
Zero Echo Time 17O-MRI Reveals Decreased Cerebral Metabolic Rate of Oxygen Consumption in a Murine Model of Amyloidosis

 , , , ,
, , , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
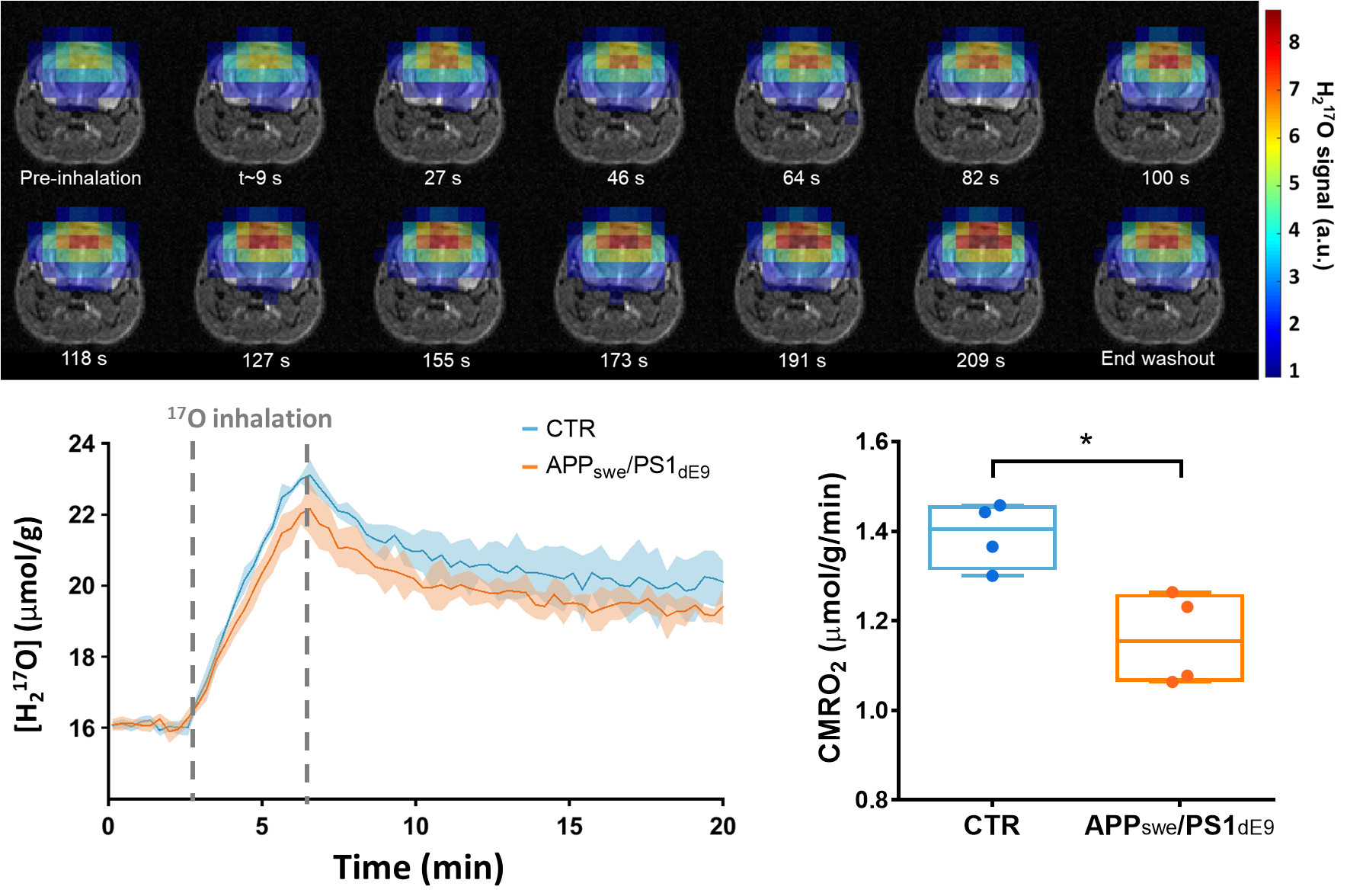
2.1. Setting Up CMRO2 Measurements in the Mouse Brain
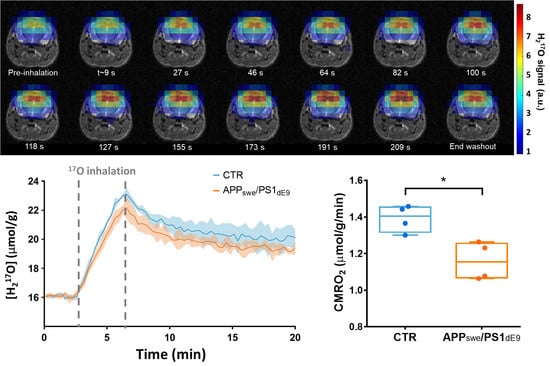
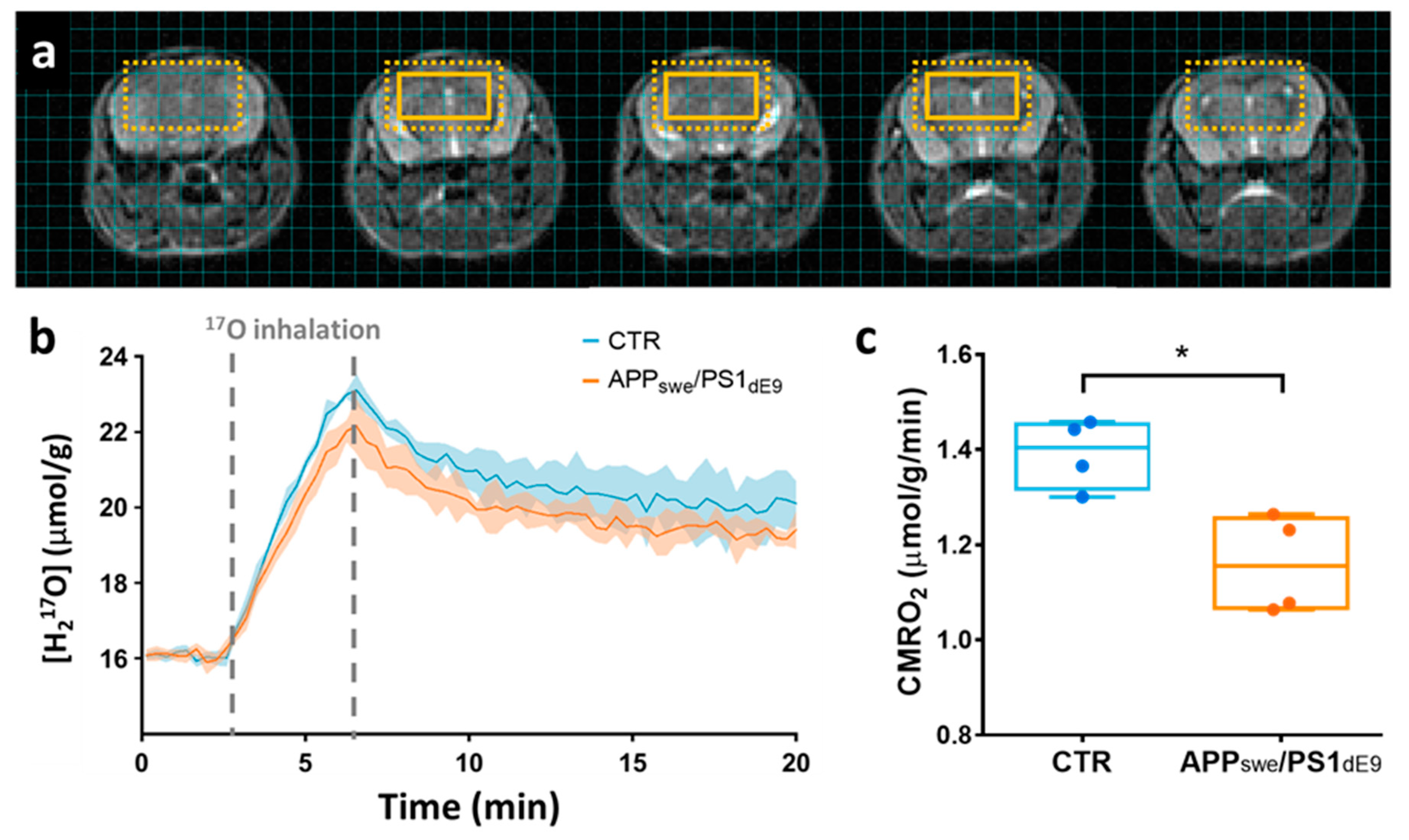
2.2. CMRO2 Is Lower in APPswe/PS1dE9 Than in Wild-Type Mice
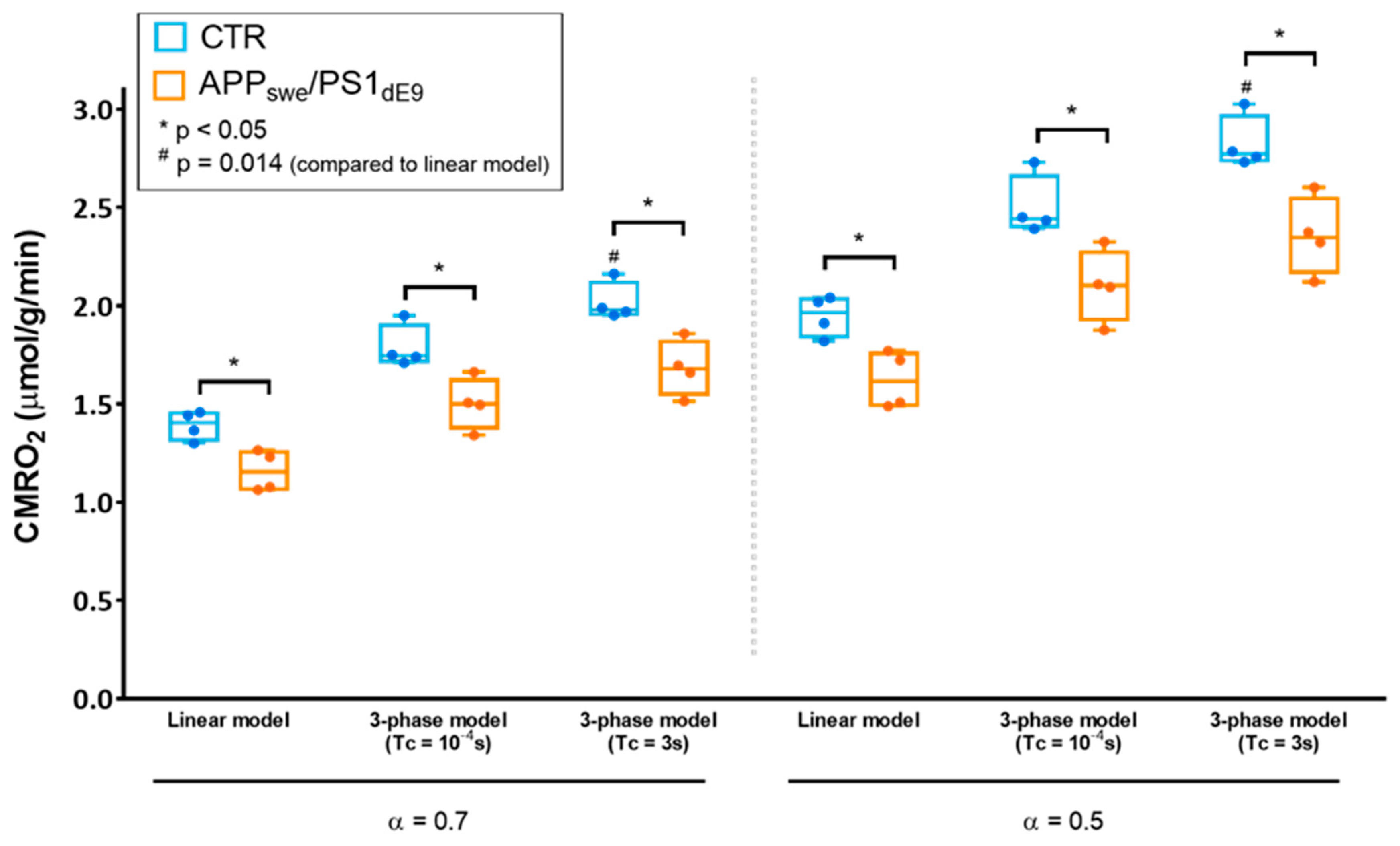
2.3. Effect of the Choice of the Model on CMRO2 Quantification
3. Discussion
4. Materials and Methods
4.1. Animals and Preparation
4.2. MR Methods
4.2.1. SNR Comparison Experiments
4.2.2. 17O2 Inhalation Experiments
4.3. CMRO2 Calculation
4.3.1. Three-Phase Model
4.3.2. Simplified Model
4.4. CBF Estimation
4.5. Immunohistochemistry
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Readnower, R.D.; Sauerbeck, A.D.; Sullivan, P.G. Mitochondria, Amyloid beta, and Alzheimer’s Disease. Int. J. Alzheimers Dis. 2011, 2011, 104545. [Google Scholar] [CrossRef] [Green Version]
- Fukuyama, H.; Ogawa, M.; Yamauchi, H.; Yamaguchi, S.; Kimura, J.; Yonekura, Y.; Konishi, J. Altered cerebral energy metabolism in Alzheimer’s disease: A PET study. J. Nucl. Med. 1994, 35, 1–6. [Google Scholar]
- Horitsugi, G.; Watabe, T.; Kanai, Y.; Ikeda, H.; Kato, H.; Naka, S.; Ishibashi, M.; Matsunaga, K.; Isohashi, K.; Shimosegawa, E.; et al. Oxygen-15 labeled CO2, O2, and CO PET in small animals: Evaluation using a 3D-mode microPET scanner and impact of reconstruction algorithms. EJNMMI Res. 2017, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Watabe, T.; Shimosegawa, E.; Watabe, H.; Kanai, Y.; Hanaoka, K.; Ueguchi, T.; Isohashi, K.; Kato, H.; Tatsumi, M.; Hatazawa, J. Quantitative evaluation of cerebral blood flow and oxygen metabolism in normal anesthetized rats: 15O-labeled gas inhalation PET with MRI Fusion. J. Nucl. Med. 2013, 54, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Camp, N.; Bramoullé, Y.; Hantraye, P. Quantification of Brain Function and Neurotransmission System In Vivo by Positron Emission Tomography: A Review of Technical Aspects and Practical Considerations in Preclinical Research. In Animal Models of Movement Disorders, 1st ed.; Lane, E.L., Dunett, S.B., Eds.; Neuromethods; Humana Press: New York, NY, USA, 2012; Volume I, pp. 151–188. [Google Scholar]
- Temma, T.; Yamazaki, M.; Miyanohara, J.; Shirakawa, H.; Kondo, N.; Koshino, K.; Kaneko, S.; Iida, H. Sequential PET estimation of cerebral oxygen metabolism with spontaneous respiration of (15)O-gas in mice with bilateral common carotid artery stenosis. J. Cereb. Blood Flow Metab. 2017, 37, 3334–3343. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, C.J.; Desjardins-Crepeau, L.; Madjar, C.; Bherer, L.; Hoge, R.D. Absolute quantification of resting oxygen metabolism and metabolic reactivity during functional activation using QUO2 MRI. Neuroimage 2012, 63, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, I.C.; Thulborn, K.R. Feasibility of mapping the tissue mass corrected bioscale of cerebral metabolic rate of oxygen consumption using 17-oxygen and 23-sodium MR imaging in a human brain at 9.4 T. Neuroimage 2010, 51, 723–733. [Google Scholar] [CrossRef]
- Hoffmann, S.H.; Radbruch, A.; Bock, M.; Semmler, W.; Nagel, A.M. Direct (17)O MRI with partial volume correction: First experiences in a glioblastoma patient. MAGMA 2014, 27, 579–587. [Google Scholar] [CrossRef]
- Kurzhunov, D.; Borowiak, R.; Reisert, M.; Ozen, A.C.; Bock, M. Direct estimation of (17) O MR images (DIESIS) for quantification of oxygen metabolism in the human brain with partial volume correction. Magn. Reson. Med. 2018, 80, 2717–2725. [Google Scholar] [CrossRef]
- Cui, W.; Zhu, X.H.; Vollmers, M.L.; Colonna, E.T.; Adriany, G.; Tramm, B.; Dubinsky, J.M.; Oz, G. Non-invasive measurement of cerebral oxygen metabolism in the mouse brain by ultra-high field (17)O MR spectroscopy. J. Cereb. Blood Flow. Metab. 2013, 33, 1846–1849. [Google Scholar] [CrossRef] [Green Version]
