
Diet Quality and Liver Health in People Living with HIV in the MASH Cohort: A Multi-Omic Analysis of the Fecal Microbiome and Metabolome
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Demographics
3.2. Diet Quality in PLWH
3.3. Diet Quality and FIB-4
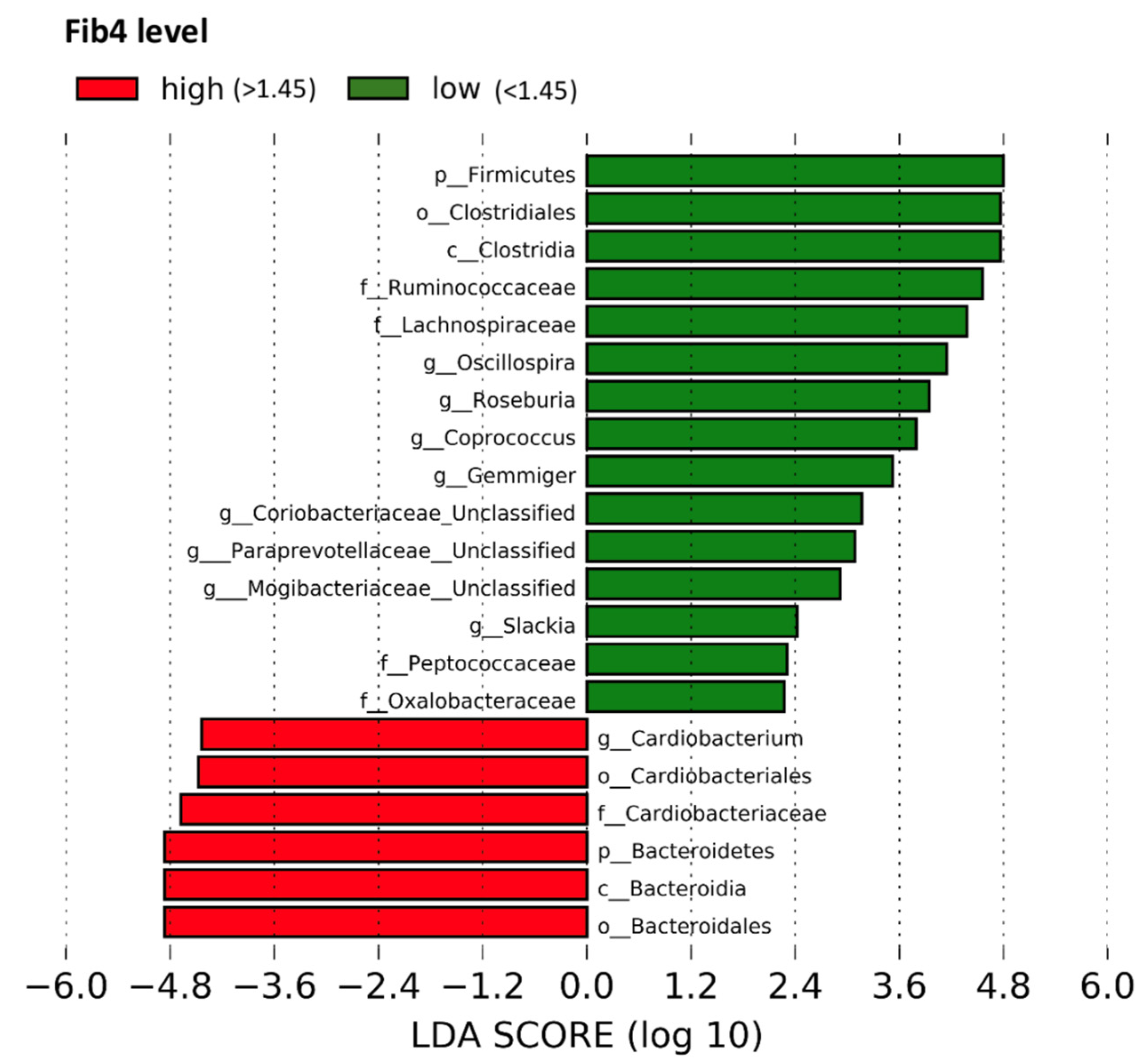
3.4. Fecal Microbial Taxa by FIB-4 Score
3.5. Microbial Metabolites by FIB-4 Score
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| d.f. | Sum of Squares | Mean Square | F | R2 | p-Value | |
|---|---|---|---|---|---|---|
| FIB-4 ≥ 1.45 1 | 1 | 0.3375 | 0.3375 | 0.9446 | 0.0192 | 0.671 |
Appendix B

References
- May, M.T.; Gompels, M.; Delpech, V.; Porter, K.; Orkin, C.; Kegg, S.; Hay, P.; Johnson, M.; Palfreeman, A.; Gilson, R.; et al. Impact on life expectancy of HIV-1 positive individuals of CD4+ cell count and viral load response to antiretroviral therapy. Aids 2014, 28, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Farahani, M.; Mulinder, H.; Farahani, A.; Marlink, R. Prevalence and distribution of non-AIDS causes of death among HIV-infected individuals receiving antiretroviral therapy: A systematic review and meta-analysis. Int. J. STD AIDS 2017, 28, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Croxford, S.; Kitching, A.; Desai, S.; Kall, M.; Edelstein, M.; Skingsley, A.; Burns, F.; Copas, A.; Brown, A.E.; Sullivan, A.K.; et al. Mortality and causes of death in people diagnosed with HIV in the era of highly active antiretroviral therapy compared with the general population: An analysis of a national observational cohort. Lancet Public Health 2017, 2, e35–e46. [Google Scholar] [CrossRef]
- Kaspar, M.B.; Sterling, R.K. Mechanisms of liver disease in patients infected with HIV. BMJ Open Gastroenterol. 2017, 4, e000166. [Google Scholar] [CrossRef]
- Sherman, K.E.; Rockstroh, J.; Thomas, D. Human immunodeficiency virus and liver disease: An update. Hepatology 2015, 62, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, J.A.; Sherman, K.E.; Sékaly, R.P.; Bordi, R.; Schlatzer, D.; Lai, S.; Khalsa, J.H.; Mandler, R.N.; Ehman, R.L.; Baum, M.K. Cocaethylene, simultaneous alcohol and cocaine use, and liver fibrosis in people living with and without HIV. Drug Alcohol Depend. 2022, 232, 109273. [Google Scholar] [CrossRef] [PubMed]
- Iacob, S.; Iacob, D.G. Infectious threats, the intestinal barrier, and its Trojan Horse: Dysbiosis. Front. Microbiol. 2019, 10, 1676. [Google Scholar] [CrossRef]
