashchenko, N.I.; Olennikov, D.N.; Chirikova, N.K. Metabolites of Geum aleppicum and Sibbaldianthe bifurca: Diversity and α-Glucosidase Inhibitory Potential. Metabolites 2023, 13, 689. https://doi.org/10.3390/metabo13060689
Kashchenko NI, Olennikov DN, Chirikova NK. Metabolites of Geum aleppicum and Sibbaldianthe bifurca: Diversity and α-Glucosidase Inhibitory Potential. Metabolites. 2023; 13(6):689. https://doi.org/10.3390/metabo13060689
Chicago/Turabian StyleKashchenko, Nina I., Daniil N. Olennikov, and Nadezhda K. Chirikova. 2023. "Metabolites of Geum aleppicum and Sibbaldianthe bifurca: Diversity and α-Glucosidase Inhibitory Potential" Metabolites 13, no. 6: 689. https://doi.org/10.3390/metabo13060689