- Lou, S.; Lepak, V.C.; Eberly, L.E.; Roth, B.; Cui, W.; Zhu, X.H.; Oz, G.; Dubinsky, J.M. Oxygen consumption deficit in Huntington disease mouse brain under metabolic stress. Hum. Mol. Genet. 2016, 25, 2813–2826. [Google Scholar] [CrossRef] [Green Version]
- Wiesner, H.M.; Balla, D.Z.; Scheffler, K.; Ugurbil, K.; Zhu, X.H.; Chen, W.; Uludag, K.; Pohmann, R. Quantitative and simultaneous measurement of oxygen consumption rates in rat brain and skeletal muscle using (17) O MRS imaging at 16.4T. Magn. Reson. Med. 2021, 85, 2232–2246. [Google Scholar] [CrossRef]
- Zhu, X.H.; Zhang, Y.; Tian, R.X.; Lei, H.; Zhang, N.; Zhang, X.; Merkle, H.; Ugurbil, K.; Chen, W. Development of (17)O NMR approach for fast imaging of cerebral metabolic rate of oxygen in rat brain at high field. Proc. Natl. Acad. Sci. USA 2002, 99, 13194–13199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafner, S. Fast imaging in liquids and solids with the Back-projection Low Angle ShoT (BLAST) technique. Magn. Reson. Imaging 1994, 12, 1047–1051. [Google Scholar] [CrossRef]
- Zhu, X.H.; Chen, J.M.; Tu, T.W.; Chen, W.; Song, S.K. Simultaneous and noninvasive imaging of cerebral oxygen metabolic rate, blood flow and oxygen extraction fraction in stroke mice. Neuroimage 2013, 64, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neveu, M.A.; Joudiou, N.; de Preter, G.; Dehoux, J.P.; Jordan, B.F.; Gallez, B. (17) O MRS assesses the effect of mild hypothermia on oxygen consumption rate in tumors. NMR Biomed. 2017, 30, e3726. [Google Scholar] [CrossRef]
