
mICA-Based fMRI Analysis of Specific CO2-Level-Dependent BOLD Signal Changes in the Human Brainstem
 , , , , ,
, , , , , 
Abstract
:1. Introduction
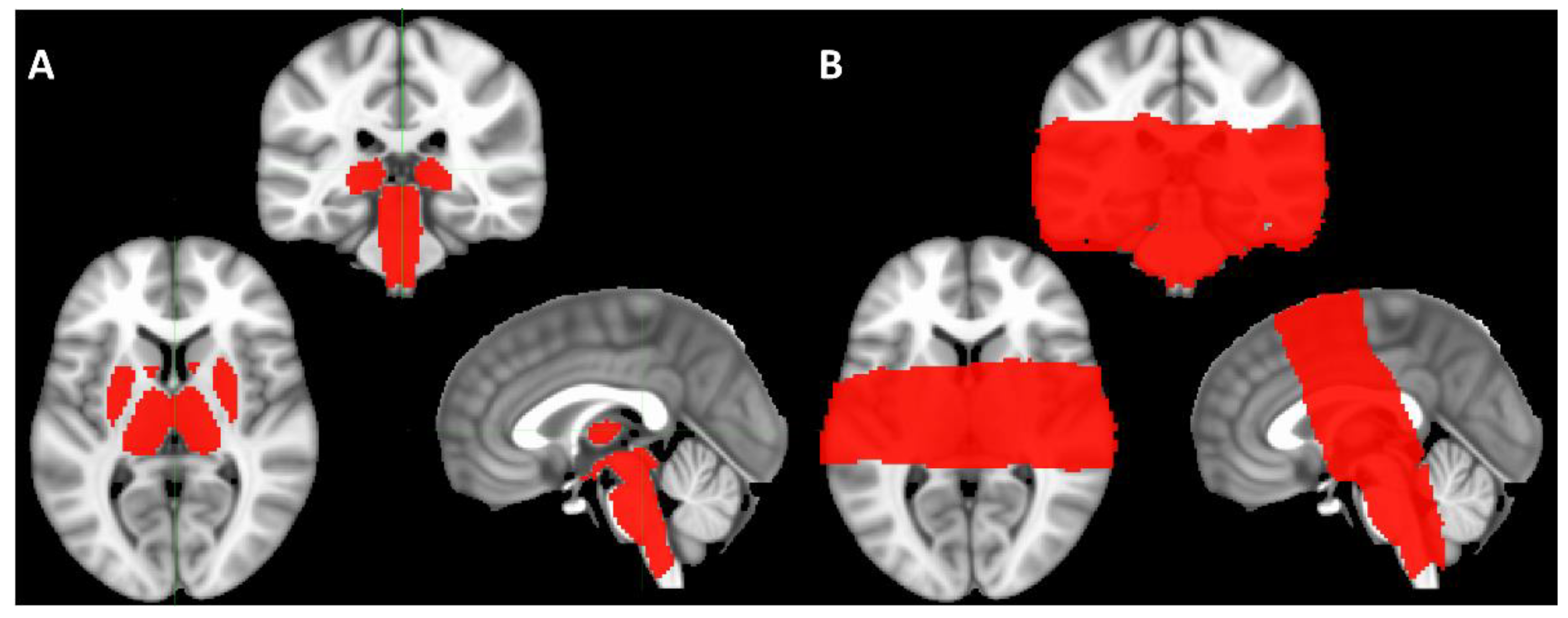
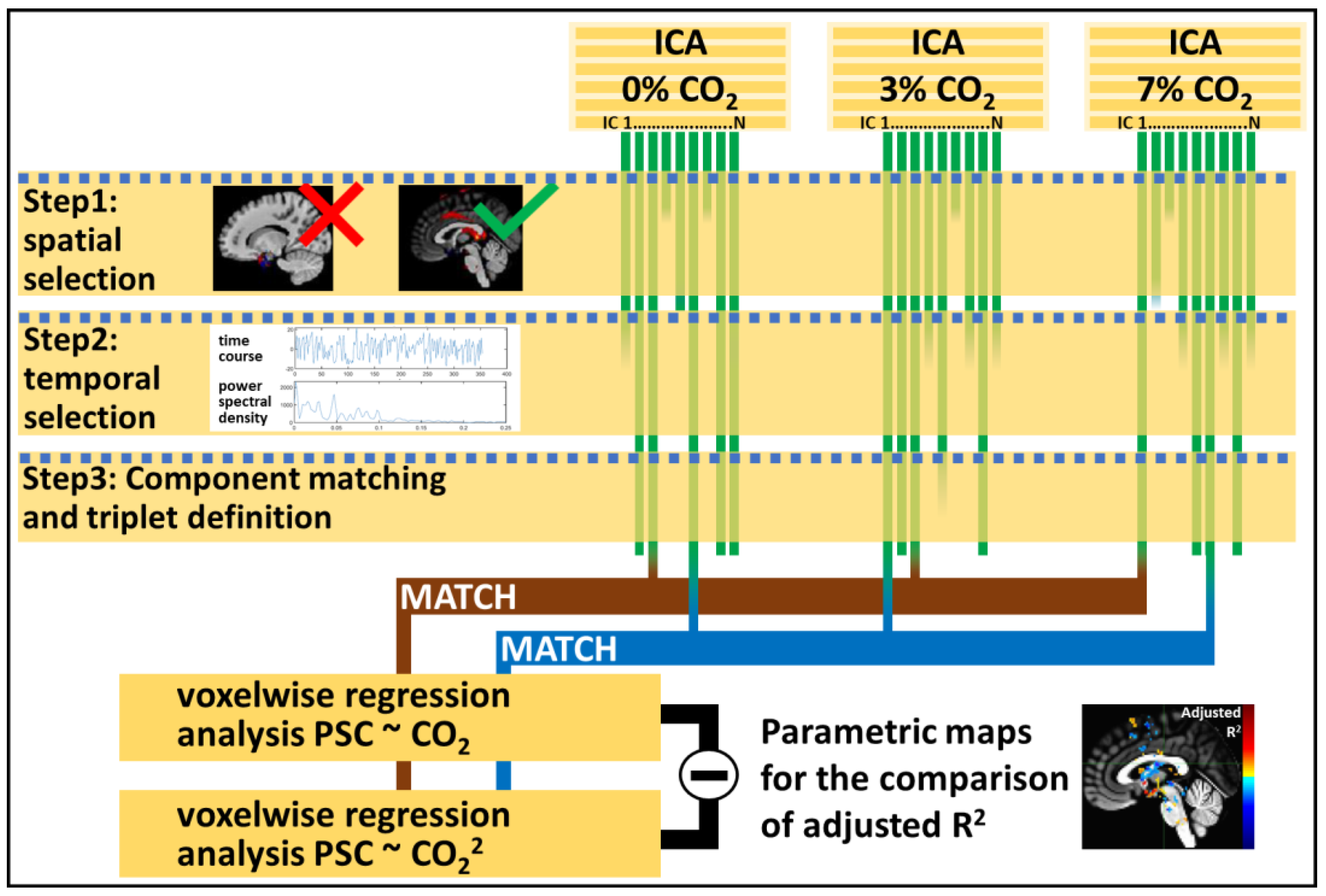
2. Materials and Methods
- (1)
- b2 > 0 as tested with a t-test with alpha = 5%
- (2)
- a1 > 0 as tested with a t-test with alpha = 5%
- (3)
- y (x = 3%) > y (x = 0%)
3. Results
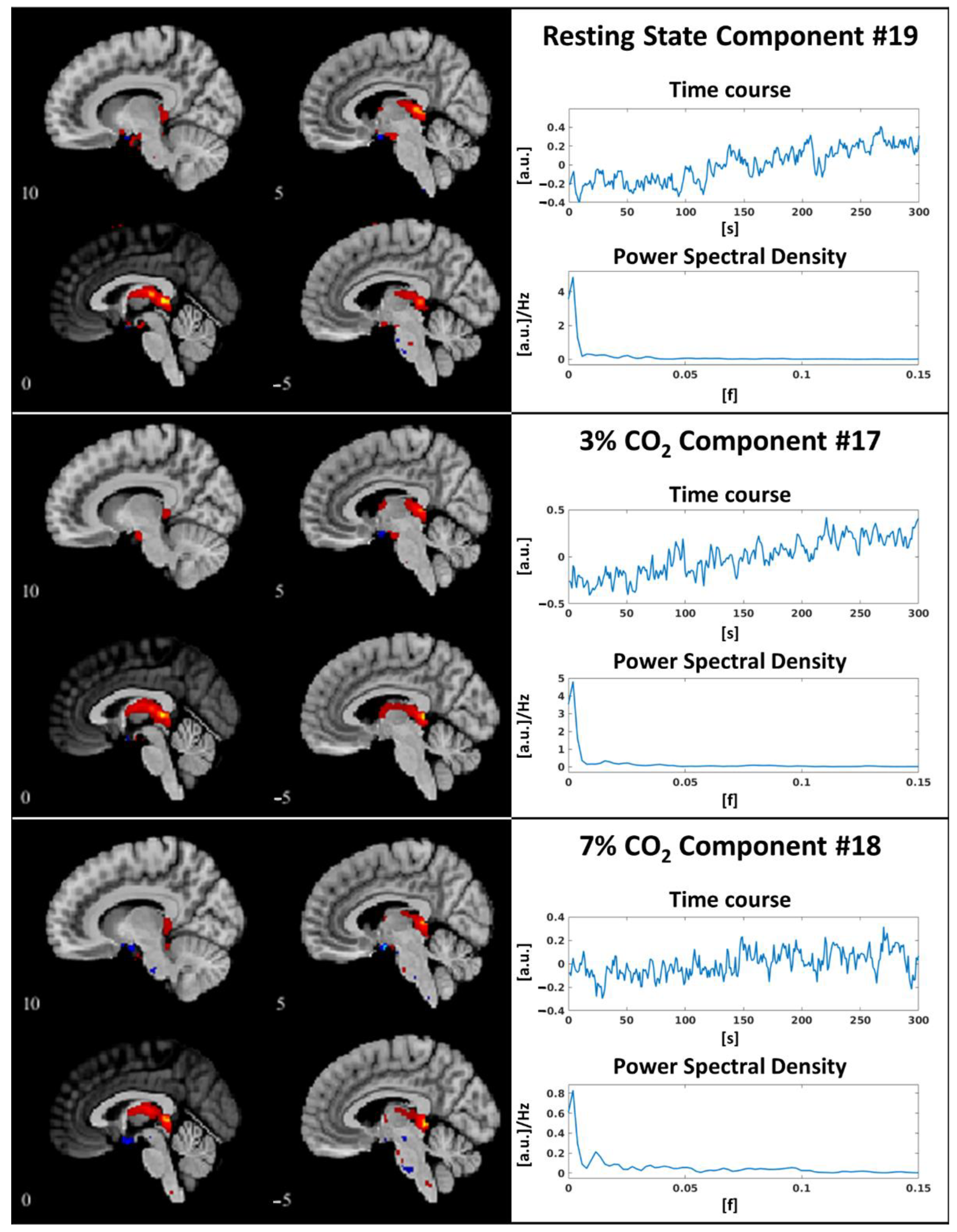
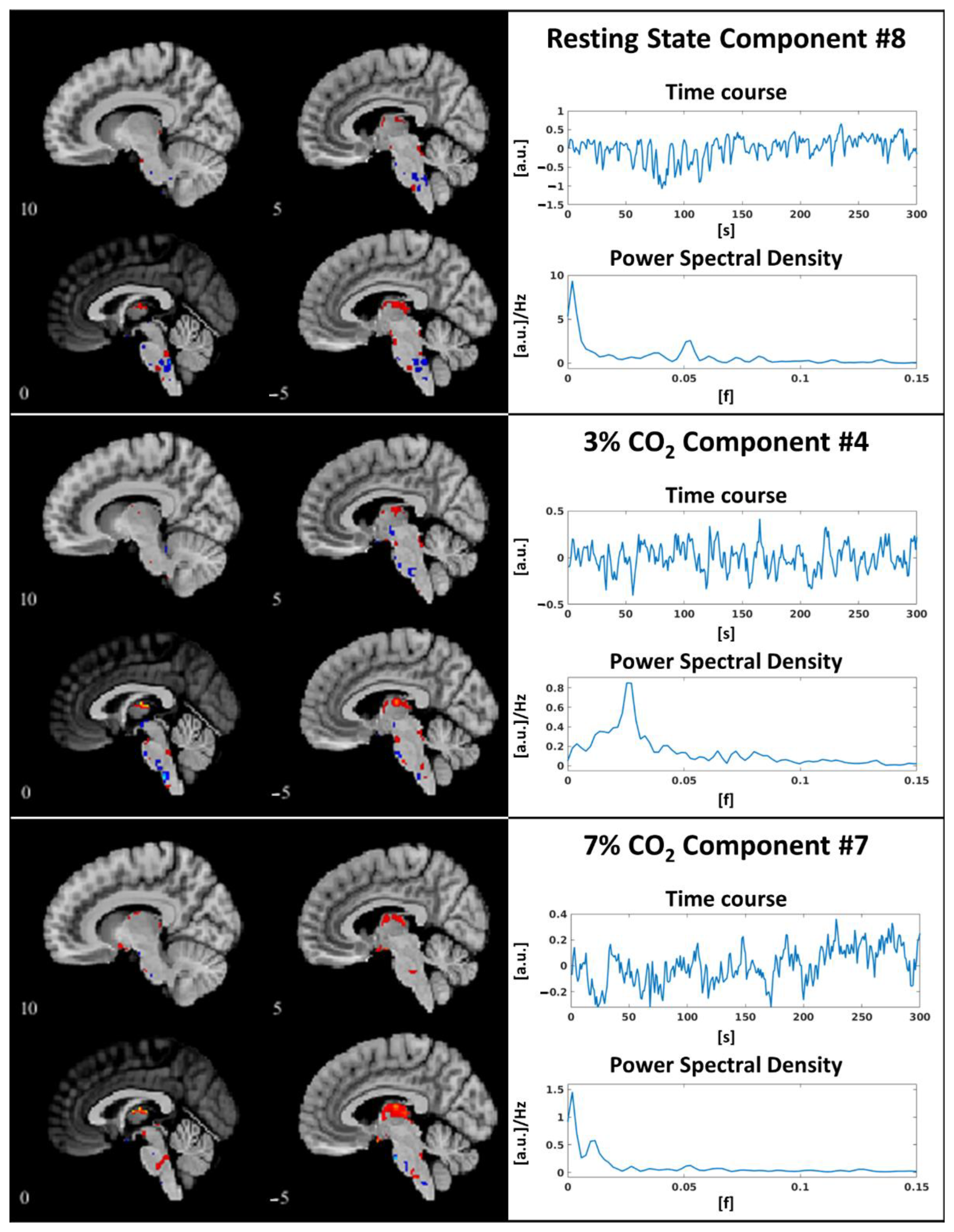
3.1. Independent Component Selection and Triad Identification
3.2. Quadratic Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Smith, J.C.; Abdala, A.P.L.; Borgmann, A.; Rybak, I.A.; Paton, J.F.R. Brainstem respiratory networks: Building blocks and microcircuits. Trends Neurosci. 2013, 36, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Giannoni, A.; Gentile, F.; Navari, A.; Borrelli, C.; Mirizzi, G.; Catapano, G.; Vergaro, G.; Grotti, F.; Betta, M.; Piepoli, M.F.; et al. Contribution of the Lung to the Genesis of Cheyne-Stokes Respiration in Heart Failure: Plant Gain Beyond Chemoreflex Gain and Circulation Time. J. Am. Heart Assoc. 2019, 8, e012419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannoni, A.; Gentile, F.; Buoncristiani, F.; Borrelli, C.; Sciarrone, P.; Spiesshoefer, J.; Bramanti, F.; Iudice, G.; Javaheri, S.; Emdin, M.; et al. Chemoreflex and Baroreflex Sensitivity Hold a Strong Prognostic Value in Chronic Heart Failure. JACC Heart Fail. 2022, 10, 662–676. [Google Scholar] [CrossRef] [PubMed]
