
Bioactive Metabolite Survey of Actinobacteria Showing Plant Growth Promoting Traits to Develop Novel Biofertilizers
 , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culturing Conditions
2.2. Estimation of PGP Traits and Abiotic Stress Tolerance
2.2.1. Indolic Compound Production
2.2.2. Organic and Inorganic Phosphate Solubilization
2.2.3. Growth under Drought and Salt Stress
2.2.4. Growth in Nitrogen-Free Medium
2.3. Metabolite Extraction and HPLC/MS/Q-TOF Analysis
2.4. In Vivo Evaluation of Plant Growth Promotion by PGP Actinobacteria
2.5. Effect of S. violaceoruber Culture Seed-Priming Treatment on Germination
2.6. Identification of VOCs Using SPME-GC/MS
2.7. Quantification of Global DNA Methylation
3. Results
3.1. Looking for Multiple PGP Traits of Three Selected Actinobacteria
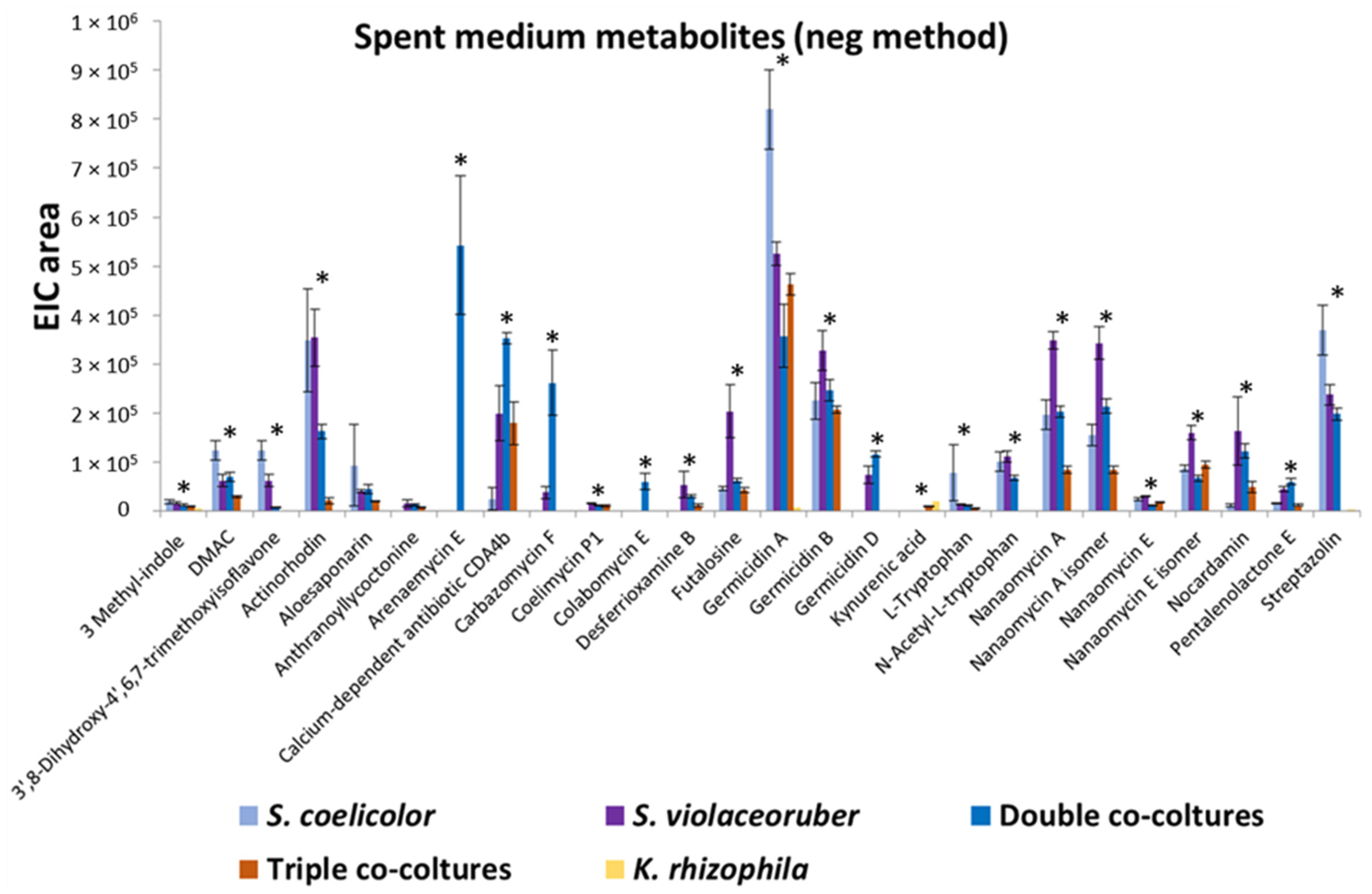
3.2. Bacterial Metabolomic Analyses of Single and Mixed Actinobacterial Cultures
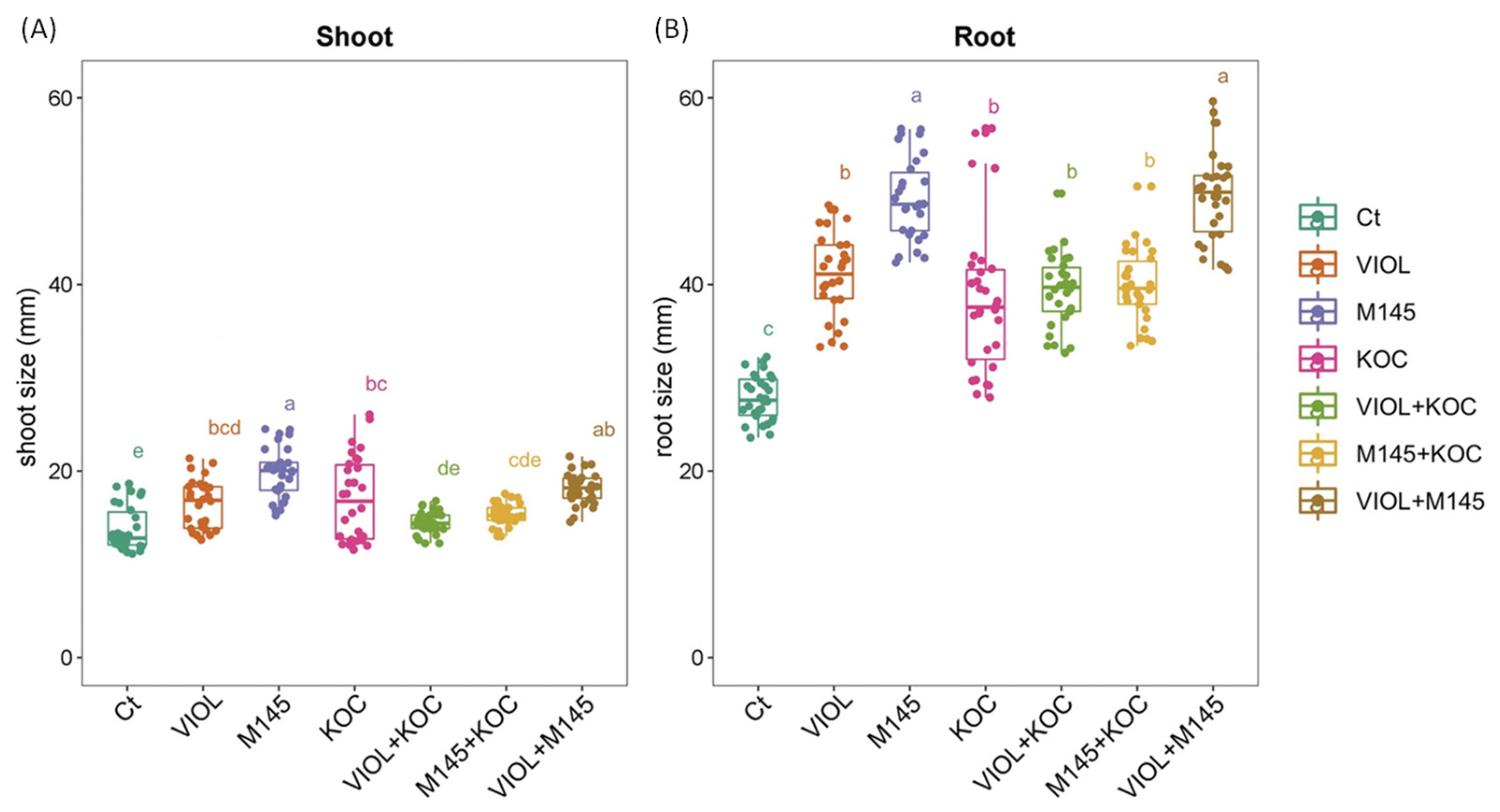
3.3. Biostimulant Effects on S. lycopersicum Seedlings from PGP Actinobacteria Treated Seeds
3.4. Effect of PGP Seed-Priming Treatment on Germination
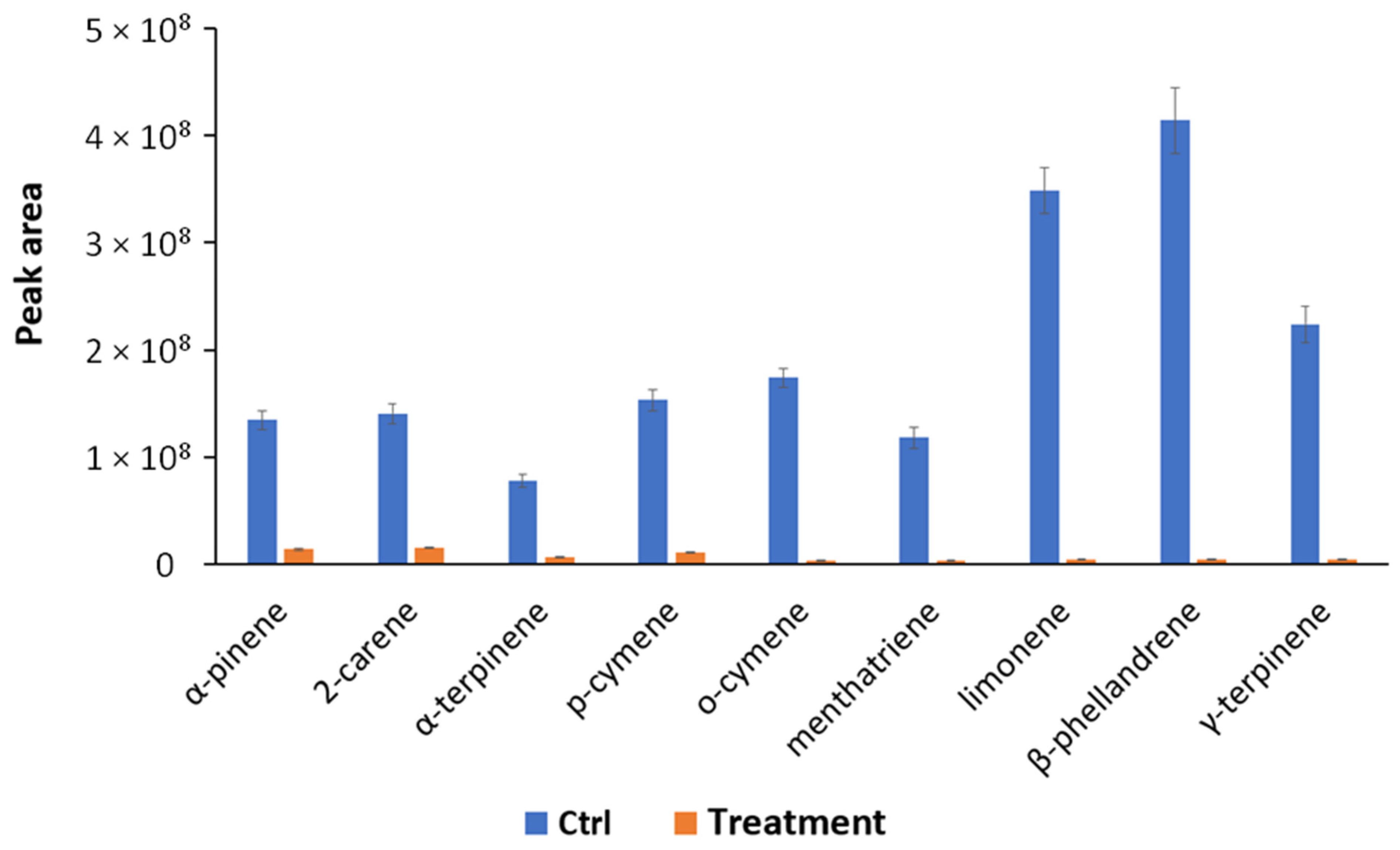
3.5. Volatile Organic Compounds Produced by S. violaceoruber and S. lycopersicum
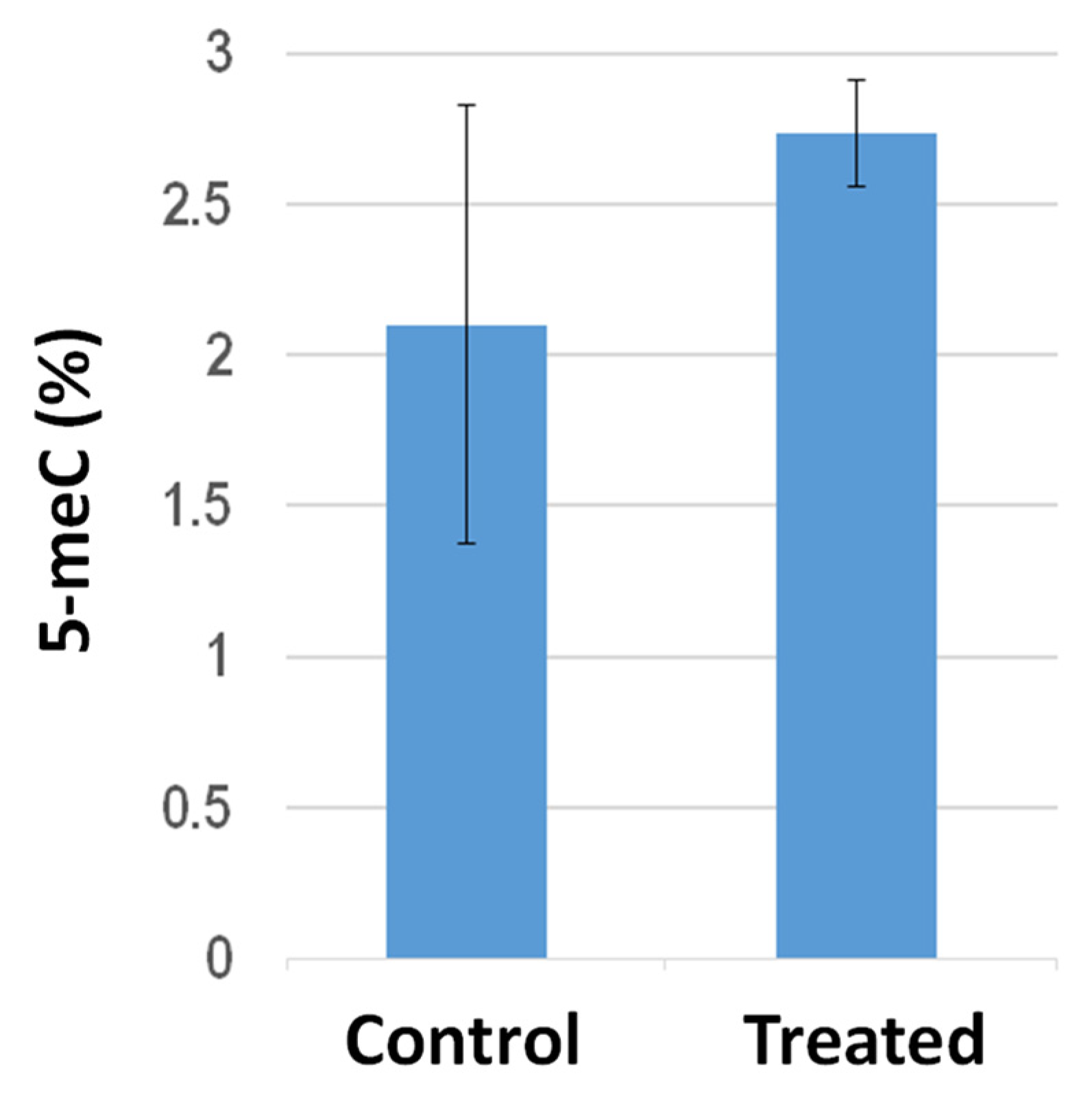
3.6. Effect of S. violaceoruber Cultivation on Global DNA Methylation Amount of S. lycopersicum Shoots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baweja, P.; Kumar, S.; Kumar, G. Fertilizers and Pesticides: Their Impact on Soil Health and Environment BT-Soil Health; Giri, B., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 265–285. ISBN 978-3-030-44364-1. [Google Scholar] [CrossRef]