- Zhang, N.; Zhu, X.H.; Lei, H.; Ugurbil, K.; Chen, W. Simplified methods for calculating cerebral metabolic rate of oxygen based on 17O magnetic resonance spectroscopic imaging measurement during a short 17O2 inhalation. J. Cereb. Blood Flow Metab. 2004, 24, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Rovainen, C.M.; Woolsey, T.A.; Blocher, N.C.; Wang, D.B.; Robinson, O.F. Blood flow in single surface arterioles and venules on the mouse somatosensory cortex measured with videomicroscopy, fluorescent dextrans, nonoccluding fluorescent beads, and computer-assisted image analysis. J. Cereb. Blood Flow Metab. 1993, 13, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Nakao, S.; Mori, K.; Ishimori, K.; Morishima, I.; Miyazawa, T.; Fritz-Zieroth, B. Cerebral oxygen utilization analyzed by the use of oxygen-17 and its nuclear magnetic resonance. Biochem. Biophys. Res. Commun. 1990, 169, 153–158. [Google Scholar] [CrossRef]
- Zhu, X.H.; Chen, W. In vivo(17)O MRS imaging—Quantitative assessment of regional oxygen consumption and perfusion rates in living brain. Anal. Biochem. 2017, 529, 171–178. [Google Scholar] [CrossRef]
- Calsolaro, V.; Edison, P. Alterations in Glucose Metabolism in Alzheimer’s Disease. Recent Pat. Endocr. Metab. Immune Drug Discov. 2016, 10, 31–39. [Google Scholar] [CrossRef]
- Landau, S.M.; Mintun, M.A.; Joshi, A.D.; Koeppe, R.A.; Petersen, R.C.; Aisen, P.S.; Weiner, M.W.; Jagust, W.J.; Alzheimer’s Disease Neuroimaging Initiative. Amyloid deposition, hypometabolism, and longitudinal cognitive decline. Ann. Neurol. 2012, 72, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, L. Brain glucose metabolism in the early and specific diagnosis of Alzheimer’s disease. FDG-PET studies in MCI and AD. Eur. J. Nucl. Med. Mol. Imaging 2005, 32, 486–510. [Google Scholar] [CrossRef]
- Chen, J.X.; Yan, S.D. Amyloid-beta-induced mitochondrial dysfunction. J. Alzheimers Dis. 2007, 12, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Rudin, M.; Klohs, J. Cortical hypoperfusion and reduced cerebral metabolic rate of oxygen in the arcAbeta mouse model of Alzheimer’s disease. Photoacoustics 2018, 10, 38–47. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Robbins, E.M.; Zhang-Nunes, S.X.; Purcell, S.M.; Betensky, R.A.; Raju, S.; Prada, C.; Greenberg, S.M.; Bacskai, B.J.; Frosch, M.P. Characterization of amyloid deposition in the APPswe/PS1dE9 mouse model of Alzheimer disease. Neurobiol. Dis. 2006, 24, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.J.; Mei, Y.; Shi, X.Q.; Zhang, Y.F.; Wang, X.Y.; Guan, L.; Wang, Q.; Pan, H.F. Albiflorin ameliorates memory deficits in APP/PS1 transgenic mice via ameliorating mitochondrial dysfunction. Brain Res. 2019, 1719, 113–123. [Google Scholar] [CrossRef]
- Chen, L.; Xu, S.; Wu, T.; Shao, Y.; Luo, L.; Zhou, L.; Ou, S.; Tang, H.; Huang, W.; Guo, K.; et al. Studies on APP metabolism related to age-associated mitochondrial dysfunction in APP/PS1 transgenic mice. Aging 2019, 11, 10242–10251. [Google Scholar] [CrossRef]
- Poisnel, G.; Herard, A.S.; El Tannir El Tayara, N.; Bourrin, E.; Volk, A.; Kober, F.; Delatour, B.; Delzescaux, T.; Debeir, T.; Rooney, T.; et al. Increased regional cerebral glucose uptake in an APP/PS1 model of Alzheimer’s disease. Neurobiol. Aging 2012, 33, 1995–2005. [Google Scholar] [CrossRef] [Green Version]
- Moeini, M.; Lu, X.; Avti, P.K.; Damseh, R.; Belanger, S.; Picard, F.; Boas, D.; Kakkar, A.; Lesage, F. Compromised microvascular oxygen delivery increases brain tissue vulnerability with age. Sci. Rep. 2018, 8, 8219. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Chen, L.; Hou, X.; van Zijl, P.C.M.; Xu, J.; Lu, H. Age-Related Alterations in Brain Perfusion, Venous Oxygenation, and Oxygen Metabolic Rate of Mice: A 17-Month Longitudinal MRI Study. Front. Neurol. 2020, 11, 559. [Google Scholar] [CrossRef]