- Giannoni, A.; Morelli, M.S.; Francis, D. Pathophysiology of Central Apneas in Heart Failure. In The Breathless Heart; Springer International Publishing: Cham, Switzerland, 2017; pp. 91–123. [Google Scholar]
- Devinsky, O.; Hesdorffer, D.C.; Thurman, D.J.; Lhatoo, S.; Richerson, G. Sudden unexpected death in epilepsy: Epidemiology, mechanisms, and prevention. Lancet Neurol. 2016, 15, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Kinney, H.C.; Richerson, G.B.; Dymecki, S.M.; Darnall, R.A.; Nattie, E.E. The Brainstem and Serotonin in the Sudden Infant Death Syndrome. Annu. Rev. Pathol. Mech. Dis. 2009, 4, 517–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianciardi, M.; Toschi, N.; Edlow, B.L.; Eichner, C.; Setsompop, K.; Polimeni, J.R.; Brown, E.N.; Kinney, H.C.; Rosen, B.R.; Wald, L.L. Toward an In Vivo Neuroimaging Template of Human Brainstem Nuclei of the Ascending Arousal, Autonomic, and Motor Systems. Brain Connect. 2015, 5, 597–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Cauzzo, S.; Garcia Gomar, M.G.; Stauder, M.; Vanello, N.; Passino, C.; Bianciardi, M.; García-Gomar, M.G.; Stauder, M.; Vanello, N.; et al. Functional connectome of arousal and motor brainstem nuclei in living humans by 7 Tesla resting-state fMRI. Neuroimage 2022, 249, 118865. [Google Scholar] [CrossRef]