- Hatfield, J.L. Climate Change: Challenges for Future Crop Adjustments. In Climate Change and Plant Abiotic Stress Tolerance; Tuteja, N., Gill, S.S., Eds.; Wiley-VCH: Hoboken, NJ, USA, 2013; pp. 1–26. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A Potential Approach for Sustainable Agriculture Development. Environ. Sci. Pollut. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers Function as Key Player in Sustainable Agriculture by Improving Soil Fertility, Plant Tolerance and Crop Productivity. Microb. Cell Fact. 2014, 13, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.-S.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The Use of Microbial Inoculants for Biological Control, Plant Growth Promotion, and Sustainable Agriculture: A Review. Eur. J. Plant Pathol. 2022, 162, 759–792. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Doolotkelvieva, T.; Bobusheva, S.; Konurbaeva, M. Effects of Streptomyces Biofertilizer to Soil Fertility and Rhizosphere’s Functional Biodiversity of Agricultural Plants. Adv. Microbiol. 2015, 5, 555–571. [Google Scholar] [CrossRef] [Green Version]
- Afridi, M.S.; Van Hamme, J.d.; Bundschuh, J.; Sumaira; Khan, M.N.; Salam, A.; Waqar, M.; Munis, M.F.H.; Chaudhary, H.J. Biotechnological Approaches in Agriculture and Environmental Management-Bacterium Kocuria Rhizophila 14ASP as Heavy Metal and Salt- Tolerant Plant Growth- Promoting Strain. Biologia 2021, 76, 3091–3105. [Google Scholar] [CrossRef]
