


A Power-Efficient Neuromorphic Digital Implementation of Neural–Glial Interactions †

Abstract
:1. Introduction
2. Dynamic Neuron and Astrocyte Models
2.1. Neuron Model
2.2. Astrocyte Model
2.3. Model of Astrocyte–Neuron Interaction
- : Time delay (s)
- , : Factors responsible for the activation and relaxation of parameter Z
- : Threshold parameter for activation of Z
- v: membrane potential of the pre-synaptic neuron.
3. Materials and Methods
4. Results
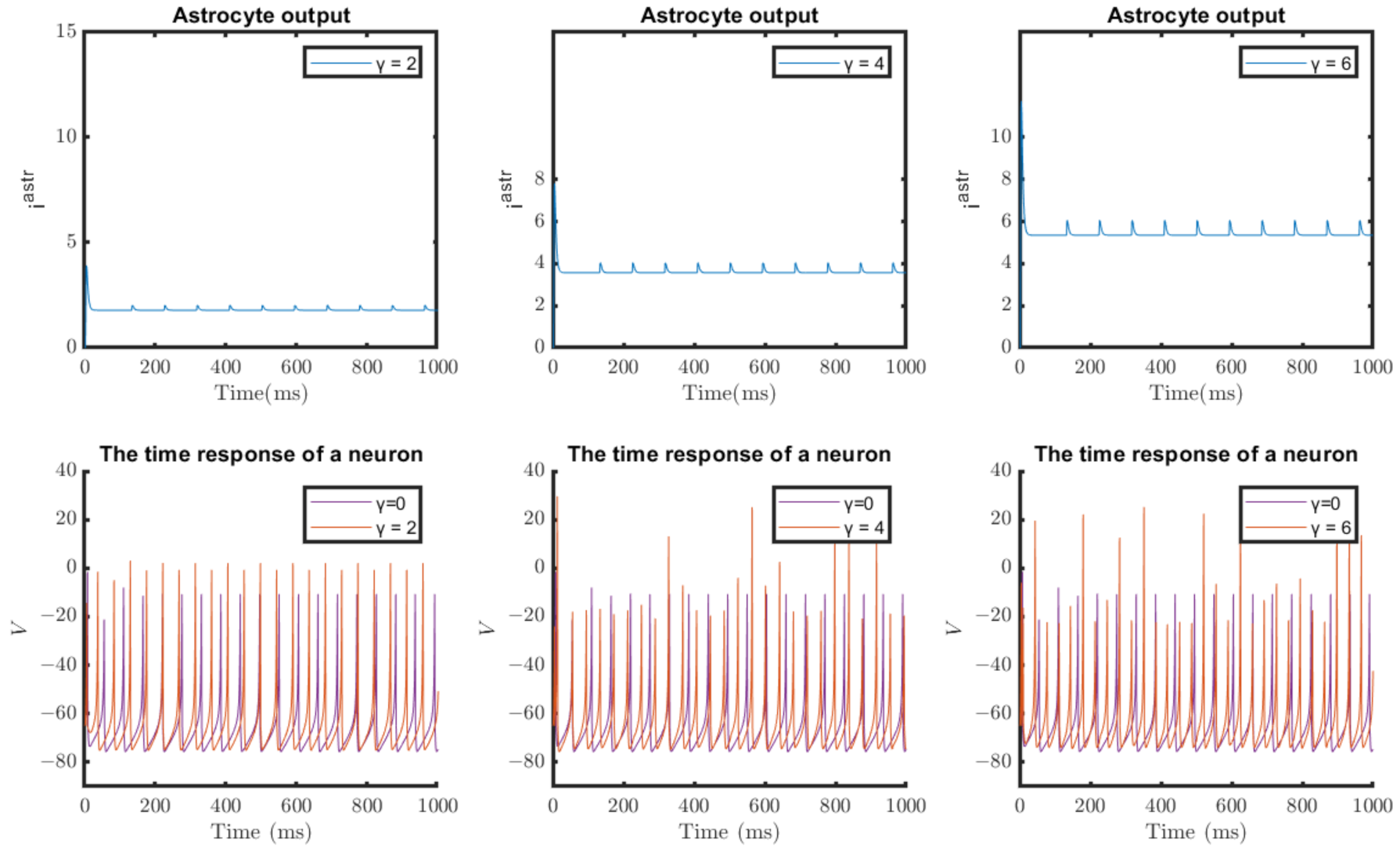
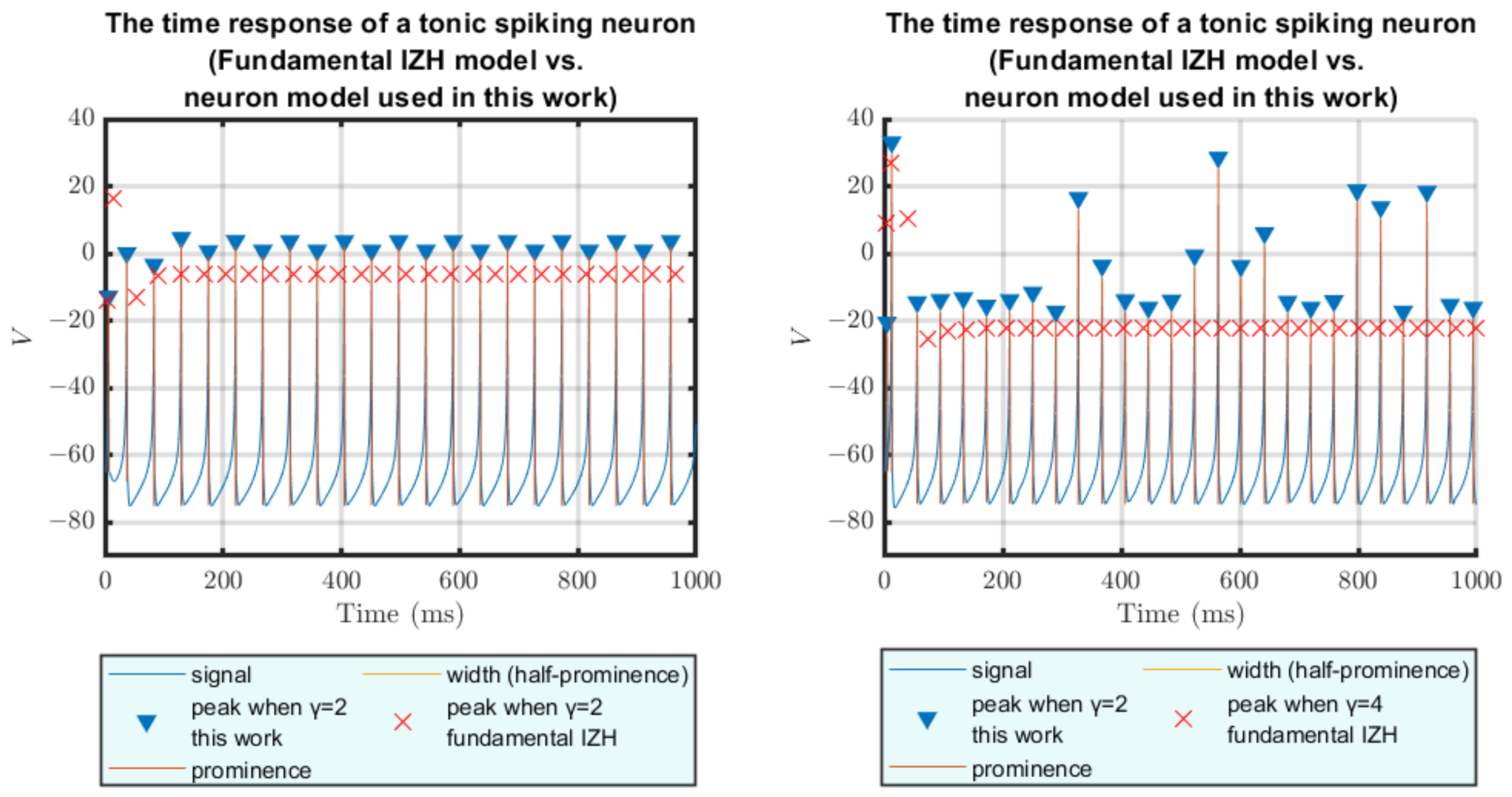
4.1. Results of Software Simulations
4.2. Results of Hardware Implementation
4.3. Error Analysis
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASIC | Application-Specific Integrated Circuit |
| ATP | Adenosine Triphosphate |
| CMOS | Complementary Metal–Oxide Semiconductor |
| DSP | Digital Signal Processor |
| ER | Endoplasmic Reticulum |
| FPGA | Field-Programmable Gate Array |
| GABA | Gamma-aminobutyric acid |
| IZH | Izhikevich Model |
| Inositol 1,4,5 Trisphosphate | |
| Inositol 1,4,5 Trisphosphate Receptors | |
| LUT | LookUp Table |
| mGluR | Metabotropic Glutamate Receptor |
| NMDA-R | N-methyl D-aspartate (NMDA) Receptors |
| ODE | Ordinary Differential Equation |
| RMSE | Root Mean Square Error |
| VHDL | Very High Speed Integrated Circuit Hardware Description Language |
| VLSI | Very Large Scale Integration |
References
- Chen, I.; Lui, F. Neuroanatomy, Neuron Action Potential; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Chrysafides, S.; Bordes, S.J. Physiology, Resting Potential; PublStatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Azevedo, F.A.; Carvalho, L.R.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.; Leite, R.E.; Filho, W.J.; Lent, R.; Herculano-Houzel, S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Cuzzocrea, S. Astrocytes: Role and Functions in Brain Pathologies. Front. Pharmacol. 2019, 10, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, Y.; Zanetti, A.T.; Hallock, R.M. The Role of Astrocytes in the Regulation of Synaptic Plasticity and Memory Formation. Neural Plast. 2013, 10, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haghiri, S.; Naderi, A.; Ghanbari, B.; Ahmadi, A. High Speed and Low Digital Resources Implementation of Hodgkin-Huxley Neuronal Model Using Base-2 Functions. IEEE Trans. Circuits Syst. I Regul. Pap. 2021, 68, 275–287. [Google Scholar] [CrossRef]
