
Assessing and Modelling of Post-Traumatic Stress Disorder Using Molecular and Functional Biomarkers
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Catalytic Factors Associated with PTSD
2.1. Oxidative Stress
2.2. Genes Variations Related to HPA Axis
2.3. Neurotransmitter Imbalances and Related Genes Variations
2.4. Inflammatory System Candidate Gene Variations
2.5. G Protein-Coupled Receptor (GPCR) System Candiate Gene Variations
2.6. Other Genes Variations
2.7. GWAS Studies

{kind=link}
{kind=link}
{kind=link}
| Type | Gene | Variation | Ref. |
|---|---|---|---|
| Hypothalamic-pituitary-adrenal axis | NR3C1 | Bcl-1 (rs41423247) ER22/23EK (rs6189/rs6190) 9β SNP (rs6198) rs258747 | [17] [21] [21] [22] |
| FKBP5 | rs3800373 rs9296158 rs1360780 rs9470080 | [22] [22] [22] [22] | |
| CRHR1 | rs110402 rs12938031 rs4792887 rs12944712 | [24] [24] [25] [25] | |
| CRHR2 | rs2267715 | [26] | |
| Serotonergic system | SLC6A4 | 5-HTTLPR/rs25531 rs16965628 | [30] [70] |
| HTR2A | rs7997012 rs977003 rs7322347 | [5] [5] [5] | |
| TPH2 | rs11178997T | [32] | |
| Dopaminergic system | DRD2/ANKK1 | DRD2/ANKK1-Taq1A (rs1800497) | [71] |
| DRD3 | rs2134655 rs201252087 rs4646996 rs9868039 rs2251177 | [72] [72] [72] [72] [72] | |
| DRD4 | VNTR 7R | [44] | |
| SLC6A3 | 3′VNTR9r | [45] | |
| DBH | rs1611115 | [46] | |
| COMT | rs4680 rs4633C | [44] [32] | |
| GABAergic 1 system | GABRA2 | Unspecified | [36] |
| Endocannabinoid system | SKA2 | rs7208505 | [37] |
| CNR1 | G1359A (rs1049353) rs806371 | [38] [39] | |
| Inflammatory system | CRP | rs1130864 | [48] |
| IL1B | rs1143633C and rs16944A | [49] | |
| TNFα | rs1800629 | [50] | |
| GPCR 2 system | RGS2 | rs4606 | [51] |
| ADCYAP1 | rs2267735 | [52] | |
| ADRB2 | rs2400707 | [53] | |
| Other | MAOB | rs1799836 | [44] |
| NPY | rs16147 | [56] | |
| APOE | ε4 ε2 | [57] [58] | |
| OXTR | rs53576 | [59] | |
| FAAH | rs324420 | [39] | |
| PPM1F | rs9610608 | [61] | |
| SLC18A2 | rs363276 | [62] | |
| OPRL1 | rs6010719 | [63] | |
| PRKCA | rs4790904 | [64] | |
| Brain-derived neurotrophic factor | BDNF | val66met (rs6265) | [73] |
| Mitochondrial | MT-ATP8 | mt8414C → T | [65] |
| MT-ND5 | mt12501G → A | [65] | |
| RNA regulation | LINC01090 | rs10170218 | [66] |
| GWAS 3 studies | RORA | rs8042149 | [74] |
| PRTFDC1 | rs6482463 | [68] | |
| TLL1 | rs6812849 rs7691872 rs406001 | [69] [69] [69] | |
| ANKRD55 | rs159572 | [70] | |
| ZNF626 | rs11085374 | [70] |
3. Exploring the Molecular Landscape: Gene Ontology and Network Analyses of Key PTSD-Associated Genes
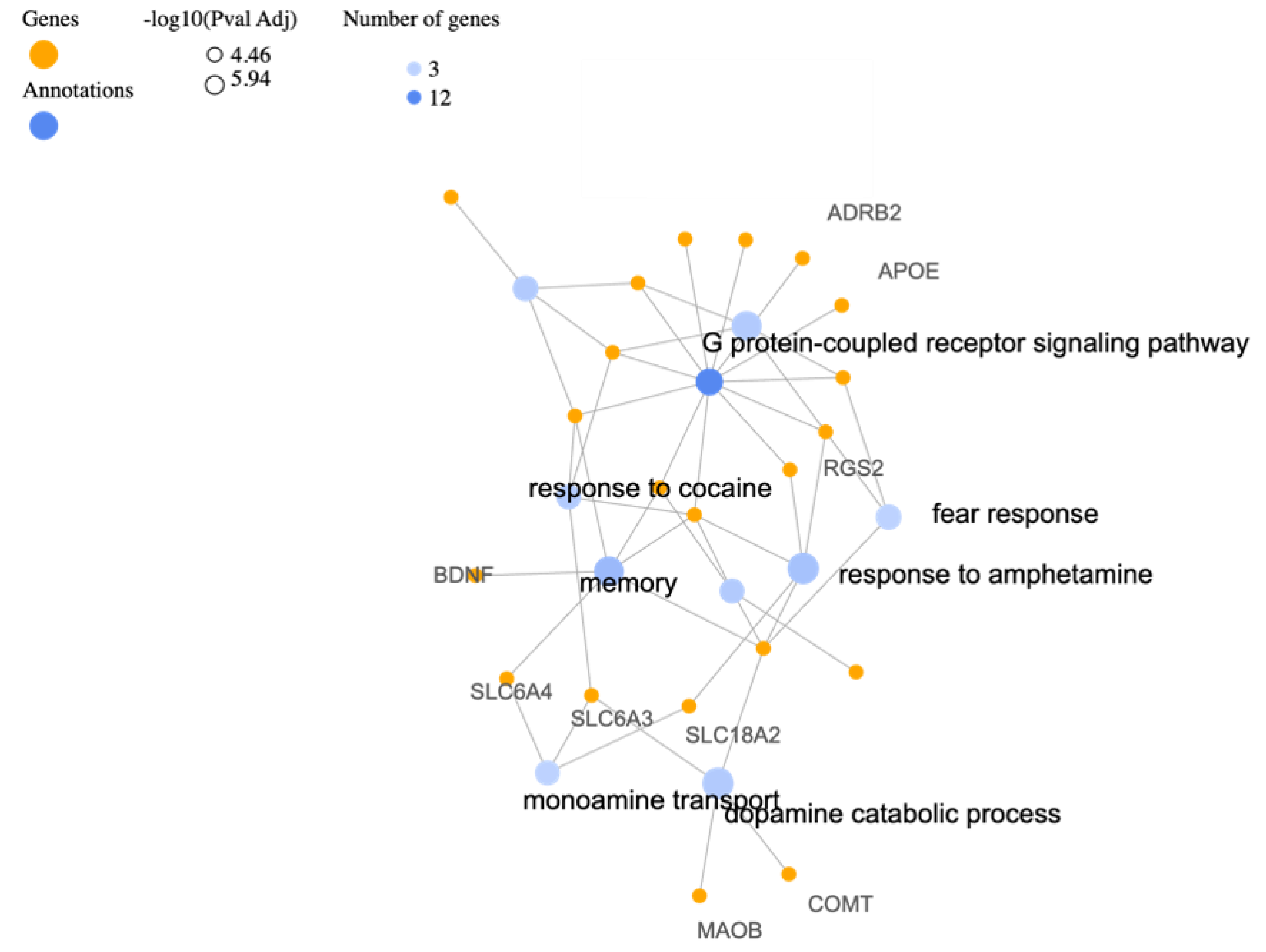
3.1. Biological Processes
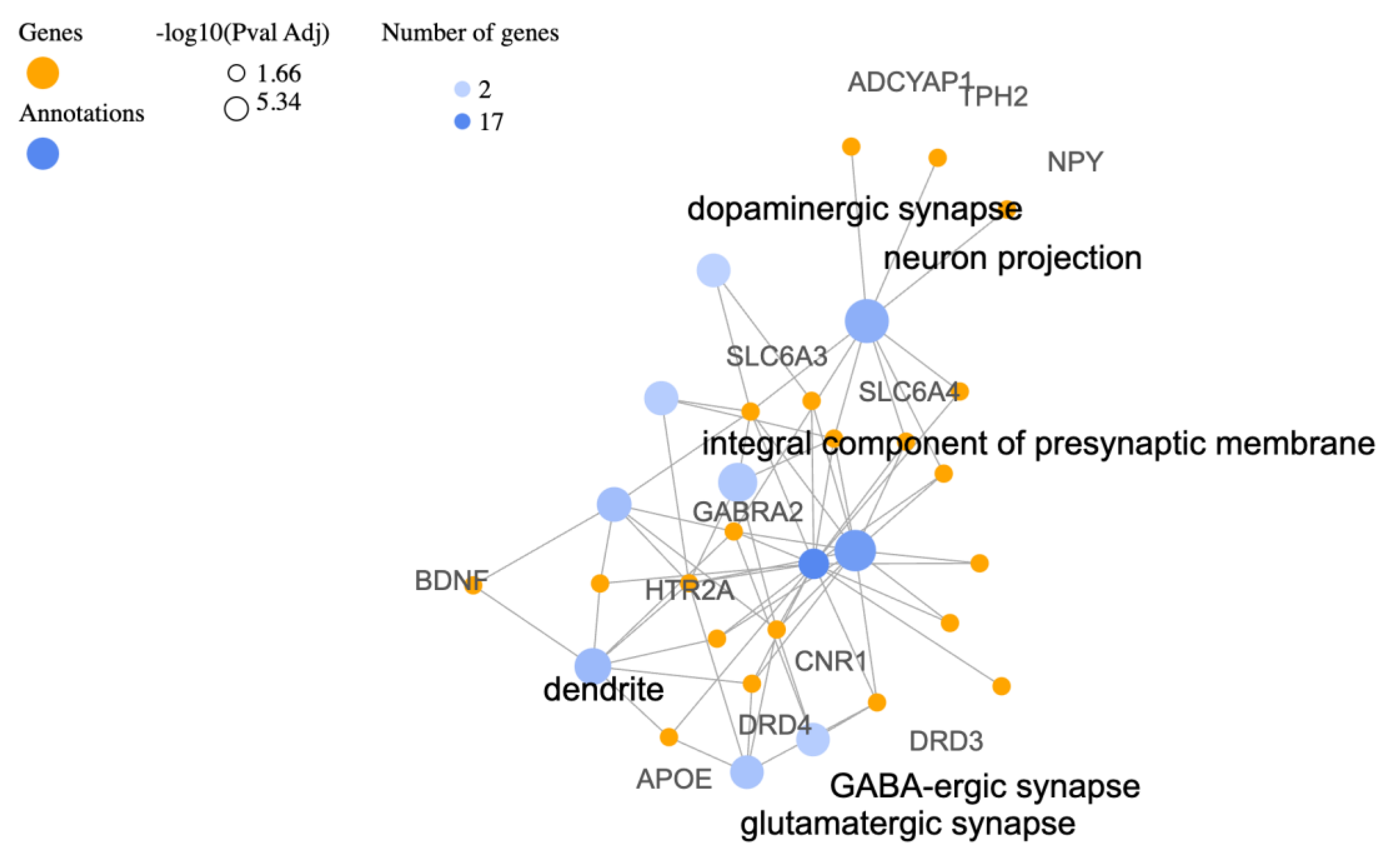
3.2. Cellular Components (CC)
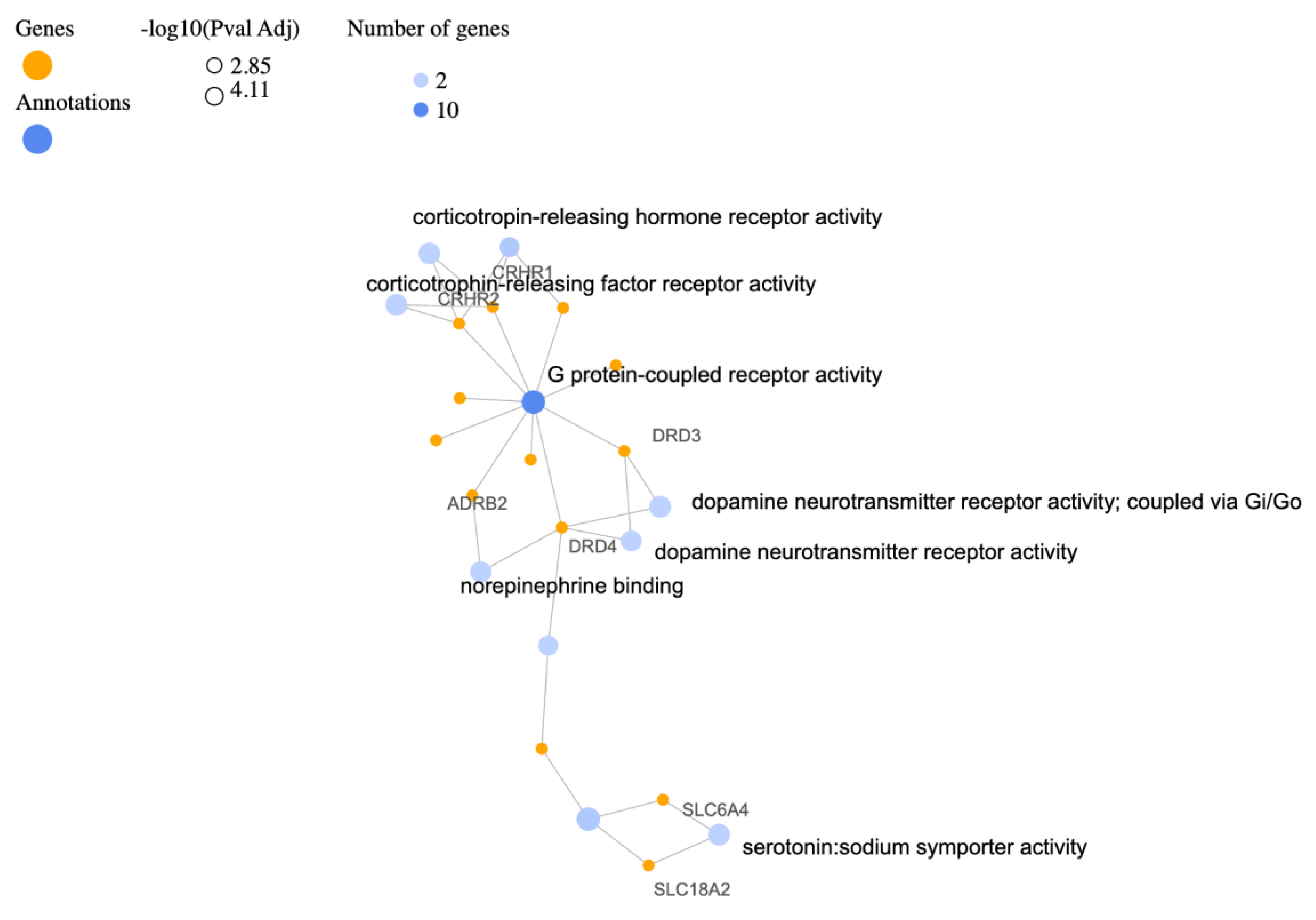
3.3. Molecular Functions (MF)
4. Proposed Function for Catalytic Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Banerjee, S.B.; Morrison, F.G.; Ressler, K.J. Genetic approaches for the study of PTSD: Advances and challenges. Neurosci. Lett. 2017, 649, 139–146. [Google Scholar]
- Dunlop, B.W.; Wong, A. The hypothalamic-pituitary-adrenal axis in PTSD: Pathophysiology and treatment interventions. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 89, 361–379. [Google Scholar]
- Rasmusson, A.M.; Pineles, S.L. Neurotransmitter, Peptide, and Steroid Hormone Abnormalities in PTSD: Biological Endophenotypes Relevant to Treatment. Curr. Psychiatry Rep. 2018, 20, 52. [Google Scholar]