- Riaz, U.; Murtaza, G.; Anum, W.; Samreen, T.; Sarfraz, M.; Nazir, M.Z. Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides BT-Microbiota and Biofertilizers: A Sustainable Continuum for Plant and Soil Health; Hakeem, K.R., Dar, G.H., Mehmood, M.A., Bhat, R.A., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 181–196. [Google Scholar] [CrossRef]
- Kaari, M.; Manikkam, R.; Annamalai, K.K.; Joseph, J. Actinobacteria as a Source of Biofertilizer/Biocontrol Agents for Bio-Organic Agriculture. J. Appl. Microbiol. 2022, 134, lxac047. [Google Scholar] [CrossRef]
- Marchawala, F.Z.; Amin, S.R. Elucidating IAA Producing Kocuria flava FA10 as a Potent PGPB and Biocontrol Agent. Int. J. Biotechnol. Biochem. 2018, 14, 185–191. [Google Scholar]
- Mansurova, M.; Ebert, B.E.; Blank, L.M.; Ibáñez, A.J. A Breath of Information: The Volatilome. Curr. Genet. 2018, 64, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F. Practical Streptomyces Genetics, 2nd ed.; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Glickmann, E.; Dessaux, Y. A Critical Examination of the Specificity of the Salkowski Reagent for Indolic Compounds Produced by Phytopathogenic Bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischler, D.; Mascher, T.; Claessen, D.; López-García, M.T.; Yagüe, P.; González-Quiñónez, N.; Rioseras, B.; Manteca, A. The SCO4117 ECF Sigma Factor Pleiotropically Controls Secondary Metabolism and Morphogenesis in Streptomyces coelicolor. Front. Microbiol. 2018, 9, 312. [Google Scholar] [CrossRef]