- Azad, F.; Shalchian, M.; Amiri, M. Circuit modelling of 2-AG indirect pathway via astrocyte as a catalyst for synaptic self repair. Analog Integr. Circuits Signal Process. 2018, 95. [Google Scholar] [CrossRef]
- Tir, S.; Shalchian, M.; Moezzi, M. Design of bioinspired tripartite synapse analog integrated circuit in 65-nm CMOS Technology. J. Comput. Electron. 2020, 19, 1313–1328. [Google Scholar] [CrossRef]
- Nazari, S.; Amiri, M.; Faez, K.; Amiri, M. Multiplier-less digital implementation of neuron–astrocyte signalling on FPGA. Neurocomputing 2015, 164, 281–292. [Google Scholar] [CrossRef]
- Haghiri, S.; Ahmadi, A.; Saif, M. Complete neuron-astrocyte interaction model: Digital multiplierless design and networking mechanism. IEEE Trans. Biomed. Circuits Syst. 2016, 11, 117–127. [Google Scholar] [CrossRef]
- Faramarzi, F.; Azad, F.; Amiri, M.; Linares-Barranco, B. A neuromorphic digital circuit for neuronal information encoding using astrocytic calcium oscillations. Front. Neurosci. 2019, 13, 998. [Google Scholar] [CrossRef]
- Taylan, O.; Abusurrah, M.; Eftekhari-Zadeh, E.; Nazemi, E.; Bano, F.; Roshani, A. Controlling Effects of Astrocyte on Neuron Behavior in Tripartite Synapse Using VHDL–AMS. Mathematics 2021, 9, 2700. [Google Scholar] [CrossRef]
- Bicaku, A.; Sapounaki, M.; Kakarountas, A. A low-complexity bit-efficient Neuromorphic Astrocyte-Neuron Circuit. In Proceedings of the 2021 28th IEEE International Conference on Electronics, Circuits, and Systems (ICECS), Dubai, United Arab Emirates, 28 November–1 December 2021; pp. 1–6. [Google Scholar]
- Cassidy, A.; Andreou, A.G. Dynamical digital silicon neurons. In Proceedings of the 2008 IEEE Biomedical Circuits and Systems Conference, Baltimore, MD, USA, 20–22 November 2008; pp. 289–292. [Google Scholar] [CrossRef]
- Ambroise, M.; Levi, T.; Joucla, S.; Yvert, B.; Saïghi, S. Real-time biomimetic Central Pattern Generators in an FPGA for hybrid experiments. Front. Neurosci. 2013, 7, 215. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, W.S.; Pitts, W. A logical calculus of the ideas immanent in nervous activity. Bull. Math. Biophys. 1943, 5, 115–133. [Google Scholar] [CrossRef]