- Aanerud, J.; Borghammer, P.; Chakravarty, M.M.; Vang, K.; Rodell, A.B.; Jonsdottir, K.Y.; Moller, A.; Ashkanian, M.; Vafaee, M.S.; Iversen, P.; et al. Brain energy metabolism and blood flow differences in healthy aging. J. Cereb. Blood Flow Metab. 2012, 32, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Eustache, F.; Rioux, P.; Desgranges, B.; Marchal, G.; Petit-Taboue, M.C.; Dary, M.; Lechevalier, B.; Baron, J.C. Healthy aging, memory subsystems and regional cerebral oxygen consumption. Neuropsychologia 1995, 33, 867–887. [Google Scholar] [CrossRef]
- Pantano, P.; Baron, J.C.; Lebrun-Grandie, P.; Duquesnoy, N.; Bousser, M.G.; Comar, D. Regional cerebral blood flow and oxygen consumption in human aging. Stroke 1984, 15, 635–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Kanno, I.; Uemura, K.; Shishido, F.; Inugami, A.; Ogawa, T.; Murakami, M.; Suzuki, K. Reduction in regional cerebral metabolic rate of oxygen during human aging. Stroke 1986, 17, 1220–1228. [Google Scholar] [CrossRef] [Green Version]
- Nakai, M.; Yamamoto, J.; Matsui, Y. Acute systemic and regional hemodynamic effects of alpha 1-adrenoceptor blockade in conscious spontaneously hypertensive rats. Clin. Exp. Hypertens A 1986, 8, 981–996. [Google Scholar] [CrossRef]
- Petrinovic, M.M.; Hankov, G.; Schroeter, A.; Bruns, A.; Rudin, M.; von Kienlin, M.; Kunnecke, B.; Mueggler, T. A novel anesthesia regime enables neurofunctional studies and imaging genetics across mouse strains. Sci. Rep. 2016, 6, 24523. [Google Scholar] [CrossRef] [Green Version]
- Eger, E.I., 2nd. The pharmacology of isoflurane. Br. J. Anaesth. 1984, 56 (Suppl. 1), 71S–99S. [Google Scholar]
- Kint, L.T.; Seewoo, B.J.; Hyndman, T.H.; Clarke, M.W.; Edwards, S.H.; Rodger, J.; Feindel, K.W.; Musk, G.C. The Pharmacokinetics of Medetomidine Administered Subcutaneously during Isoflurane Anaesthesia in Sprague-Dawley Rats. Animals 2020, 10, 1050. [Google Scholar] [CrossRef]
- Hyder, F.; Herman, P.; Bailey, C.J.; Moller, A.; Globinsky, R.; Fulbright, R.K.; Rothman, D.L.; Gjedde, A. Uniform distributions of glucose oxidation and oxygen extraction in gray matter of normal human brain: No evidence of regional differences of aerobic glycolysis. J. Cereb. Blood Flow Metab. 2016, 36, 903–916. [Google Scholar] [CrossRef] [Green Version]