- Brooks, J.C.W.; Faull, O.K.; Pattinson, K.T.S.; Jenkinson, M. Physiological Noise in Brainstem fMRI. Front. Hum. Neurosci. 2013, 7, 623. [Google Scholar] [CrossRef] [Green Version]
- Beissner, F. Functional MRI of the Brainstem: Common Problems and their Solutions. Clin. Neuroradiol. 2015, 25, 251–257. [Google Scholar] [CrossRef]
- Cauzzo, S.; Callara, A.L.; Morelli, M.S.; Hartwig, V.; Esposito, F.; Montanaro, D.; Passino, C.; Emdin, M.; Giannoni, A.; Vanello, N. Mapping dependencies of BOLD signal change to end-tidal CO2: Linear and nonlinear modeling, and effect of physiological noise correction. J. Neurosci. Methods 2021, 362, 109317. [Google Scholar] [CrossRef]
- Ciumas, C.; Rheims, S.; Ryvlin, P. fMRI studies evaluating central respiratory control in humans. Front. Neural Circuits 2022, 16, 982963. [Google Scholar] [CrossRef] [PubMed]
- Pattinson, K.T.S.; Mitsis, G.D.; Harvey, A.K.; Jbabdi, S.; Dirckx, S.; Mayhew, S.D.; Rogers, R.; Tracey, I.; Wise, R.G. Determination of the human brainstem respiratory control network and its cortical connections in vivo using functional and structural imaging. Neuroimage 2009, 44, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Beissner, F.; Schumann, A.; Brunn, F.; Eisenträger, D.; Bär, K.