- Peruzzolo, T.L.; Pinto, J.V.; Roza, T.H.; Shintani, A.O.; Anzolin, A.P.; Gnielka, V.; Kohmann, A.M.; Marin, A.S.; Lorenzon, V.R.; Brunoni, A.R.; et al. Inflammatory and oxidative stress markers in post-traumatic stress disorder: A systematic review and meta-analysis. Mol. Psychiatry 2022, 27, 3150–3163. [Google Scholar]
- Miller, M.W.; Lin, A.P.; Wolf, E.J.; Miller, D.R. Oxidative Stress, Inflammation, and Neuroprogression in Chronic PTSD. Harv. Rev. Psychiatry 2018, 26, 57–69. [Google Scholar]
- Daskalakis, N.P.; Rijal, C.M.; King, C.; Huckins, L.M.; Ressler, K.J. Recent Genetics and Epigenetics Approaches to PTSD. Curr. Psychiatry Rep. 2018, 20, 30. [Google Scholar] [CrossRef]
- Smoller, J.W. The Genetics of Stress-Related Disorders: PTSD, Depression, and Anxiety Disorders. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2016, 41, 297–319. [Google Scholar]
- Jiang, M.; Jia, K.; Wang, L.; Li, W.; Chen, B.; Liu, Y.; Wang, H.; Zhao, S.; He, Y.; Zhou, C. Alterations of DNA damage repair in cancer: From mechanisms to applications. Ann. Transl. Med. 2020, 8, 1685. [Google Scholar]
- Atli, A.; Bulut, M.; Bez, Y.; Kaplan, İ.; Özdemir, P.G.; Uysal, C.; Selçuk, H.; Sir, A. Altered lipid peroxidation markers are related to post-traumatic stress disorder (PTSD) and not trauma itself in earthquake survivors. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 266, 329–336. [Google Scholar]
- Maier, A.; Dharan, A.; Oliver, G.; Berk, M.; Redston, S.; Back, S.E.; Kalivas, P.; Ng, C.; Kanaan, R.A. A multi-centre, double-blind, 12-week, randomized, placebo-controlled trial to assess the efficacy of adjunctive N-Acetylcysteine for treatment-resistant PTSD: A study protocol. BMC Psychiatry 2020, 20, 397. [Google Scholar]
- Dell’Oste, V.; Fantasia, S.; Gravina, D.; Palego, L.; Betti, L.; Dell’Osso, L.; Giannaccini, G.; Carmassi, C. Metabolic and Inflammatory Response in Post-Traumatic Stress Disorder (PTSD): A Systematic Review on Peripheral Neuroimmune Biomarkers. Int. J. Environ. Res. Public Health 2023, 20, 2937. [Google Scholar]
- Ramos-Cejudo, J.; Genfi, A.; Abu-Amara, D.; Debure, L.; Qian, M.; Laska, E.; Siegel, C.; Milton, N.; Newman, J.; Blessing, E.; et al. CRF serum levels differentiate PTSD from healthy controls and TBI in military veterans. Psychiatr. Res. Clin. Pract. 2021, 3, 153–162. [Google Scholar]
- D’elia, A.T.D.; Juruena, M.F.; Coimbra, B.M.; Mello, M.F.; Mello, A.F. Posttraumatic stress disorder (PTSD) and depression severity in sexually assaulted women: Hypothalamic-pituitary-adrenal (HPA) axis alterations. BMC Psychiatry 2021, 21, 174. [Google Scholar]
- Danan, D.; Todder, D.; Zohar, J.; Cohen, H. Is PTSD-Phenotype Associated with HPA-Axis Sensitivity? Feedback Inhibition and Other Modulating Factors of Glucocorticoid Signaling Dynamics. Int. J. Mol. Sci. 2021, 22, 6050. [Google Scholar]
- Szeszko, P.R.; Lehrner, A.; Yehuda, R. Glucocorticoids and Hippocampal Structure and Function in PTSD. Harv. Rev. Psychiatry 2018, 26, 142–157. [Google Scholar]
- Speer, K.E.; Semple, S.; Naumovski, N.; D’Cunha, N.M.; McKune, A.J. HPA axis function and diurnal cortisol in post-traumatic stress disorder: A systematic review. Neurobiol. Stress 2019, 11, 100180. [Google Scholar]
- Fischer, S.; Schumacher, T.; Knaevelsrud, C.; Ehlert, U.; Schumacher, S. Genes and hormones of the hypothalamic-pituitary-adrenal axis in post-traumatic stress disorder. What is their role in symptom expression and treatment response? J. Neural Transm. 2021, 128, 1279–1286. [Google Scholar]