- Borisyuk, A.; Rinzel, J. Understanding Neuronal Dynamics by Geometrical Dissection of Minimal Models; Elsevier Science Publisher B.V.: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Izhikevich, E.M. Simple model of spiking neurons. IEEE Trans. Neural Netw. 2003, 14, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Izhikevich, E.M. Which Model to Use for Cortical Spiking Neurons? IEEE Trans. Neural Netw. 2004, 15, 1063–1070. [Google Scholar] [CrossRef]
- Heidarpur, M.; Ahmadi, A.; Ahmadi, M. Time Step Impact on Performance and Accuracy of Izhikevich Neuron: Software Simulation and Hardware Implementation. In Proceedings of the 2020 IEEE International Symposium on Circuits and Systems (ISCAS), Seville, Spain, 12–14 October 2020; pp. 1–5. [Google Scholar] [CrossRef]
- Garaffa, L.C.; Aljuffri, A.; Reinbrecht, C.; Hamdioui, S.; Taouil, M.; Sepulveda, J. Revealing the Secrets of Spiking Neural Networks: The Case of Izhikevich Neuron. In Proceedings of the 2021 24th Euromicro Conference on Digital System Design (DSD), Palermo, Italy, 1–3 September 2021; pp. 514–518. [Google Scholar] [CrossRef]
- Yang, S.; Liu, P.; Xue, J.; Sun, R.; Ying, R. An Efficient FPGA Implementation of Izhikevich Neuron Model. In Proceedings of the 2020 International SoC Design Conference (ISOCC), Yeosu, Republic of Korea, 21–24 October 2020; pp. 141–142. [Google Scholar] [CrossRef]
- Kafraj, M.S.; Parastesh, F.; Jafari, S. Firing patterns of an improved Izhikevich neuron model under the effect of electromagnetic induction and noise. Chaos Solitons Fractals 2020, 137, 109782. [Google Scholar] [CrossRef]
- Wang, J.; Peng, Z.; Zhan, Y.; Li, Y.; Yu, G.; Chong, K.S.; Wang, C. A High-Accuracy and Energy-Efficient CORDIC based Izhikevich Neuron with Error Suppression and Compensation. IEEE Trans. Biomed. Circuits Syst. 2022, 1–14. [Google Scholar] [CrossRef]
- Vivekanandhan, G.; Hamarash, I.I.; Ali, A.M.A.; He, S.; Sun, K. Firing patterns of Izhikevich neuron model under electric field and its synchronization patterns. Eur. Phys. J. Spec. Top. 2022, 231, 4017–4023. [Google Scholar] [CrossRef]
- Cassidy, A.; Denham, S.; Kanold, P.; Andreou, A. FPGA Based Silicon Spiking Neural Array. In Proceedings of the 2007 IEEE Biomedical Circuits and Systems Conference, Montreal, QC, Canada, 27–30 November 2007; pp. 75–78. [Google Scholar] [CrossRef]