- Faddetta, T.; Abbate, L.; Alibrandi, P.; Arancio, W.; Siino, D.; Strati, F.; De Filippo, C.; Fatta Del Bosco, S.; Carimi, F.; Puglia, A.M.; et al. The Endophytic Microbiota of Citrus Limon Is Transmitted from Seed to Shoot Highlighting Differences of Bacterial and Fungal Community Structures. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S. An Efficient Microbiological Growth Medium for Screening Phosphate Solubilizing Microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Matthews, S.; Suhaimi, M. Selection of Suitable Growth Medium for Free-Living Diazotrophs Isolated from Compost. J. Trop. Agric. Food Sci. 2010, 38, 211–219. [Google Scholar]
- Emanuele, S.; Notaro, A.; Piccionello, A.P.; Maggio, A.; Lauricella, M.; D’Anneo, A.; Cernigliaro, C.; Calvaruso, G.; Giuliano, M. Sicilian Litchi Fruit Extracts Induce Autophagy versus Apoptosis Switch in Human Colon Cancer Cells. Nutrients 2018, 10, 1490. [Google Scholar] [CrossRef] [Green Version]
- Raimondo, S.; Urzì, O.; Meraviglia, S.; Di Simone, M.; Corsale, A.M.; Rabienezhad Ganji, N.; Palumbo Piccionello, A.; Polito, G.; Lo Presti, E.; Dieli, F.; et al. Anti-Inflammatory Properties of Lemon-Derived Extracellular Vesicles Are Achieved through the Inhibition of ERK/NF-ΚB Signalling Pathways. J. Cell. Mol. Med. 2022, 26, 4195–4209. [Google Scholar] [CrossRef]
- METLIN. Available online: https://metlin.scripps.edu/landing_page.php?pgcontent=mainPage (accessed on 23 February 2022).
- ANOVA Calculator: One-Way Analysis of Variance Calculator. Available online: https://goodcalculators.com/one-way-anova-calculator/ (accessed on 1 February 2023).
- R-Packages. Available online: http://www.r-project.org (accessed on 1 February 2023).