- Kim, C.J.; McKinnon, L.R.; Kovacs, C.; Kandel, G.; Huibner, S.; Chege, D.; Shahabi, K.; Benko, E.; Loutfy, M.; Ostrowski, M.; et al. Mucosal Th17 cell function is altered during HIV infection and is an independent predictor of systemic immune activation. J. Immunol. 2013, 191, 2164–2173. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Villar, S.; Rojo, D.; Martínez-Martínez, M.; Deusch, S.; Vázquez-Castellanos, J.F.; Bargiela, R.; Sainz, T.; Vera, M.; Moreno, S.; Estrada, V.; et al. Gut bacteria metabolism impacts immune recovery in HIV-infected individuals. EBioMedicine 2016, 8, 203–216. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, F.; Lu, H.; Wang, B.; Lei, D.; Wang, Y.; Zhu, B.; Li, L. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 2011, 54, 562–572. [Google Scholar] [CrossRef]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut microbiome-based metagenomic signature for non-invasive detection of advanced fibrosis in human nonalcoholic fatty liver disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef]
- Lang, S.; Farowski, F.; Martin, A.; Wisplinghoff, H.; Vehreschild, M.J.G.T.; Krawczyk, M.; Nowag, A.; Kretzschmar, A.; Scholz, C.; Kasper, P.; et al. Prediction of advanced fibrosis in non-alcoholic fatty liver disease using gut microbiota-based approaches compared with simple non-invasive tools. Sci. Rep. 2020, 10, 9385. [Google Scholar] [CrossRef]
- Humphreys, C. Intestinal Permeability. In Textbook of Natural Medicine, 5th ed.; Elsevier: Churchill Livingstone, London, UK, 2020; pp. 166–177.e4. [Google Scholar] [CrossRef]
- Luukkonen, P.K.; Sädevirta, S.; Zhou, Y.; Kayser, B.; Ali, A.; Ahonen, L.; Lallukka, S.; Pelloux, V.; Gaggini, M.; Jian, C.; et al. Saturated fat is more metabolically harmful for the human liver than unsaturated fat or simple sugars. Diabetes Care 2018, 41, 1732–1739. [Google Scholar] [CrossRef] [PubMed]
- Zelber-Sagi, S.; Ivancovsky-Wajcman, D.; Fliss Isakov, N.; Webb, M.; Orenstein, D.; Shibolet, O.; Kariv, R. High red and processed meat consumption is associated with non-alcoholic fatty liver disease and insulin resistance. J. Hepatol. 2018, 68, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.-f.; Shao, X.-f.; Li, J.-m.; Li, S.; Li, K.-l.; Li, D. Fruit and vegetable intake and liver cancer risk: A meta-analysis of prospective cohort studies. Food Funct. 2019, 10, 4478–4485. [Google Scholar] [CrossRef]
- Ross, A.B.; Godin, J.P.; Minehira, K.; Kirwan, J.P. Increasing whole grain intake as part of prevention and treatment of nonalcoholic Fatty liver disease. Int. J. Endocrinol. 2013, 2013, 585876. [Google Scholar] [CrossRef]