- Azadi, S.; Azarpira, N.; Roozbeh, J.; Ezzatzadegan-Jahromi, S.; Raees-Jalali, G.A.; Foroughinia, F.; Karimzadeh, I. Genetic polymorphisms of glucocorticoid receptor and their association with new-onset diabetes mellitus in kidney transplant recipients. Gene 2023, 856, 147138. [Google Scholar]
- Mottaghi, S.; Sagheb, M.M.; Azarpira, N.; Abdizadeh, F.; Faeghi, R.; Karimzadeh, I. Association between the Three Polymorphisms of the Glucocorticoid Receptor Gene and the Early Clinical Outcome in Kidney Transplantation Patients. Iran. J. Med. Sci. 2021, 46, 444. [Google Scholar]
- Zoladz, P.R.; Duffy, T.J.; Mosley, B.E.; Fiely, M.K.; Nagle, H.E.; Scharf, A.R.; Brown, C.M.; Earley, M.B.; Rorabaugh, B.R.; Dailey, A.M. Interactive influence of sex, stressor timing, and the BclI glucocorticoid receptor polymorphism on stress-induced alterations of long-term memory. Acute Stress Mem. Brain 2019, 133, 72–83. [Google Scholar]
- Castro-Vale, I.; Durães, C.; van Rossum, E.F.C.; Staufenbiel, S.M.; Severo, M.; Lemos, M.C.; Carvalho, D. The Glucocorticoid Receptor Gene (NR3C1) 9β SNP Is Associated with Posttraumatic Stress Disorder. Healthcare 2021, 9, 173. [Google Scholar]
- Sheerin, C.M.; Lind, M.J.; Bountress, K.E.; Marraccini, M.E.; Amstadter, A.B.; Bacanu, S.-A.; Nugent, N.R. Meta-Analysis of Associations Between Hypothalamic-Pituitary-Adrenal Axis Genes and Risk of Posttraumatic Stress Disorder. J. Trauma. Stress 2020, 33, 688–698. [Google Scholar]
- Wang, L.; Zhang, J.; Li, G.; Cao, C.; Fang, R.; Liu, P.; Luo, S.; Zhao, G.; Zhang, Y.; Zhang, K. The ADCYAP1R1 Gene Is Correlated With Posttraumatic Stress Disorder Symptoms Through Diverse Epistases in a Traumatized Chinese Population. Front. Psychiatry 2021, 12, 665599. [Google Scholar]
- Pape, J.C.; Carrillo-Roa, T.; Rothbaum, B.O.; Nemeroff, C.B.; Czamara, D.; Zannas, A.S.; Iosifescu, D.; Mathew, S.J.; Neylan, T.C.; Mayberg, H.S.; et al. DNA methylation levels are associated with CRF(1) receptor antagonist treatment outcome in women with post-traumatic stress disorder. Clin. Epigenet. 2018, 10, 136. [Google Scholar]
- White, S.; Acierno, R.; Ruggiero, K.J.; Koenen, K.C.; Kilpatrick, D.G.; Galea, S.; Gelernter, J.; Williamson, V.; McMichael, O.; Vladimirov, V.I.; et al. Association of CRHR1 variants and posttraumatic stress symptoms in hurricane exposed adults. J. Anxiety Disord. 2013, 27, 678–683. [Google Scholar]
- Zhang, K.; Wang, L.; Li, G.; Cao, C.; Fang, R.; Liu, P.; Luo, S.; Zhang, X. Correlation between hypothalamic-pituitary-adrenal axis gene polymorphisms and posttraumatic stress disorder symptoms. Horm. Behav. 2020, 117, 104604. [Google Scholar]
- Ogłodek, E.A. Changes in the Serum Concentration Levels of Serotonin, Tryptophan and Cortisol among Stress-Resilient and Stress-Susceptible Individuals after Experiencing Traumatic Stress. Int. J. Environ. Res. Public Health 2022, 19, 16517. [Google Scholar]
- Hendrickson, R.C.; Raskind, M.A.; Millard, S.P.; Sikkema, C.; Terry, G.E.; Pagulayan, K.F.; Li, G.; Peskind, E.R. Evidence for altered brain reactivity to norepinephrine in Veterans with a history of traumatic stress. Neurobiol. Stress 2018, 8, 103–111. [Google Scholar]
- Aliev, G.; Beeraka, N.M.; Nikolenko, V.N.; Svistunov, A.A.; Rozhnova, T.; Kostyuk, S.; Cherkesov, I.; Gavryushova, L.V.; Chekhonatsky, A.A.; Mikhaleva, L.M.; et al. Neurophysiology and Psychopathology Underlying PTSD and Recent Insights into the PTSD Therapies—A Comprehensive Review. J. Clin. Med. 2020, 9, 2951. [Google Scholar]