- Campobenedetto, C.; Grange, E.; Mannino, G.; Van Arkel, J.; Beekwilder, J.; Karlova, R.; Garabello, C.; Contartese, V.; Bertea, C.M. A Biostimulant Seed Treatment Improved Heat Stress Tolerance during Cucumber Seed Germination by Acting on the Antioxidant System and Glyoxylate Cycle. Front. Plant Sci. 2020, 11, 836. [Google Scholar] [CrossRef]
- Polito, G.; Semenzato, G.; Del Duca, S.; Castronovo, L.M.; Vassallo, A.; Chioccioli, S.; Borsetti, D.; Calabretta, V.; Puglia, A.M.; Fani, R.; et al. Endophytic Bacteria and Essential Oil from Origanum vulgare ssp. Vulgare Share Some VOCs with an Antibacterial Activity. Microorganisms 2022, 10, 1424. [Google Scholar] [CrossRef]
- Boubekri, K.; Soumare, A.; Mardad, I.; Lyamlouli, K.; Ouhdouch, Y.; Hafidi, M.; Kouisni, L. Multifunctional Role of Actinobacteria in Agricultural Production Sustainability: A Review. Microbiol. Res. 2022, 261, 127059. [Google Scholar] [CrossRef] [PubMed]
- Thilagam, R.; Hemalatha, N. Plant Growth Promotion and Chilli Anthracnose Disease Suppression Ability of Rhizosphere Soil Actinobacteria. J. Appl. Microbiol. 2019, 126, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Kol, S.; Elena Merlo, M.; Scheltema, R.A.; De Vries, M.; Vonk, R.J.; Kikkert, N.A.; Dijkhuizen, L.; Breitling, R.; Takano, E. Metabolomic Characterization of the Salt Stress Response in Streptomyces coelicolor. Appl. Environ. Microbiol. 2010, 76, 2574–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, S.D.; Chater, K.F.; Cerdeñ O-Tá Rraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete Genome Sequence of the Model Actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef]
- Faddetta, T.; Renzone, G.; Vassallo, A.; Rimini, E.; Nasillo, G.; Buscarino, G.; Agnello, S.; Licciardi, M.; Botta, L.; Scaloni, A.; et al. Streptomyces coelicolor Vesicles: Many Molecules to Be Delivered. Appl. Environ. Microbiol. 2022, 88, e01881-21. [Google Scholar] [CrossRef]
- Nodwell, J.R. Microbe Profile: Streptomyces coelicolor: A Burlesque of Pigments and Phenotypes. Microbiology 2019, 165, 953–955. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The Antimicrobial Volatile Power of the Rhizospheric Isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [Green Version]
- Ross, Z.M.; O’Gara, E.A.; Hill, D.J.; Sleightholme, H.V.; Maslin, D.J. Antimicrobial Properties of Garlic Oil against Human Enteric Bacteria: Evaluation of Methodologies and Comparisons with Garlic Oil Sulfides and Garlic Powder. Appl. Environ. Microbiol. 2001, 67, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Becher, P.G.; Verschut, V.; Bibb, M.J.; Bush, M.J.; Molnár, B.P.; Barane, E.; Al-Bassam, M.M.; Chandra, G.; Song, L.; Challis, G.L. Developmentally Regulated Volatiles Geosmin and 2-Methylisoborneol Attract a Soil Arthropod to Streptomyces Bacteria Promoting Spore Dispersal. Nat. Microbiol. 2020, 5, 821–829. [Google Scholar] [CrossRef]
- Zhou, F.; Pichersky, E. The Complete Functional Characterisation of the Terpene Synthase Family in Tomato. New Phytol. 2020, 226, 1341–1360. [Google Scholar] [CrossRef] [Green Version]
- Merzaeva, O.V.; Shirokikh, I.G. Colonization of Plant Rhizosphere by Actinomycetes of Different Genera. Microbiology 2006, 75, 226–230. [Google Scholar] [CrossRef]
- Bonaldi, M.; Chen, X.; Kunova, A.; Pizzatti, C.; Saracchi, M.; Cortesi, P. Colonization of Lettuce Rhizosphere and Roots by Tagged Streptomyces. Front. Microbiol. 2015, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Kour, D.; Rana, K.L.; Kumar, V.; Dhaliwa, S.; Verma, P.; Singh, B.; Chauahan, V.S.; Sugitha, T.C.K.; Saxena, A.K. Plant Microbiomes and Its Beneficial Multifunctional Plant Growth Promoting Attributes. Int. J. Environ. Sci. Nat. Resour. 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant Growth Promoting Rhizobacteria as Biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Sadeghi, A.; Karimi, E.; Dahaji, P.A.; Javid, M.G.; Dalvand, Y.; Askari, H. Plant Growth Promoting Activity of an Auxin and Siderophore Producing Isolate of Streptomyces under Saline Soil Conditions. World J. Microbiol. Biotechnol. 2012, 28, 1503–1509. [Google Scholar] [CrossRef]