- Kratz, M.; Marcovina, S.; Nelson, J.E.; Yeh, M.M.; Kowdley, K.V.; Callahan, H.S.; Song, X.; Di, C.; Utzschneider, K.M. Dairy fat intake is associated with glucose tolerance, hepatic and systemic insulin sensitivity, and liver fat but not β-cell function in humans. Am. J. Clin. Nutr. 2014, 99, 1385–1396. [Google Scholar] [CrossRef]
- Kennedy, O.J.; Fallowfield, J.A.; Poole, R.; Hayes, P.C.; Parkes, J.; Roderick, P.J. All coffee types decrease the risk of adverse clinical outcomes in chronic liver disease: A UK Biobank study. BMC Public Health 2021, 21, 970. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yanagi, K.; Cheng, C.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Interactions between gut microbiota and non-alcoholic liver disease: The role of microbiota-derived metabolites. Pharmacol. Res. 2019, 141, 521–529. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Sobhonslidsuk, A.; Chanprasertyothin, S.; Pongrujikorn, T.; Kaewduang, P.; Promson, K.; Petraksa, S.; Ongphiphadhanakul, B. The association of gut microbiota with nonalcoholic steatohepatitis in Thais. BioMed. Res. Int. 2018, 2018, 9340316. [Google Scholar] [CrossRef]
- Weiss, J.J.; Sanchez, L.; Hubbard, J.; Lo, J.; Grinspoon, S.K.; Fitch, K.V. Diet quality is low and differs by sex in people with HIV. J. Nutr. 2019, 149, 78–87. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Xu, C.; Krampe, N.; Dillon, S.M.; Sette, P.; Falwell, E.; Haret-Richter, G.S.; Butterfield, T.; Dunsmore, T.L.; McFadden, W.M.; et al. High-fat diet exacerbates SIV pathogenesis and accelerates disease progression. J. Clin. Investig. 2019, 129, 5474–5488. [Google Scholar] [CrossRef]
- Moshfegh, A.J.; Rhodes, D.G.; Baer, D.J.; Murayi, T.; Clemens, J.C.; Rumpler, W.V.; Paul, D.R.; Sebastian, R.S.; Kuczynski, K.J.; Ingwersen, L.A.; et al. The US Department of Agriculture Automated Multiple-Pass Method reduces bias in the collection of energy intakes. Am. J. Clin. Nutr. 2008, 88, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Reedy, J.; Lerman, J.L.; Krebs-Smith, S.M.; Kirkpatrick, S.I.; Pannucci, T.E.; Wilson, M.M.; Subar, A.F.; Kahle, L.L.; Tooze, J.A. Evaluation of the Healthy Eating Index-2015. J. Acad. Nutr. Diet. 2018, 118, 1622–1633. [Google Scholar] [CrossRef]
- United States Department of Agriculture: Food and Nutrition Service: Center for Nutrition Policy and Promotion (CNPP). How the HEI Is Scored. Available online: https://www.fns.usda.gov/how-hei-scored (accessed on 30 December 2020).