-J. Advances in functional magnetic resonance imaging of the human brainstem. Neuroimage 2014, 86, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napadow, V.; Dhond, R.; Park, K.; Kim, J.; Makris, N.; Kwong, K.K.; Harris, R.E.; Purdon, P.L.; Kettner, N.; Hui, K.K.S. Time-variant fMRI activity in the brainstem and higher structures in response to acupuncture. Neuroimage 2009, 47, 289–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, R.W. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, V.D.; Adali, T.; Pearlson, G.D.; Pekar, J.J. A method for making group inferences from functional MRI data using independent component analysis. Hum. Brain Mapp. 2001, 14, 140–151. [Google Scholar] [CrossRef]
- Vanello, N.; Ricciardi, E.; Landini, L. Analysis of Residual Dependencies of Independent Components Extracted from fMRI Data. Comput. Intell. Neurosci. 2016, 2016, 2961727. [Google Scholar] [CrossRef] [Green Version]
- Hyvärinen, A.; Oja, E. A Fast Fixed-Point Algorithm for Independent Component Analysis. Neural Comput. 1997, 9, 1483–1492. [Google Scholar] [CrossRef]
- Himberg, J.; Hyvärinen, A.; Esposito, F. Validating the independent components of neuroimaging time series via clustering and visualization. Neuroimage 2004, 22, 1214–1222. [Google Scholar] [CrossRef]
- Griffanti, L.; Douaud, G.; Bijsterbosch, J.; Evangelisti, S.; Alfaro-Almagro, F.; Glasser, M.F.; Duff, E.P.; Fitzgibbon, S.; Westphal, R.; Carone, D.; et al. Hand classification of fMRI ICA noise components. Neuroimage 2017, 154, 188–205. [Google Scholar] [CrossRef]