- Sellstedt, A.; Richau, K.H. Aspects of Nitrogen-Fixing Actinobacteria, in Particular Free-Living and Symbiotic Frankia. FEMS Microbiol. Lett. 2013, 342, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Sinha, N. Tomato (Solanum lycopersicum): A Model Fruit-Bearing Crop. Cold Spring Harb. Protoc. 2008, 2008, pdb-emo105. [Google Scholar] [CrossRef]
- Pannacci, E.; Baratta, S.; Falcinelli, B.; Farneselli, M.; Tei, F. Mugwort (Artemisia vulgaris L.) Aqueous Extract: Hormesis and Biostimulant Activity for Seed Germination and Seedling Growth in Vegetable Crops. Agriculture 2022, 12, 1329. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-Acetic Acid in Plant–Microbe Interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Bernabeu-Meana, M.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S. Effect of Plant Growth-Promoting Rhizobacteria on Salicornia ramosissima Seed Germination under Salinity, CO2 and Temperature Stress. Agronomy 2019, 9, 655. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Yang, S.H.; Palaniyandi, S.A.; Han, J.S.; Yoon, T.M.; Kim, T.J.; Suh, J.W. Azalomycin F Complex Is an Antifungal Substance Produced by Streptomyces malaysiensis MJM1968 Isolated from Agricultural Soil. J. Appl. Biol. Chem. 2010, 53, 545–552. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Qiao, X.; Li, Z.; Li, F.; Chen, M.; Wang, Y.; Huang, Y.; Cui, H. Antifungal Activity of Volatile Organic Compounds from Streptomyces alboflavus TD-1. FEMS Microbiol. Lett. 2013, 341, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-Growth Promotion and Biocontrol Properties of Three Streptomyces spp. Isolates to Control Bacterial Rice Pathogens. Front. Microbiol. 2019, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Tarabily, K.A.; AlKhajeh, A.S.; Ayyash, M.M.; Alnuaimi, L.H.; Sham, A.; ElBaghdady, K.Z.; Tariq, S.; AbuQamar, S.F. Growth Promotion of Salicornia bigelovii by Micromonospora chalcea UAE1, an Endophytic 1-Aminocyclopropane-1-Carboxylic Acid Deaminase-Producing Actinobacterial Isolate. Front. Microbiol. 2019, 10, 1694. [Google Scholar] [CrossRef] [PubMed]
- Mathew, B.T.; Torky, Y.; Amin, A.; Mourad, A.H.I.; Ayyash, M.M.; El-Keblawy, A.; Hilal-Alnaqbi, A.; AbuQamar, S.F.; El-Tarabily, K.A. Halotolerant Marine Rhizosphere-Competent Actinobacteria Promote Salicornia bigelovii Growth and Seed Production Using Seawater Irrigation. Front. Microbiol. 2020, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Le, K.D.; Kim, J.; Nguyen, H.T.; Yu, N.H.; Park, A.R.; Lee, C.W.; Kim, J.C. Streptomyces Sp. JCK-6131 Protects Plants Against Bacterial and Fungal Diseases via Two Mechanisms. Front. Plant Sci. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam Palaniyandi, S.; Hwan Yang, S.; Zhang, L.; Suh, J.-W. Effects of Actinobacteria on Plant Disease Suppression and Growth Promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef]
- Al Raish, S.M.; Saeed, E.E.; Alyafei, D.M.; El-Tarabily, K.A.; AbuQamar, S.F. Evaluation of Streptomycete Actinobacterial Isolates as Biocontrol Agents against Royal Poinciana Stem Canker Disease Caused by the Fungal Pathogen Neoscytalidium Dimidiatum. Biol. Control 2021, 164, 104783. [Google Scholar] [CrossRef]
- Aoki, Y.; Matsumoto, D.; Kawaide, H.; Natsume, M. Physiological Role of Germicidins in Spore Germination and Hyphal Elongation in Streptomyces coelicolor A3(2). J. Antibiot. 2011, 64, 607–611. [Google Scholar] [CrossRef]
- Powell, P.E.; Szaniszlo, P.J.; Cline, G.R.; Reid, C.P.P. Hydroxamate Siderophores in the Iron Nutrition of Plants. J. Plant Nutr. 1982, 5, 653–673. [Google Scholar] [CrossRef]
- Smits, T.H.M.; Duffy, B. Genomics of Iron Acquisition in the Plant Pathogen Erwinia Amylovora: Insights in the Biosynthetic Pathway of the Siderophore Desferrioxamine E. Arch. Microbiol. 2011, 193, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Vassallo, A.; Palazzotto, E.; Renzone, G.; Botta, L.; Faddetta, T.; Scaloni, A.; Puglia, A.M.; Gallo, G. The Streptomyces coelicolor Small ORF TrpM Stimulates Growth and Morphological Development and Exerts Opposite Effects on Actinorhodin and Calcium-Dependent Antibiotic Production. Front. Microbiol. 2020, 11, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelou, P.-K.; Kokotou, M.G.; Constantinou-Kokotou, V. Identification of Auxin Metabolites in Brassicaceae by Ultra-Performance Liquid Chromatography Coupled with High-Resolution Mass Spectrometry. Molecules 2019, 24, 2615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Brumos, J.; Li, H.; Ji, Y.; Ke, M.; Gong, X.; Zeng, Q.; Li, W.; Zhang, X.; An, F.; et al. A Small-Molecule Screen Identifies L-Kynurenine as a Competitive Inhibitor of TAA1/TAR Activity in Ethylene-Directed Auxin Biosynthesis and Root Growth in Arabidopsis. Plant Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [Green Version]