- Bishop, J.R.; Lee, A.M.; Mills, L.J.; Thuras, P.D.; Eum, S.; Clancy, D.; Erbes, C.R.; Polusny, M.A.; Lamberty, G.J.; Lim, K.O. Methylation of FKBP5 and SLC6A4 in Relation to Treatment Response to Mindfulness Based Stress Reduction for Posttraumatic Stress Disorder. Front. Psychiatry 2018, 9, 418. [Google Scholar]
- Miller, M.W.; Sperbeck, E.; Robinson, M.E.; Sadeh, N.; Wolf, E.J.; Hayes, J.P.; Logue, M.; Schichman, S.A.; Stone, A.; Milberg, W.; et al. 5-HT2A Gene Variants Moderate the Association between PTSD and Reduced Default Mode Network Connectivity. Front. Neurosci. 2016, 10, 299. [Google Scholar]
- Goenjian, A.K.; Noble, E.P.; Steinberg, A.M.; Walling, D.P.; Stepanyan, S.T.; Dandekar, S.; Bailey, J.N. Association of COMT and TPH-2 genes with DSM-5 based PTSD symptoms. J. Affect. Disord. 2015, 172, 472–478. [Google Scholar]
- Swanberg, K.M.; Campos, L.; Abdallah, C.G.; Juchem, C. Proton Magnetic Resonance Spectroscopy in Post-Traumatic Stress Disorder-Updated Systematic Review and Meta-Analysis. Chronic Stress 2022, 6. [Google Scholar] [CrossRef]
- Huang, J.; Xu, F.; Yang, L.; Tuolihong, L.; Wang, X.; Du, Z.; Zhang, Y.; Yin, X.; Li, Y.; Lu, K.; et al. Involvement of the GABAergic system in PTSD and its therapeutic significance. Front. Mol. Neurosci. 2023, 16, 1052288. [Google Scholar]
- Seidemann, R.; Duek, O.; Jia, R.; Levy, I.; Harpaz-Rotem, I. The Reward System and Post-Traumatic Stress Disorder: Does Trauma Affect the Way We Interact With Positive Stimuli? Chronic Stress 2021, 5. [Google Scholar] [CrossRef]
- Nisar, S.; Bhat, A.A.; Hashem, S.; Syed, N.; Yadav, S.K.; Uddin, S.; Fakhro, K.; Bagga, P.; Thompson, P.; Reddy, R.; et al. Genetic and Neuroimaging Approaches to Understanding Post-Traumatic Stress Disorder. Int. J. Mol. Sci. 2020, 21, 4503. [Google Scholar]
- Boks, M.P.; Rutten, B.P.F.; Geuze, E.; Houtepen, L.C.; Vermetten, E.; Kaminsky, Z.; Vinkers, C.H. SKA2 Methylation is Involved in Cortisol Stress Reactivity and Predicts the Development of Post-Traumatic Stress Disorder (PTSD) After Military Deployment. Neuropsychopharmacology 2016, 41, 1350–1356. [Google Scholar]
- Korem, N.; Duek, O.; Xu, K.; Harpaz-Rotem, I.; Pietrzak, R.H. Cannabinoid Receptor 1 rs1049353 Variant, Childhood Abuse, and the Heterogeneity of PTSD Symptoms: Results From the National Health and Resilience in Veterans Study. Chronic Stress 2021, 5. [Google Scholar] [CrossRef]
- de Roon-Cassini, T.A.; Bergner, C.L.; Chesney, S.A.; Schumann, N.R.; Lee, T.S.; Brasel, K.J.; Hillard, C.J. Circulating endocannabinoids and genetic polymorphisms as predictors of posttraumatic stress disorder symptom severity: Heterogeneity in a community-based cohort. Transl. Psychiatry 2022, 12, 48. [Google Scholar]
- Aupperle, R.L.; Melrose, A.J.; Stein, M.B.; Paulus, M.P. Executive function and PTSD: Disengaging from trauma. Neuropharmacology 2012, 62, 686–694. [Google Scholar]
- Pitman, R.K.; Rasmusson, A.M.; Koenen, K.C.; Shin, L.M.; Orr, S.P.; Gilbertson, M.W.; Milad, M.R.; Liberzon, I. Biological studies of post-traumatic stress disorder. Nat. Reviews. Neurosci. 2012, 13, 769–787. [Google Scholar]
- Niu, Y.-M.; Zhang, J.; Tang, H.; Cao, L.-H.; Jiang, T.-Y.; Hu, Y.-Y. Association between DRD2/ANKK1 rs1800497 C > T polymorphism and post-traumatic stress disorder susceptibility: A multivariate meta-analysis. Front. Neurosci. 2023, 17, 1102573. [Google Scholar]