- Holland, P.W.; Welsch, R.E. Robust regression using iteratively reweighted least-squares. Commun. Stat. Theory Methods 1977, 6, 813–827. [Google Scholar] [CrossRef]
- Prokopiou, P.C.; Pattinson, K.T.S.; Wise, R.G.; Mitsis, G.D. Modeling of dynamic cerebrovascular reactivity to spontaneous and externally induced CO2 fluctuations in the human brain using BOLD-fMRI. Neuroimage 2019, 186, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Nattie, E.; Li, A. Central Chemoreceptors: Locations and Functions. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; Volume 2, pp. 221–254. ISBN 9788578110796. [Google Scholar]
- Saper, C.B. Convergence of autonomic and limbic connections in the insular cortex of the rat. J. Comp. Neurol. 1982, 210, 163–173. [Google Scholar] [CrossRef]
- Edlow, B.L.; Takahashi, E.; Wu, O.; Benner, T.; Dai, G.; Bu, L.; Grant, P.E.; Greer, D.M.; Greenberg, S.M.; Kinney, H.C.; et al. Neuroanatomic Connectivity of the Human Ascending Arousal System Critical to Consciousness and Its Disorders. J. Neuropathol. Exp. Neurol. 2012, 71, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Mark, G.P.; Shabani, S.; Dobbs, L.K.; Hansen, S.T. Cholinergic modulation of mesolimbic dopamine function and reward. Physiol. Behav. 2011, 104, 76–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Proch, K.L.; Teran, F.A.; Lechtenberg, R.J.; Kothari, H.; Richerson, G.B. Chemosensitivity of Phox2b-expressing retrotrapezoid neurons is mediated in part by input from 5-HT neurons. J. Physiol. 2019, 597, 2741–2766. [Google Scholar] [CrossRef]