- Balahbib, A.; El Omari, N.; Hachlafi, N.E.L.; Lakhdar, F.; El Menyiy, N.; Salhi, N.; Mrabti, H.N.; Bakrim, S.; Zengin, G.; Bouyahya, A. Health Beneficial and Pharmacological Properties of P-Cymene. Food Chem. Toxicol. 2021, 153, 112259. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential Oils of Oregano: Biological Activity beyond Their Antimicrobial Properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Oyedemi, S.O.; Okoh, A.I.; Mabinya, L.V.; Pirochenva, G.; Afolayan, A.J. The proposed mechanism of bactericidal action of eugenol, α-terpineol and γ-terpinene against Listeria monocytogenes, Streptococcus pyogenes, Proteus vulgaris and Escherichia coli. Afr. J. Biotechnol. 2009, 8, 1280–1286. [Google Scholar]
- Le, K.D.; Yu, N.H.; Park, A.R.; Park, D.J.; Kim, C.J.; Kim, J.C. Streptomyces Sp. AN090126 as a Biocontrol Agent against Bacterial and Fungal Plant Diseases. Microorganisms 2022, 10, 791. [Google Scholar] [CrossRef]
- Sudha, A.; Durgadevi, D.; Archana, S.; Muthukumar, A.; Suthin Raj, T.; Nakkeeran, S.; Poczai, P.; Nasif, O.; Ansari, M.J.; Sayyed, R.Z. Unraveling the Tripartite Interaction of Volatile Compounds of Streptomyces rochei with Grain Mold Pathogens Infecting Sorghum. Front. Microbiol. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Cavalieri, V. Chapter 3—Model Organisms and Their Application in Environmental Epigenetics. In Translational Epigenetics; Fry, R., Ed.; Elsevier, Academic Press: Cambridge, MA, USA, 2020; Volume 22, pp. 67–87. ISBN 25425358. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | IAA Production | Growth Using FePO4 | Growth Using Ca3P2O8 | Growth Using AlPO4 | Growth Using Fitate | Growth Using N2 | Growth under Drought Stress | Growth under Saline Stress |
|---|---|---|---|---|---|---|---|---|
| S. coelicolor | + | g+ | g+ | g+ | g+ | N.A. | + | N.D. |
| S. violaceoruber | + | g+ | g+ | g+ | g+ | + | + | + |
| K. rhizophila | + | g+ | g+ | g+ | g+ | N.A. | + | + |
| tR (min) | Compounds | Molecular Formula | ESI− [M − H]− (m/z) Exp. | ESI+ [M + H]+ (m/z) Exp. | Classes | Occurrence 1 | |
|---|---|---|---|---|---|---|---|
| 1.22 | 3 Methyl-indole a | C9H9N | 130.0869 | - | Tryptophan metabolism | 1–7 neg | - |
| 2.41 | L-Tryptophan a | C11H12N2O2 | 203.0822 | 205.0980 | Amino Acid | 1–6 neg | 1–6 pos |
| 2.69 | Kynurenic acid | C10H7NO3 | 188.0332 | 190.0509 | Tryptophan metabolism | 5–7 neg | 5–7 pos |
| 3.41 | Frenolicin E | C18H20O8 | - | 365.1190 | Polyketide | - | 2–6 pos |
| 3.68 | Frenolicin E isomer | C18H20O8 | - | 365.1190 | Polyketide | - | 2–6 pos |
| 4.20 | Coelimycin P1 | C17H20N2O4S | 347.1033 | 349.1251 | Alkaloid | 2–5 neg | 2–6 pos |
| 4.27 | Desferrioxamine B | C25H48N6O8 | 559.3428 | 561.3649 | Siderophore | 2–6 neg | 2–6 pos |
| 4.62 | Carbazomycin F | C16H15NO4 | 284.0938 | - | Alkaloid | 2–4, 6 neg | - |
| 4.71 | Futalosine | C19H18N4O7 | 413.1090 | 415.1260 | Inosine | 1–6 neg | 1–6 pos |
| 4.72 | Streptophenazine F | C25H30N2O5 | - | 439.2264 | Phenazine | - | 3,4,6 pos |
| 4.82 | Pimprinethine a | C13H12N2O | - | 213.1034 | Alkaloid | 1–7 pos | |
| 4.84 | Anthranoyllycoctonine | C32H46N2O8 | 585.3142 | 587.332 | Diterpenoid | 2–6 neg | 2–6 pos |
| 5.07 | Nocardamin | C27H48N6O9 | 599.3336 | 601.3565 | Siderophore | 1–6 neg | 1–6 pos |
| 5.15 | Streptazolin a | C11H13NO3 | 206.0820 | - | Alkaloid antibiotic | 1–4,6,7 neg | - |
| 5.30 | Nanaomycin E | C16H14O7 | 317.0659 | 319.0833 | Polyketide | 1–6 neg | 1,2,3,5 pos |
| 5.46 | N-Acetyl-L-tryptophan | C13H14N2O3 | 245.0922 | - | Tryptophan metabolism | 1–4, 6 neg | - |
| 5.47 | Nanaomycin A a | C16H14O6 | 301.0707 | 303.0880 | Polyketide | 1–6 neg | 1–6 pos |
| 5.48 | Nanaomycin E isomer | C16H14O8 | 317.0658 | - | Polyketide | 1–6 neg | - |
| 5.56 | Streptophenazine A | C24H28N2O5 | - | 425.2081 | Phenazine | - | 2–6 pos |