- Vlassenko, A.G.; Raichle, M.E. Brain aerobic glycolysis functions and Alzheimer’s disease. Clin. Transl. Imaging 2015, 3, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudeffant, C.; Vandesquille, M.; Herbert, K.; Garin, C.M.; Alves, S.; Blanchard, V.; Comoy, E.E.; Petit, F.; Dhenain, M. Contrast-enhanced MR microscopy of amyloid plaques in five mouse models of amyloidosis and in human Alzheimer’s disease brains. Sci. Rep. 2017, 7, 4955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streif, J.U.; Hiller, K.H.; Waller, C.; Nahrendorf, M.; Wiesmann, F.; Bauer, W.R.; Rommel, E.; Haase, A. In vivo assessment of absolute perfusion in the murine skeletal muscle with spin labeling MRI. Magnetic resonance imaging. J. Magn. Reson. Imaging 2003, 17, 147–152. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baligand, C.; Barret, O.; Tourais, A.; Pérot, J.-B.; Thenadey, D.; Petit, F.; Liot, G.; Gaillard, M.-C.; Flament, J.; Dhenain, M.; et al. Zero Echo Time 17O-MRI Reveals Decreased Cerebral Metabolic Rate of Oxygen Consumption in a Murine Model of Amyloidosis. Metabolites 2021, 11, 263. https://doi.org/10.3390/metabo11050263
Baligand C, Barret O, Tourais A, Pérot J-B, Thenadey D, Petit F, Liot G, Gaillard M-C, Flament J, Dhenain M, et al. Zero Echo Time 17O-MRI Reveals Decreased Cerebral Metabolic Rate of Oxygen Consumption in a Murine Model of Amyloidosis. Metabolites. 2021; 11(5):263. https://doi.org/10.3390/metabo11050263
Chicago/Turabian StyleBaligand, Celine, Olivier Barret, Amélie Tourais, Jean-Baptiste Pérot, Didier Thenadey, Fanny Petit, Géraldine Liot, Marie-Claude Gaillard, Julien Flament, Marc Dhenain, and et al. 2021. "Zero Echo Time 17O-MRI Reveals Decreased Cerebral Metabolic Rate of Oxygen Consumption in a Murine Model of Amyloidosis" Metabolites 11, no. 5: 263. https://doi.org/10.3390/metabo11050263