- Magwai, T.; Xulu, K.R. Physiological Genomics Plays a Crucial Role in Response to Stressful Life Events, the Development of Aggressive Behaviours, and Post-Traumatic Stress Disorder (PTSD). Genes 2022, 13, 300. [Google Scholar]
- Reith ME, A.; Kortagere, S.; Wiers, C.E.; Sun, H.; Kurian, M.A.; Galli, A.; Volkow, N.D.; Lin, Z. The dopamine transporter gene SLC6A3: Multidisease risks. Mol. Psychiatry 2022, 27, 1031–1046. [Google Scholar]
- Mustapic, M.; Maihofer, A.X.; Mahata, M.; Chen, Y.; Baker, D.G.; O’Connor, D.T.; Nievergelt, C.M. The catecholamine biosynthetic enzyme dopamine β-hydroxylase (DBH): First genome-wide search positions trait-determining variants acting additively in the proximal promoter. Hum. Mol. Genet. 2014, 23, 6375–6384. [Google Scholar]
- Al Jowf, G.I.; Ahmed, Z.T.; Reijnders, R.A.; de Nijs, L.; Eijssen, L.M.T. To Predict, Prevent, and Manage Post-Traumatic Stress Disorder (PTSD): A Review of Pathophysiology, Treatment, and Biomarkers. Int. J. Mol. Sci. 2023, 24, 5238. [Google Scholar]
- Michopoulos, V.; Rothbaum, A.O.; Jovanovic, T.; Almli, L.M.; Bradley, B.; Rothbaum, B.O.; Gillespie, C.F.; Ressler, K.J. Association of CRP genetic variation and CRP level with elevated PTSD symptoms and physiological responses in a civilian population with high levels of trauma. Am. J. Psychiatry 2015, 172, 353–362. [Google Scholar]
- Hovhannisyan, L.; Stepanyan, A.; Arakelyan, A. Genetic variability of interleukin-1 beta as prospective factor from developing post-traumatic stress disorder. Immunogenetics 2017, 69, 703–708. [Google Scholar]
- Bruenig, D.; Mehta, D.; Morris, C.P.; Harvey, W.; Lawford, B.; Young, R.M.; Voisey, J. Genetic and serum biomarker evidence for a relationship between TNFα and PTSD in Vietnam war combat veterans. Compr. Psychiatry 2017, 74, 125–133. [Google Scholar]
- Dunn, E.C.; Solovieff, N.; Lowe, S.R.; Gallagher, P.J.; Chaponis, J.; Rosand, J.; Koenen, K.C.; Waters, M.C.; Rhodes, J.E.; Smoller, J.W. Interaction between genetic variants and exposure to Hurricane Katrina on post-traumatic stress and post-traumatic growth: A prospective analysis of low income adults. J. Affect. Disord. 2014, 152–154, 243–249. [Google Scholar]
- Lind, M.J.; Marraccini, M.E.; Sheerin, C.M.; Bountress, K.; Bacanu, S.-A.; Amstadter, A.B.; Nugent, N.R. Association of Posttraumatic Stress Disorder With rs2267735 in the ADCYAP1R1 Gene: A Meta-Analysis. J. Trauma. Stress 2017, 30, 389–398. [Google Scholar]
- Liberzon, I.; King, A.P.; Ressler, K.J.; Almli, L.M.; Zhang, P.; Ma, S.T.; Cohen, G.H.; Tamburrino, M.B.; Calabrese, J.R.; Galea, S. Interaction of the ADRB2 Gene Polymorphism With Childhood Trauma in Predicting Adult Symptoms of Posttraumatic Stress Disorder. JAMA Psychiatry 2014, 71, 1174–1182. [Google Scholar]
- Bountress, K.E.; Bacanu, S.-A.; Tomko, R.L.; Korte, K.J.; Hicks, T.; Sheerin, C.; Lind, M.J.; Marraccini, M.; Nugent, N.; Amstadter, A.B. The Effects of a BDNF Val66Met Polymorphism on Posttraumatic Stress Disorder: A Meta-Analysis. Neuropsychobiology 2017, 76, 136–142. [Google Scholar]
- Svob Strac, D.; Kovacic Petrovic, Z.; Nikolac Perkovic, M.; Umolac, D.; Nedic Erjavec, G.; Pivac, N. Platelet monoamine oxidase type B, MAOB intron 13 and MAOA-uVNTR polymorphism and symptoms of post-traumatic stress disorder. Stress 2016, 19, 362–373. [Google Scholar]