- Iceman, K.E.; Richerson, G.B.; Harris, M.B. Medullary serotonin neurons are CO2 sensitive in situ. J. Neurophysiol. 2013, 110, 2536–2544. [Google Scholar] [CrossRef] [Green Version]
- Morelli, M.S.; Greco, A.; Valenza, G.; Giannoni, A.; Emdin, M.; Scilingo, E.P.; Vanello, N. Analysis of generic coupling between EEG activity and PETCO2 in free breathing and breath-hold tasks using Maximal Information Coefficient (MIC). Sci. Rep. 2018, 8, 4492. [Google Scholar] [CrossRef]
- Stickland, R.C.; Zvolanek, K.M.; Moia, S.; Ayyagari, A.; Caballero-Gaudes, C.; Bright, M.G. A practical modification to a resting state fMRI protocol for improved characterization of cerebrovascular function. Neuroimage 2021, 239, 118306. [Google Scholar] [CrossRef]
- Liu, P.; Xu, C.; Lin, Z.; Sur, S.; Li, Y.; Yasar, S.; Rosenberg, P.; Albert, M.; Lu, H. Cerebrovascular reactivity mapping using intermittent breath modulation. Neuroimage 2020, 215, 116787. [Google Scholar] [CrossRef]
- Colasanti, A.; Salamon, E.; Schruers, K.; van Diest, R.; van Duinen, M.; Griez, E.J. Carbon Dioxide-Induced Emotion and Respiratory Symptoms in Healthy Volunteers. Neuropsychopharmacology 2008, 33, 3103–3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Yang, Z. Impact of inter-individual variability on the estimation of default mode network in temporal concatenation group ICA. Neuroimage 2021, 237, 118114. [Google Scholar] [CrossRef] [PubMed]