- Chen, Z.; Hui, P.C.; Hui, M.; Yeoh, Y.K.; Wong, P.Y.; Chan, M.C.W.; Wong, M.C.S.; Ng, S.C.; Chan, F.K.L.; Chan, P.K.S. Impact of preservation method and 16S rRNA hypervariable region on gut microbiota profiling. mSystems 2019, 4, e00271-18. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A. The functional gastrointestinal disorders and the Rome III process. Gastroenterology 2006, 130, 1377–1390. [Google Scholar] [CrossRef]
- Sterling, R.K.; Lissen, E.; Clumeck, N.; Sola, R.; Correa, M.C.; Montaner, J.; Sulkowski, M.S.; Torriani, F.J.; Dieterich, D.T.; Thomas, D.L.; et al. Development of a simple noninvasive index to predict significant fibrosis in patients with HIV/HCV coinfection. Hepatology 2006, 43, 1317–1325. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Armitage, E.G.; Godzien, J.; Alonso-Herranz, V.; López-Gonzálvez, Á.; Barbas, C. Missing value imputation strategies for metabolomics data. Electrophoresis 2015, 36, 3050–3060. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; von Heydebreck, A.; Sültmann, H.; Poustka, A.; Vingron, M. Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 2002, 18 (Suppl. S1), S96–S104. [Google Scholar] [CrossRef]
- Mock, A.; Warta, R.; Dettling, S.; Brors, B.; Jäger, D.; Herold-Mende, C. MetaboDiff: An R package for differential metabolomic analysis. Bioinformatics 2018, 34, 3417–3418. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Perez, D.; Guan, H.; Madhivanan, P.; Mathee, K.; Narasimhan, G. So you think you can PLS-DA? BMC Bioinform. 2020, 21 (Suppl. S1), 2. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture: Food and Nutrition Service: Center for Nutrition Policy and Promotion: National Center for Health Statistics: What We Eat in America/National Health and Nutrition Examination Survey, 2013–2014. Healthy Eating Index-2015, Scores. Available online: https://www.fns.usda.gov/healthy-eating-index-hei (accessed on 30 December 2020).
- Volpe, G.E.; Ward, H.; Mwamburi, M.; Dinh, D.; Bhalchandra, S.; Wanke, C.; Kane, A.V. Associations of cocaine use and HIV infection with the intestinal microbiota, microbial translocation, and inflammation. J. Stud. Alcohol Drugs 2014, 75, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, J.A.; Sherman, K.E.; Campa, A.; Martinez, S.S.; Li, T.; Hernandez, J.; Teeman, C.; Mandler, R.N.; Chen, J.; Ehman, R.L.; et al. Food insecurity is associated with magnetic resonance-determined nonalcoholic fatty liver and liver fibrosis in low-income, middle-aged adults with and without HIV. Am. J. Clin. Nutr. 2021, 113, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H. Role of gut dysbiosis in liver diseases: What have we learned so far? Diseases 2019, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Weitkunat, K.; Stuhlmann, C.; Postel, A.; Rumberger, S.; Fankhänel, M.; Woting, A.; Petzke, K.J.; Gohlke, S.; Schulz, T.J.; Blaut, M.; et al. Short-chain fatty acids and inulin, but not guar gum, prevent diet-induced obesity and insulin resistance through differential mechanisms in mice. Sci. Rep. 2017, 7, 6109. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Heuman, D.M.; Hylemon, P.B.; Sanyal, A.J.; White, M.B.; Monteith, P.; Noble, N.A.; Unser, A.B.; Daita, K.; Fisher, A.R.; et al. Altered profile of human gut microbiome is associated with cirrhosis and its complications. J. Hepatol. 2014, 60, 940–947. [Google Scholar] [CrossRef]
- Liu, Y.; Ajami, N.J.; El-Serag, H.B.; Hair, C.; Graham, D.Y.; White, D.L.; Chen, L.; Wang, Z.; Plew, S.; Kramer, J.; et al. Dietary quality and the colonic mucosa-associated gut microbiome in humans. Am. J. Clin. Nutr. 2019, 110, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.W.; Fouts, D.E.; Brandl, J.; Stärkel, P.; Torralba, M.; Schott, E.; Tsukamoto, H.; Nelson, K.E.; Brenner, D.A.; Schnabl, B. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology 2011, 53, 96–105. [Google Scholar] [CrossRef]
- Palmisano, L.; Giuliano, M.; Bucciardini, R.; Andreotti, M.; Fragola, V.; Pirillo, M.F.; Weimer, L.E.; Mancini, M.G.; Vella, S. Modifications of residual viraemia in human immunodeficiency virus-1-infected subjects undergoing repeated highly active antiretroviral therapy interruptions. J. Med. Microbiol. 2009, 58, 121–124. [Google Scholar] [CrossRef] [PubMed]
- DallaPiazza, M.; Amorosa, V.K.; Localio, R.; Kostman, J.R.; Lo Re, V. Prevalence and risk factors for significant liver fibrosis among HIV-monoinfected patients. BMC Infect. Dis. 2010, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Human Metabolome Database: Showing Metabocard for Caffeine (HMDB0001847). Available online: https://hmdb.ca/metabolites/HMDB0001847 (accessed on 18 July 2021).