- Schmeltzer, S.N.; Herman, J.P.; Sah, R. Neuropeptide Y (NPY) and posttraumatic stress disorder (PTSD): A translational update. Exp. Neurol. 2016, 284 (Pt. B), 196–210. [Google Scholar]
- Averill, L.A.; Abdallah, C.G.; Levey, D.F.; Han, S.; Harpaz-Rotem, I.; Kranzler, H.R.; Southwick, S.M.; Krystal, J.H.; Gelernter, J.; Pietrzak, R.H. Apolipoprotein E gene polymorphism, posttraumatic stress disorder, and cognitive function in older U.S. veterans: Results from the National Health and Resilience in Veterans Study. Depress. Anxiety 2019, 36, 834–845. [Google Scholar]
- Johnson, L.A.; Zuloaga, D.G.; Bidiman, E.; Marzulla, T.; Weber, S.; Wahbeh, H.; Raber, J. ApoE2 Exaggerates PTSD-Related Behavioral, Cognitive, and Neuroendocrine Alterations. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2015, 40, 2443–2453. [Google Scholar]
- Sippel, L.M.; Han, S.; Watkins, L.E.; Harpaz-Rotem, I.; Southwick, S.M.; Krystal, J.H.; Olff, M.; Sherva, R.; Farrer, L.A.; Kranzler, H.R.; et al. Oxytocin receptor gene polymorphisms, attachment, and PTSD: Results from the National Health and Resilience in Veterans Study. J. Psychiatr. Res. 2017, 94, 139–147. [Google Scholar]
- Crombie, K.M.; Privratsky, A.A.; Schomaker, C.M.; Heilicher, M.; Ross, M.C.; Sartin-Tarm, A.; Sellnow, K.; Binder, E.B.; Andrew James, G.; Cisler, J.M. The influence of FAAH genetic variation on physiological, cognitive, and neural signatures of fear acquisition and extinction learning in women with PTSD. NeuroImage Clin. 2022, 33, 102922. [Google Scholar]
- Sullivan, D.R.; Morrison, F.G.; Wolf, E.J.; Logue, M.W.; Fortier, C.B.; Salat, D.H.; Fonda, J.R.; Stone, A.; Schichman, S.; Milberg, W.; et al. The PPM1F gene moderates the association between PTSD and cortical thickness. J. Affect. Disord. 2019, 259, 201–209. [Google Scholar]
- Solovieff, N.; Roberts, A.L.; Ratanatharathorn, A.; Haloosim, M.; De Vivo, I.; King, A.P.; Liberzon, I.; Aiello, A.; Uddin, M.; Wildman, D.E.; et al. Genetic association analysis of 300 genes identifies a risk haplotype in SLC18A2 for post-traumatic stress disorder in two independent samples. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2014, 39, 1872–1879. [Google Scholar]
- Andero, R.; Brothers, S.P.; Jovanovic, T.; Chen, Y.T.; Salah-Uddin, H.; Cameron, M.; Bannister, T.D.; Almli, L.; Stevens, J.S.; Bradley, B.; et al. Amygdala-dependent fear is regulated by Oprl1 in mice and humans with PTSD. Sci. Transl. Med. 2013, 5, 188ra73. [Google Scholar]
- Liu, Y.; Rimmler, J.; Dennis, M.F.; Ashley-Koch, A.E.; Hauser, M.A.; Mid-Atlantic Mental Illness Research Education Clinical Center Workgroup; Beckham, J.C. Association of Variant rs4790904 in Protein Kinase C Alpha with Posttraumatic Stress Disorder in a U.S. Caucasian and African-American Veteran Sample. J. Depress. Anxiety 2013, 2, S4001. [Google Scholar]
- Flaquer, A.; Baumbach, C.; Ladwig, K.-H.; Kriebel, J.; Waldenberger, M.; Grallert, H.; Baumert, J.; Meitinger, T.; Kruse, J.; Peters, A.; et al. Mitochondrial genetic variants identified to be associated with posttraumatic stress disorder. Transl. Psychiatry 2015, 5, e524. [Google Scholar]
- Guffanti, G.; Galea, S.; Yan, L.; Roberts, A.L.; Solovieff, N.; Aiello, A.E.; Smoller, J.W.; De Vivo, I.; Ranu, H.; Uddin, M.; et al. Genome-wide association study implicates a novel RNA gene, the lincRNA AC068718.1, as a risk factor for post-traumatic stress disorder in women. Psychoneuroendocrinology 2013, 38, 3029–3038. [Google Scholar]