| 5.69 | Arenaemycin E a | C15H16O5 | 275.0906 | 277.1078 | Sesquiterpene lactone | 3–6 neg | 3–6 pos |
| 5.87 | 3′,8-Dihydroxy-4′,6,7-trimethoxyisoflavone | C18H16O7 | 343.0700 | - | Isoflavone | 1–4, 6 neg | - |
| 6.20 | Nanaomycin A isomer | C16H14O6 | 301.0709 | 303.0880 | Polyketide | 1–6 neg | 1–6 pos |
| 6.40 | Germicidin B a | C10H14O3 | 181.0866 | 183.1027 | Pyranone Polyketide antibiotic | 1–6 neg | 1–6 pos |
| 6.55 | Pentalenolactone E | C15H18O4 | 261.1118 | - | Sesquiterpene lactone | 1–6 neg | - |
| 7.15 | Germicidin A a | C11H16O3 | 195.1027 | 197.1184 | Pyranone Polyketide antibiotic | 1–7 neg | 1–7 pos |
| 7.20 | Calcium-dependent antibiotic CDA4b | C67H80N14O26 | 1495.5206 | 1497.5536 | Polypeptide antibiotic | 1–6 neg | 2–6 pos |
| 7.72 | Colabomycin E | C32H32N2O7 | 555.2174 | - | Manumycin | 3,4,6 neg | - |
| 7.85 | Germicidin D a | C11H16O4 | 211.0962 | - | Pyranone Polyketide antibiotic | 2–4,6 neg | - |
| 7.98 | 3,8-Dihydroxy-1- methylanthraquinone-2-carboxylic acid (DMAC) a | C16H10O6 | 297.039 | - | Anthracene polyketide | 1–6 neg | - |
| 9.39 | Aloesaponarin a | C15H10O4 | 253.0501 | - | Anthracene (polyketide) | 1–6 neg | - |
| 9.56 | Actinorhodin a | C32H22O14 | 629.0917 | 631.1143 | Polyketide antibiotic | 1–6 neg | 1–6 pos |
| 10.21 | Streptorubin B a | C25H33N3O | - | 392.2730 | Prodiginine antibiotic | - | 1–6 pos |
| 11.12 | Undecylprodigiosin a | C25H35N3O | 392.2692 | 394.2881 | Prodiginine antibiotic | 1–6 neg | 1–6 pos |
| Thesis 1 | Tissue | Mean Value 2 | Standard Deviation | Q3 3 | Growth Rate (%) 4 |
|---|---|---|---|---|---|
| Ct | Root | 27.81 | 2.43 | 29.82 | - |
| Shoot | 13.80 | 2.38 | 15.61 | - | |
| VIOL | Root | 41.16 | 4.51 | 44.25 | 47.97 |
| Shoot | 16.41 | 2.62 | 18.32 | 18.90 | |
| M145 | Root | 49.24 | 4.35 | 52.01 | 77.04 |
| Shoot | 19.79 | 2.71 | 20.92 | 43.44 | |
| KOC | Root | 38.39 | 7.97 | 41.58 | 38.03 |
| Shoot | 16.92 | 4.50 | 20.66 | 22.61 | |
| VIOL + KOC | Root | 39.40 | 3.91 | 41.81 | 41.65 |
| Shoot | 14.49 | 1.18 | 15.28 | 5.05 | |
| M145 + KOC | Root | 39.87 | 3.88 | 42.48 | 43.33 |
| Shoot | 15.36 | 1.19 | 16.06 | 11.35 | |
| VIOL + M145 | Root | 49.58 | 4.90 | 51.68 | 78.24 |
| Shoot | 18.14 | 1.66 | 19.22 | 31.45 |
| Treatments * | Germination Index % # | Root Lenght (cm) # | Hypocotyl Lenght (cm) # |
|---|---|---|---|
| T1 | 4.75 ᵃ ᵇ | 1.11 ᵃ ᵇ | 1.01 ᵃ ᵇ |
| T2 | 5.11 ᵃ | 1.41 ᵃ | 1.18 ᵃ |
| CTRL | 4.61 ᵇ | 1.06 ᵇ | 0.89 ᵇ |
| tR (min) | Compounds | Area % |
|---|---|---|
| 7.75 | Disulfide, dimethyl | 76.89 |
| 20.09 | Dimethyl trisulfide | 1.09 |
| 25.60 | Hexanoic acid, 2-ethyl-, methyl ester | 14.74 |
| 36.41 | 2-Methylisoborneol | 0.35 |
| 39.24 | 1H-Indene, 1-ethylideneoctahydro-7a-methyl | 0.89 |
| 39.49 | 1H-Indene, 1-ethylideneoctahydro-7a-methyl-, isomer | 0.41 |
| 51.52 | Geosmin | 4.02 |
| 53.69 | Cadinene | 1.61 |
| tR (min) | Compounds | % Remaining on Treated Leaves vs. Ctrl |
|---|---|---|
| 15.9 | α-pinene | 10.39 |
| 16.7 | 2-carene | 10.93 |
| 17.4 | α-terpinene | 9.17 |
| 21.4 | p-cymene | 7.17 |
| 21.9 | o-cymene | 2.33 |
| 22.5 | menthatriene | 3.34 |
| 23.9 | limonene | 1.45 |
| 24.1 | β-phellandrene | 1.09 |
| 24.5 | γ-terpinene | 1.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faddetta, T.; Polito, G.; Abbate, L.; Alibrandi, P.; Zerbo, M.; Caldiero, C.; Reina, C.; Puccio, G.; Vaccaro, E.; Abenavoli, M.R.; et al. Bioactive Metabolite Survey of Actinobacteria Showing Plant Growth Promoting Traits to Develop Novel Biofertilizers. Metabolites 2023, 13, 374. https://doi.org/10.3390/metabo13030374
Faddetta T, Polito G, Abbate L, Alibrandi P, Zerbo M, Caldiero C, Reina C, Puccio G, Vaccaro E, Abenavoli MR, et al. Bioactive Metabolite Survey of Actinobacteria Showing Plant Growth Promoting Traits to Develop Novel Biofertilizers. Metabolites. 2023; 13(3):374. https://doi.org/10.3390/metabo13030374
Chicago/Turabian StyleFaddetta, Teresa, Giulia Polito, Loredana Abbate, Pasquale Alibrandi, Marcello Zerbo, Ciro Caldiero, Chiara Reina, Guglielmo Puccio, Edoardo Vaccaro, Maria Rosa Abenavoli, and et al. 2023. "Bioactive Metabolite Survey of Actinobacteria Showing Plant Growth Promoting Traits to Develop Novel Biofertilizers" Metabolites 13, no. 3: 374. https://doi.org/10.3390/metabo13030374