- Tůma, P.; Samcová, E.; Balínová, P. Determination of 3-methylhistidine and 1-methylhistidine in untreated urine samples by capillary electrophoresis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 821, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Arvind, A.; Osganian, S.A.; Cohen, D.E.; Corey, K.E. Lipid and Lipoprotein Metabolism in Liver Disease. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]



| Variable 1 | Total n = 50 | FIB-4 < 1.45 n = 36 | FIB-4 ≥ 1.45 2 n = 14 | p |
|---|---|---|---|---|
| Age, years | 55 ± 6.8 | 54.5 ± 7.5 | 56.2 ± 4.6 | 0.437 |
| Sex, male | 29 (58.0) | 19 (52.8) | 10 (71.4) | 0.341 |
| Race/ethnicity | 0.538 | |||
| White non-Hispanic | 3 (6.0) | 3 (8.3) | 0 | |
| Black non-Hispanic | 34 (68.0) | 22 (61.1) | 12 (85.7) | |
| White Hispanic | 12 (24.0) | 10 (27.8) | 2 (14.3) | |
| Other | 1 (2.0) | 1 (2.8) | 0 | |
| Education level | ||||
| <High school | 20 (40.0) | 17 (47.2) | 3 (21.4) | 0.227 |
| High school diploma/GED | 18 (36.0) | 12 (33.3) | 6 (42.9) | |
| Some college + | 12 (24.0) | 7 (19.4) | 5 (35.7) | |
| Annual income | 0.525 | |||
| <USD 12,500 | 34 (68.0) | 24 (66.7) | 10 (71.4) | |
| USD 12,500–USD 35,000 | 14 (28.0) | 11 (30.6) | 3 (21.4) | |
| ≥USD 35,000 | 2 (4.0) | 1 (2.8) | 1 (7.1) | |
| Tobacco use | 26 (52.0) | 17 (47.2) | 9 (64.3) | 0.196 |
| Non-smoker | 24 (48.0) | 19 (52.8) | 5 (35.7) | |
| Every day smoker | 13 (26) | 10 (27.8) | 3 (21.4) | |
| Some days smoker | 13 (26) | 7 (19.4) | 6 (42.9) | |
| Hazardous alcohol use 3 | 7 (14.0) | 5 (13.9) | 2 (14.3) | 1.000 |
| Cocaine use | 25 (50.0) | 17 (47.2) | 8 (57.1) | 0.754 |
| Obesity 4 | 20 (40.0) | 16 (44.4) | 4 (28.6) | 0.445 |
| Food insecurity 5 | 6 (12.0) | 3 (8.3) | 3 (21.4) | 0.331 |
| On ART | 49 (98.0) | 36 (100.0) | 13 (92.9) | 0.280 |
| ART regimen | ||||
| Multi-class combination products | 46 (92.0) | 35 (97.2) | 11 (78.6) | 0.061 |
| NRTI | 0 | 0 | 0 | - |
| NNRTI | 1 (2.0) | 0 | 1 (7.1) | 0.280 |
| Protease inhibitors | 7 (14.0) | 4 (11.1) | 3 (21.4) | 0.384 |
| Fusion inhibitors | 0 | 0 | 0 | - |
| Entry inhibitors | 0 | 0 | 0 | - |
| HIV integrase strand transfer inhibitors | 5 (10.0) | 3 (8.3) | 2 (14.3) | 0.611 |
| Pharmacokinetic enhancers | 0 | 0 | 0 | - |
| Post-attachment inhibitor | 0 | 0 | 0 | - |
| ART adherence 6 | 0.53 ± 1.3 | 0.41 ± 1.1 | 0.85 ± 1.7 | 0.316 |
| Virally suppressed 7 | 34 (68.0) | 27 (75.0) | 7 (50.0) | 0.105 |
| CD4 lymphocyte count, cells/µL | 634.6 ± 337.1 | 694.9 ± 367.0 | 479.4 ± 172.3 | 0.007 * |
| Years living with HIV | 17.6 ± 8.2 | 17.5 ± 7.8 | 17.8 ± 9.4 | 0.922 |
| Variable 1 | Max Possible Score | Total n = 50 | FIB-4 < 1.45 n = 36 | FIB-4 ≥ 1.45 2 n = 14 | p |