- Logue, M.W.; Baldwin, C.; Guffanti, G.; Melista, E.; Wolf, E.J.; Reardon, A.F.; Uddin, M.; Wildman, D.; Galea, S.; Koenen, K.C.; et al. A genome-wide association study of post-traumatic stress disorder identifies the retinoid-related orphan receptor alpha (RORA) gene as a significant risk locus. Mol. Psychiatry 2013, 18, 937–942. [Google Scholar]
- Nievergelt, C.M.; Maihofer, A.X.; Mustapic, M.; Yurgil, K.A.; Schork, N.J.; Miller, M.W.; Logue, M.W.; Geyer, M.A.; Risbrough, V.B.; O’Connor, D.T.; et al. Genomic predictors of combat stress vulnerability and resilience in U.S. Marines: A genome-wide association study across multiple ancestries implicates PRTFDC1 as a potential PTSD gene. Psychoneuroendocrinology 2015, 51, 459–471. [Google Scholar]
- Xie, P.; Kranzler, H.R.; Yang, C.; Zhao, H.; Farrer, L.A.; Gelernter, J. Genome-wide Association Study Identifies New Susceptibility Loci for Posttraumatic Stress Disorder. Biomark. Posttraumatic Stress Disord. 2013, 74, 656–663. [Google Scholar]
- Stein, M.B.; Chen, C.-Y.; Ursano, R.J.; Cai, T.; Gelernter, J.; Heeringa, S.G.; Jain, S.; Jensen, K.P.; Maihofer, A.X.; Mitchell, C.; et al. Genome-wide Association Studies of Posttraumatic Stress Disorder in 2 Cohorts of US Army Soldiers. JAMA Psychiatry 2016, 73, 695–704. [Google Scholar]
- Morey, R.A.; Gold, A.L.; LaBar, K.S.; Beall, S.K.; Brown, V.M.; Haswell, C.C.; Nasser, J.D.; Wagner, H.R.; McCarthy, G.; for the Mid-Atlantic MIRECC Workgroup. Amygdala Volume Changes in Posttraumatic Stress Disorder in a Large Case-Controlled Veterans Group. Arch. Gen. Psychiatry 2012, 69, 1169–1178. [Google Scholar]
- Li, L.; Bao, Y.; He, S.; Wang, G.; Guan, Y.; Ma, D.; Wang, P.; Huang, X.; Tao, S.; Zhang, D.; et al. The Association Between Genetic Variants in the Dopaminergic System and Posttraumatic Stress Disorder: A Meta-Analysis. Medicine 2016, 95, e3074. [Google Scholar]
- Wolf, E.J.; Mitchell, K.S.; Logue, M.W.; Baldwin, C.T.; Reardon, A.F.; Aiello, A.; Galea, S.; Koenen, K.C.; Uddin, M.; Wildman, D.; et al. The dopamine D3 receptor gene and posttraumatic stress disorder. J. Trauma. Stress 2014, 27, 379–387. [Google Scholar]
- Pitts, B.L.; Whealin, J.M.; Harpaz-Rotem, I.; Duman, R.S.; Krystal, J.H.; Southwick, S.M.; Pietrzak, R.H. BDNF Val66Met polymorphism and posttraumatic stress symptoms in U.S. military veterans: Protective effect of physical exercise. Psychoneuroendocrinology 2019, 100, 198–202. [Google Scholar]
- Qi, R.; Cao, Z.; Surento, W.; Zhang, L.; Qiu, L.; Xia, Z.; Ching, C.R.K.; Xu, Q.; Yin, Y.; Zhang, L.J.; et al. RORA rs8042149 polymorphism moderates the association between PTSD symptom severity and transverse temporal gyrus thickness in Han Chinese adults who lost their only child. J. Affect. Disord. 2022, 314, 318–324. [Google Scholar]
- Ferrucci, M.; Limanaqi, F.; Ryskalin, L.; Biagioni, F.; Busceti, C.L.; Fornai, F. The Effects of Amphetamine and Methamphetamine on the Release of Norepinephrine, Dopamine and Acetylcholine From the Brainstem Reticular Formation. Front. Neuroanat. 2019, 13, 48. [Google Scholar]
- Pitts, B.L.; Eisenberg, M.L.; Bailey, H.R.; Zacks, J.M. PTSD is associated with impaired event processing and memory for everyday events. Cogn. Res. Princ. Implic. 2022, 7, 35. [Google Scholar]
- Bienvenu TC, M.; Dejean, C.; Jercog, D.; Aouizerate, B.; Lemoine, M.; Herry, C. The advent of fear conditioning as an animal model of post-traumatic stress disorder: Learning from the past to shape the future of PTSD research. Neuron 2021, 109, 2380–2397. [Google Scholar]