- Terman, D.; Rubin, J.E.; Yew, A.; Wilson, C. Activity patterns in a model for the subthalamopallidal network of the basal ganglia. J. Neurosci. 2002, 22, 2963–2976. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, S.; Jung, P. Modeling synaptic transmission of the tripartite synapse. Phys. Biol. 2007, 4, 1. [Google Scholar] [CrossRef]
- De Pittà, M.; Goldberg, M.; Volman, V.; Berry, H.; Ben-Jacob, E. Glutamate regulation of calcium and IP3 oscillating and pulsating dynamics in astrocytes. J. Biol. Phys. 2009, 35, 383–411. [Google Scholar] [CrossRef] [Green Version]
- Oschmann, F.; Berry, H.; Obermayer, K.; K, L. From in silico astrocyte cell models to neuron-astrocyte network models: A review. Brain Res. Bull. 2019, 35, 383–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manninen, T.; Havela, R.; Linne, M.a. Computational Models for Calcium-Mediated Astrocyte Functions. Front. Comput. Neurosci. 2018, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postnov, D.E.; Brazhe, N.A.; Sosnovtseva, O.V. Functional modeling of neural-glial interaction. In Biosimulation in Biomedical Research, Health Care and Drug Development; Springer: Vienna, Austria, 2011; pp. 133–151. [Google Scholar]
- Keener, J.; Sneyd, J. Mathematical Physiology: II: Systems Physiology; Springer: Vienna, Austria, 2009. [Google Scholar]
- Araque, A.; Sanzgiri, R.P.; Parpura, V.; Haydon, P.G. Calcium elevation in astrocytes causes an NMDA receptor-dependent increase in the frequency of miniature synaptic currents in cultured hippocampal neurons. J. Neurosci. 1998, 18, 6822–6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, R.C.; Parpura, V. Mitochondria Modulate Ca2+-Dependent Glutamate Release from Rat Cortical Astrocytes. J. Neurosci. 2008, 28, 9682–9691. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, S.; Jung, P. Dressed neurons: Modeling neural–glial interactions. Phys. Biol. 2004, 1, 35. [Google Scholar] [CrossRef]
- Volman, V.; Ben-Jacob, E.; Levine, H. The astrocyte as a gatekeeper of synaptic information transfer. Neural Comput. 2007, 19, 303–326. [Google Scholar] [CrossRef]