|---|---|---|---|---|---|
| Total caloric intake (kcal) | – | 2087 ± 757 | 2200 ± 804 | 1795 ± 540 | 0.089 |
| Adequacy components: | |||||
| Total fruit HEI score 3 | 5 | 2.16 ± 1.80 | 2.13 ± 1.86 | 2.23 ± 1.70 | 0.872 |
| Total fruit intake (cups) | – | 1.01 ± 1.35 | 1.13 ± 1.55 | 0.69 ± 0.56 | 0.142 |
| Whole fruit HEI score 4 | 5 | 1.48 ± 1.94 | 1.50 ± 1.89 | 1.45 ± 2.14 | 0.930 |
| Whole fruit intake (cups) | – | 0.51 ± 0.98 | 0.60 ± 1.10 | 0.28 ± 0.49 | 0.167 |
| Total vegetable HEI score 5 | 5 | 2.53 ± 1.43 | 2.64 ± 1.50 | 2.23 ± 1.22 | 0.370 |
| Total vegetable intake (cups) | – | 1.22 ± 0.91 | 1.31 ± 0.91 | 1.00 ± 0.91 | 0.292 |
| Greens & beans HEI score 5 | 5 | 4.05 ± 1.56 | 4.23 ± 1.46 | 3.60 ± 1.79 | 0.205 |
| Greens & beans (cups) | – | 0.66 ± 0.60 | 0.74 ± 0.62 | 0.47 ± 0.52 | 0.157 |
| Whole grains HEI score | 10 | 2.80 ± 3.25 | 2.61 ± 3.30 | 3.26 ± 3.18 | 0.533 |
| Whole grains intake (oz) | – | 1.00 ± 1.47 | 0.95 ± 1.45 | 1.12 ± 1.56 | 0.720 |
| Dairy HEI score 6 | 10 | 2.32 ± 1.80 | 2.65 ± 1.77 | 1.47 ± 1.65 | 0.036 * |
| Dairy intake (cups) | – | 0.61 ± 0.47 | 0.73 ± 0.48 | 0.33 ± 0.29 | 0.006 * |
| Total protein foods HEI score 5 | 5 | 5.00 ± 0.00 | 5.00 ± 0.00 | 5.00 ± 0.00 | 1.000 |
| Protein intake (oz) | – | 10.90 ± 4.45 | 10.74 ± 4.52 | 11.34 ± 4.41 | 0.672 |
| Seafood and plant proteins HEI score 5,7 | 5 | 4.15 ± 1.44 | 4.31 ± 1.35 | 3.74 ± 1.63 | 0.214 |
| Seafood/plant protein intake (oz) | – | 3.03 ± 2.45 | 3.35 ± 2.61 | 2.21 ± 1.82 | 0.141 |
| Fatty acids HEI score 8 | 10 | 0.27 ± 1.01 | 0.24 ± 1.02 | 0.37 ± 1.00 | 0.688 |
| Polyunsaturated fatty acid intake (g) | – | 4.84 ± 5.41 | 5.17 ± 5.94 | 3.99 ± 3.78 | 0.493 |
| Monounsaturated fatty acid intake (g) | – | 6.87 ± 6.19 | 7.51 ± 6.85 | 5.21 ± 3.72 | 0.135 |
| Moderation components: | |||||
| Saturated fatty acid HEI score | 10 | 6.37 ± 3.04 | 6.43 ± 2.88 | 6.21 ± 3.53 | 0.822 |
| Saturated fatty acid intake (g) | – | 25.72 ± 13.33 | 27.00 ± 13.48 | 22.41 ± 12.82 | 0.278 |
| Refined grains HEI score | 10 | 4.22 ± 3.62 | 4.19 ± 3.63 | 4.28 ± 3.72 | 0.939 |
| Refined grains intake (oz) | – | 7.37 ± 4.40 | 7.66 ± 4.52 | 6.63 ± 4.16 | 0.462 |
| Sodium HEI score | 10 | 3.93 ± 3.46 | 3.81 ± 3.45 | 4.24 ± 3.61 | 0.694 |
| Sodium intake (g) | – | 3.48 ± 1.16 | 3.63 ± 1.09 | 3.10 ± 1.30 | 0.151 |
| Added sugar HEI score | 10 | 6.39 ± 2.90 | 6.34 ± 3.05 | 6.52 ± 2.59 | 0.845 |
| Added sugar intake (% energy) | – | 0.13 ± 0.06 | 0.14 ± 0.07 | 0.13 ± 0.06 | 0.775 |
| HEI total score 9 | 100 | 45.67 ± 11.54 | 46.08 ± 11.52 | 44.60 ± 11.96 | 0.689 |
| Biochemical | Super Pathway | Sub Pathway | Metabolite | PLS-DA Effect Size | Spearman r | Spearman p-Value |
|---|---|---|---|---|---|---|