- Kairov, U.; Cantini, L.; Greco, A.; Molkenov, A.; Czerwinska, U.; Barillot, E.; Zinovyev, A. Determining the optimal number of independent components for reproducible transcriptomic data analysis. BMC Genom. 2017, 18, 712. [Google Scholar] [CrossRef] [Green Version]
- Majeed, W.; Avison, M.J. Robust Data Driven Model Order Estimation for Independent Component Analysis of fMRI Data with Low Contrast to Noise. PLoS ONE 2014, 9, e94943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeown, M.J. Detection of Consistently Task-Related Activations in fMRI Data with Hybrid Independent Component Analysis. Neuroimage 2000, 11, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Miller, K.L.; Moeller, S.; Xu, J.; Auerbach, E.J.; Woolrich, M.W.; Beckmann, C.F.; Jenkinson, M.; Andersson, J.; Glasser, M.F.; et al. Temporally-independent functional modes of spontaneous brain activity. Proc. Natl. Acad. Sci. USA 2012, 109, 3131–3136. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
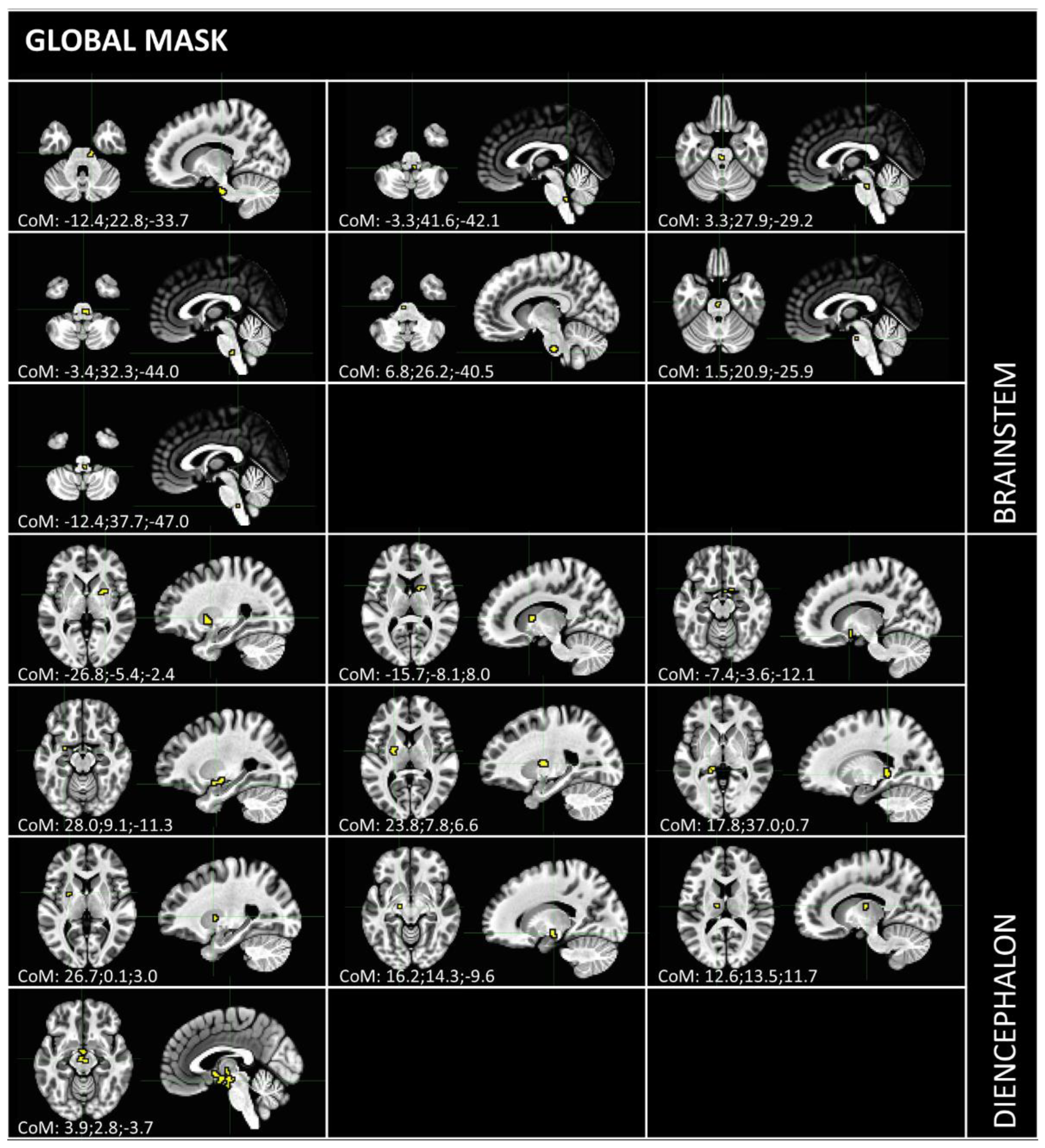{kind=link}
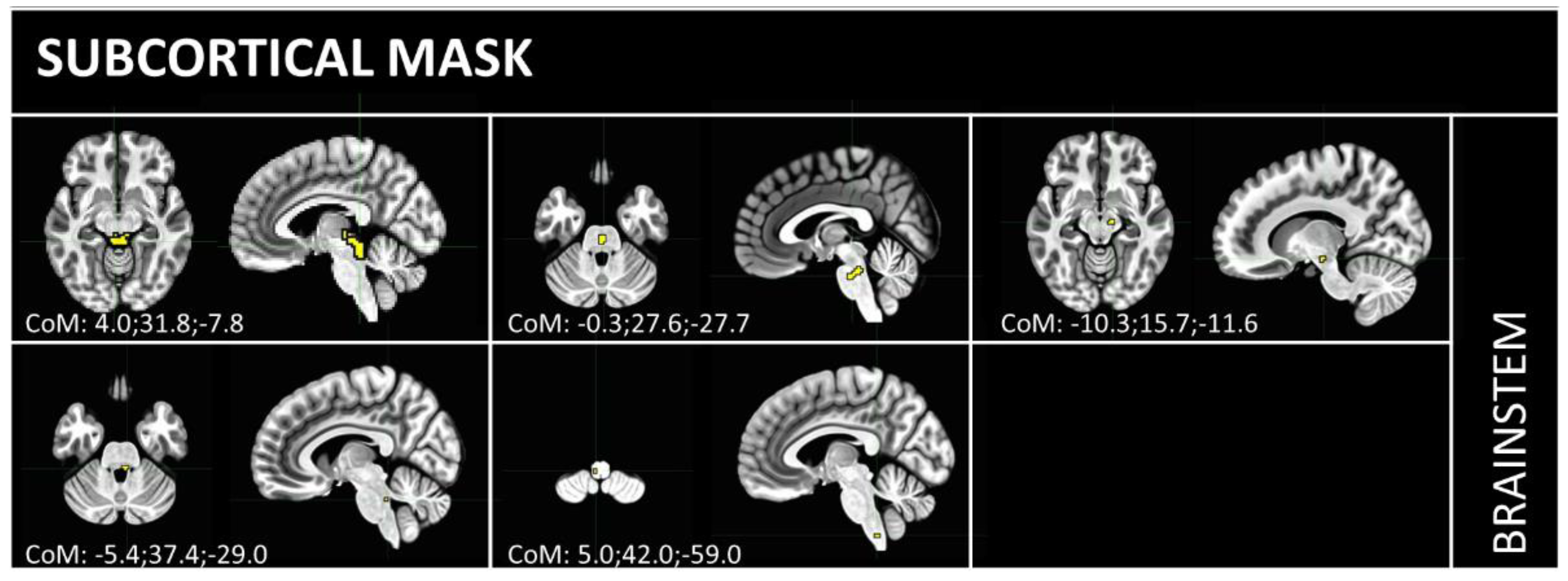{kind=link}
| Global Mask | Subcortical Mask | ||||
|---|---|---|---|---|---|
| 3–7% (ρ) | 3–0% (ρ) | 7–0% (ρ) | 3–7% (ρ) | 3–0% (ρ) | 7–0% (ρ) |