- Wade, J.J.; McDaid, L.J.; Harkin, J.; Crunelli, V.; Kelso, J.S.; Beiu, V. Exploring retrograde signaling via astrocytes as a mechanism for self repair. In Proceedings of the The 2011 International Joint Conference on Neural Networks, San Jose, CA, USA, 31 July–5 August 2011; pp. 3149–3155. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Operators | This Work | Bicaku et al. [13] | Nazari et al. [9] |
|---|---|---|---|
| Adders | 7 | 12 | 14 |
| Adder/Subtractors | 10 | 11 | 0 |
| Subtractors | 0 | 0 | 6 |
| Comparators | 2 | 1 | 2 |
| Multiplexers | 3 | 3 | 3 |
| Multipliers | 2 | 3 | 0 |
| Shifters1 | 12 | 16 | 15 |
| Parameters | Values for Tonic Spiking Behavior | Values for Tonic Bursting Behavior |
|---|---|---|
| a | 1/64 | 1/64 |
| b | 0.156250 | 0.234375 |
| c | −50.508 | −39.063 |
| d | 6.2500 | 3.9062 |
| 10.9375 mA | 0.58594 mA | |
| 30 mV | 30 mV | |
| −65 mV | −65 mV | |
| −10.1562 mV | −10.1562 mV |
| Parameters | Values |
|---|---|
| 0.0722 µM | |
| 0.16 µM | |
| 0.2 | |
| 0.02 |
| = 0 | = 2 | = 4 | |||||
|---|---|---|---|---|---|---|---|
| This Work | This Work | Glion+IZH Neuron [18] | Nazari et al. [9] | This Work | Glion+IZH Neuron [18] | Nazari et al. [9] | |
| # spikes | 19 | 22 | 27 | 26 | 27 | 32 | 30 |
| Accuracy | Operation Frequency | Area | Power | ||||||
|---|---|---|---|---|---|---|---|---|---|
| bit | MHz | Slice LUTs | Slice Registers | DSPs | LUTRAM | Static (W) | Dynamic (W) | Total (W) | |
| Nazari et al. [9] | 20 | 318 | 856 | 838 | 5 | - | - | - | - |
| Bicaku et al. [13] | 32 | 120 | 826 | 1615 | 4 | 7 | 0.107 | 0.194 | 0.301 |
| Bicaku et al. [13] 1 | 20 | 120 | 397 | 985 | 3 | 7 | 0.106 | 0.112 | 0.218 |
| this work | 20 | 124 | 324 | 531 | 2 | 0 | 0.104 | 0.065 | 0.169 |
| Area | Power | ||||||
|---|---|---|---|---|---|---|---|
| Slice LUTs | Slice Registers | DSPs | LUTRAM | Static (W) | Dynamic (W) | Total (W) | |
| Nazari et al. [9] | 164.20% | 36.63% | 150% | - | - | - | - |
| Bicaku et al. [13] | 22.53% | 46.09% | 50% | - | 0.94% | 72.31% | 28.07% |
| Spiking Activity | ||||||
|---|---|---|---|---|---|---|
| = 0 | = 2 | = 4 | ||||
| 10.10 | 16.16 | 10.10 | 16.16 | 10.10 | 16.16 | |
| V | 0.270683 | 0.005765 | 1.197075 | 0.082194 | 2.626134 | 0.115209 |
| U | 0.001322 | 0.000026 | 0.037754 | 0.000955 | 1.648498 | 0.001562 |
| Gm | 0.008915 | 0.000573 | 0.0079326 | 0.000563 | 0.060797 | 0.000556 |