| Pregnen-diol disulfate * | Lipid | Pregnenolone steroids | 32,562 | 0.249 | 0.333 | 0.018 |
| 1-stearoyl-GPS (18:0) * | Lipid | Lysophospholipid | 45,966 | −0.253 | −0.283 | 0.047 |
| 1,3-dimethylurate | Xenobiotics | Xanthine metabolism | 32,391 | −0.090 | −0.292 | 0.040 |
| 2-deoxyuridine | Nucleotide | Pyrimidine metabolism, uracil containing | 52,602 | 0.262 | 0.344 | 0.015 |
| Serotonin | Amino acid | Tryptophan metabolism | 2342 | −0.059 | −0.316 | 0.025 |
| Inosine 5-monophosphate (IMP) | Nucleotide | Purine metabolism, (hypo)xanthine/inosine containing | 2133 | −0.174 | −0.396 | 0.004 |
| 1-stearoyl-2-oleoyl-GPS (18:0/18:1) | Lipid | Phosphatidylserine (PS) | 19265 | −0.192 | −0.391 | 0.005 |
| Cysteine-glutathione disulfide | Amino acid | Glutathione metabolism | 35,159 | −0.074 | −0.288 | 0.042 |
| 2-hydroxy-4-(methylthio)butanoic acid | Amino acid | Methionine, cysteine, SAM, and taurine metabolism | 63,739 | 0.109 | 0.299 | 0.035 |
| AMP | Nucleotide | Purine metabolism, adenine containing | 32,342 | −0.204 | −0.503 | 0.0002 |
| 9,10-diHOME | Lipid | Fatty acid, dihydroxy | 38,399 | 0.213 | 0.454 | 0.001 |
| 6-oxopiperidine-2-carboxylate | Amino acid | Lysine metabolism | 43,231 | 0.227 | 0.280 | 0.049 |
| Octadecanedioylcarnitine (C18-DC) * | Lipid | Fatty acid metabolism (Acyl carnitine, dicarboxylate) | 61,867 | 0.134 | 0.375 | 0.007 |
| S-methylcysteine sulfoxide | Amino acid | Methionine, cysteine, SAM, and taurine metabolism | 43,378 | −0.004 | −0.317 | 0.025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, H.R.; Sales Martinez, S.; Stebliankin, V.; Tamargo, J.A.; Campa, A.; Narasimhan, G.; Hernandez, J.; Rodriguez, J.A.B.; Teeman, C.; Johnson, A.; et al. Diet Quality and Liver Health in People Living with HIV in the MASH Cohort: A Multi-Omic Analysis of the Fecal Microbiome and Metabolome. Metabolites 2023, 13, 271. https://doi.org/10.3390/metabo13020271
Martin HR, Sales Martinez S, Stebliankin V, Tamargo JA, Campa A, Narasimhan G, Hernandez J, Rodriguez JAB, Teeman C, Johnson A, et al. Diet Quality and Liver Health in People Living with HIV in the MASH Cohort: A Multi-Omic Analysis of the Fecal Microbiome and Metabolome. Metabolites. 2023; 13(2):271. https://doi.org/10.3390/metabo13020271
Chicago/Turabian StyleMartin, Haley R., Sabrina Sales Martinez, Vitalii Stebliankin, Javier A. Tamargo, Adriana Campa, Giri Narasimhan, Jacqueline Hernandez, Jose A. Bastida Rodriguez, Colby Teeman, Angelique Johnson, and et al. 2023. "Diet Quality and Liver Health in People Living with HIV in the MASH Cohort: A Multi-Omic Analysis of the Fecal Microbiome and Metabolome" Metabolites 13, no. 2: 271. https://doi.org/10.3390/metabo13020271