| 4–2 (0.73) | 1–1 (0.66) | 2–2 (0.76) | 1–3 (0.72) | 1–3 (0.65) | 3–3 (0.69) |
| 11–5 (0.65) | 4–2 (0.69) | 5–10 (0.57) | 6–2 (0.60) | 2–1 (0.59) | 5–1 (0.61) |
| 17–18 (0.64) 1 | 11–10 (0.60) | 18–19 (0.58) 1 | 2–5 (0.64) | 4–8 (0.58) 2 | 7–8 (0.60) 2 |
| 20–20 (0.54) | 17–19 (0.73) 1 | 20–20 (0.52) | 4–7 (0.75) 2 | 6–4 (0.59) | 2–4 (0.57) |
| 9–9 (0.51) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basile, M.; Cauzzo, S.; Callara, A.L.; Montanaro, D.; Hartwig, V.; Morelli, M.S.; Frijia, F.; Giannoni, A.; Passino, C.; Emdin, M.; et al. mICA-Based fMRI Analysis of Specific CO2-Level-Dependent BOLD Signal Changes in the Human Brainstem. Electronics 2023, 12, 290. https://doi.org/10.3390/electronics12020290
Basile M, Cauzzo S, Callara AL, Montanaro D, Hartwig V, Morelli MS, Frijia F, Giannoni A, Passino C, Emdin M, et al. mICA-Based fMRI Analysis of Specific CO2-Level-Dependent BOLD Signal Changes in the Human Brainstem. Electronics. 2023; 12(2):290. https://doi.org/10.3390/electronics12020290
Chicago/Turabian StyleBasile, Miriam, Simone Cauzzo, Alejandro Luis Callara, Domenico Montanaro, Valentina Hartwig, Maria Sole Morelli, Francesca Frijia, Alberto Giannoni, Claudio Passino, Michele Emdin, and et al. 2023. "mICA-Based fMRI Analysis of Specific CO2-Level-Dependent BOLD Signal Changes in the Human Brainstem" Electronics 12, no. 2: 290. https://doi.org/10.3390/electronics12020290