| Sm | 0.000550 | 0.000098 | 0.003438 | 0.000010 | 0.007438 | 0.000010 |
| Bursting Activity | ||||||
|---|---|---|---|---|---|---|
| = 0 | = 2 | = 4 | ||||
| 10.10 | 16.16 | 10.10 | 16.16 | 10.10 | 16.16 | |
| V | 0.054521 | 0.001111 | 0.559855 | 0.049529 | 0.920400 | 0.065973 |
| U | 0.000806 | 0.000027 | 0.013021 | 0.000824 | 0.026106 | 0.00138 |
| Gm | 0.009579 | 0.000559 | 0.011099 | 0.000532 | 0.050347 | 0.000522 |
| Sm | 0.000549 | 0.000010 | 0.000541 | 0.000010 | 0.004702 | 0.000010 |
| Spiking Activity | ||||||
|---|---|---|---|---|---|---|
| = 0 | = 2 | = 4 | ||||
| Nazari et al. [9] | This Work | Nazari et al. [9] | This Work | Nazari et al. [9] | This Work | |
| V | 59.640823 | 13.757812 | 59.569458 | 15.894531 | 59.018311 | 21.773438 |
| U | 5.263519 | 9.136719 | 5.337692 | 7.955078 | 5.033951 | 7.068359 |
| Gm | 0.136078 | 0.000000 | 0.137466 | 0.035156 | 0.128769 | 0.068359 |
| Sm | 0.005356 | 0.000000 | 0.004883 | 0.003906 | 0.007401 | 0.005859 |
| Bursting Activity | ||||||
|---|---|---|---|---|---|---|
| = 0 | = 2 | = 4 | ||||
| Nazari et al. [9] | This Work | Nazari et al. [9] | This Work | Nazari et al. [9] | This Work | |
| V | 59.410980 | 33.068359 | 58.209061 | 31.195312 | 57.547958 | 34.619141 |
| U | 7.648270 | 11.839844 | 7.826736 | 11.833008 | 8.773743 | 13.263672 |
| Gm | 0.132126 | 0.129883 | 0.127670 | 0.085938 | 0.123352 | 0.059570 |
| Sm | 0.006546 | 0.009766 | 0.007462 | 0.006836 | 0.008362 | 0.005859 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bicaku, A.; Sapounaki, M.; Kakarountas, A.; Tasoulis, S.K. A Power-Efficient Neuromorphic Digital Implementation of Neural–Glial Interactions. J. Low Power Electron. Appl. 2023, 13, 10. https://doi.org/10.3390/jlpea13010010
Bicaku A, Sapounaki M, Kakarountas A, Tasoulis SK. A Power-Efficient Neuromorphic Digital Implementation of Neural–Glial Interactions. Journal of Low Power Electronics and Applications. 2023; 13(1):10. https://doi.org/10.3390/jlpea13010010
Chicago/Turabian StyleBicaku, Angeliki, Maria Sapounaki, Athanasios Kakarountas, and Sotiris K. Tasoulis. 2023. "A Power-Efficient Neuromorphic Digital Implementation of Neural–Glial Interactions" Journal of Low Power Electronics and Applications 13, no. 1: 10. https://doi.org/10.3390/jlpea13010010