- Stevens, J.S.; Jovanovic, T. Role of social cognition in post-traumatic stress disorder: A review and meta-analysis. Genes Brain Behav. 2019, 18, e12518. [Google Scholar]
- Paizanis, E.; Crotti, M.; Petit, A.; Règue, M.; Beray-Berthat, V.; Noble, F.; Lanfumey, L.; Mongeau, R. Effects of Alcohol and Cocaine in a Mutant Mouse Model of Predisposition to Post-Traumatic Stress Disorder. Front. Pharmacol. 2020, 11, 623. [Google Scholar]
- Skórzewska, A.; Lehner, M.; Wisłowska-Stanek, A.; Turzyńska, D.; Sobolewska, A.; Krząścik, P.; Szyndler, J.; Maciejak, P.; Chmielewska, N.; Kołosowska, K.; et al. Individual susceptibility or resistance to posttraumatic stress disorder-like behaviours. Behav. Brain Res. 2020, 386, 112591. [Google Scholar]
- Krystal, J.H.; Abdallah, C.G.; Averill, L.A.; Kelmendi, B.; Harpaz-Rotem, I.; Sanacora, G.; Southwick, S.M.; Duman, R.S. Synaptic Loss and the Pathophysiology of PTSD: Implications for Ketamine as a Prototype Novel Therapeutic. Curr. Psychiatry Rep. 2017, 19, 74. [Google Scholar]
- Reichmann, F.; Holzer, P. Neuropeptide Y: A stressful review. Neuropeptides 2016, 55, 99–109. [Google Scholar]
- Branco, R.C.; Burkett, J.P.; Black, C.A.; Winokur, E.; Ellsworth, W.; Dhamsania, R.K.; Lohr, K.M.; Schroeder, J.P.; Weinshenker, D.; Jovanovic, T.; et al. Vesicular monoamine transporter 2 mediates fear behavior in mice. Genes Brain Behav. 2020, 19, e12634. [Google Scholar]



| Description | Relative Enrichment | Genes |
|---|---|---|
| response to amphetamine | 82.7 | OXTR, DBH, DRD4, SLC18A2, RGS2 |
| dopamine catabolic process | 211.8 | SLC6A3, MAOB, DBH, COMT |
| memory | 36.5 | OXTR, BDNF, DBH, CNR1, HTR2A, SLC6A4 |
| negative regulation of voltage-gated calcium channel activity | 141.2 | CRHR1, OPRL1, DRD3, DRD4 |
| G protein-coupled receptor signaling pathway | 6.7 | ADRB2, CRHR1, OXTR, OPRL1, APOE, DRD3, CNR1, ADCYAP1, HTR2A, CRHR2, DRD4, RGS2 |
| fear response | 226.9 | CRHR1, DBH, DRD4 |
| negative regulation of blood pressure | 75.6 | NPY, OPRL1, DRD3, CNR1 |
| positive regulation of vasoconstriction | 66.2 | FAAH, OXTR, DBH, HTR2A |
| response to cocaine | 62.3 | SLC6A3, OXTR, DRD3, CNR1 |
| monoamine transport | 176.5 | SLC6A3, SLC6A4, SLC18A2 |
| behavioral response to ethanol | 158.9 | CRHR1, DBH, DRD4 |
| response to ethanol | 25.7 | SLC6A3, MAOB, DRD3, CNR1, RGS2 |
| behavioral response to cocaine | 113.5 | DRD3, HTR2A, DRD4 |
| social behavior | 40.7 | OXTR, DRD3, DRD4, SLC6A4 |
| dopamine metabolic process | 99.3 | DRD3, DRD4, COMT |
| Description | Relative Enrichment | Genes |
|---|---|---|
| neuron projection | 11.9 | SLC6A3, TPH2, CRHR1, NPY, OPRL1, GABRA2, ADCYAP1, SLC6A4, RGS2 |
| integral component of plasma membrane | 5.2 | ADRB2, SLC6A3, CRHR1, OXTR, OPRL1, GABRA2, DRD3, CNR1, HTR2A, CRHR2, DRD4, SLC6A4, SLC18A2 |
| integral component of presynaptic membrane | 41.7 | SLC6A3, CNR1, HTR2A, SLC6A4 |
| dendrite | 8.5 | BDNF, GABRA2, APOE, HTR2A, CRHR2, DRD4, COMT |
| axon | 8.5 | SLC6A3, BDNF, GABRA2, CNR1, HTR2A, COMT |
| integral component of postsynaptic membrane | 32.4 | SLC6A3, HTR2A, SLC6A4 |
| dopaminergic synapse | 104.1 | SLC6A3, SLC18A2 |
| glutamatergic synapse | 8.9 | APOE, DRD3, CNR1, HTR2A, DRD4 |
| GABA-ergic synapse | 24.9 | GABRA2, DRD3, CNR1 |
| plasma membrane | 1.9 | ADRB2, PRKCA, SLC6A3, CRHR1, OXTR, OPRL1, GABRA2, APOE, DRD3, CNR1, HTR2A, CRHR2, DRD4, SLC6A4, SLC18A2, RGS2, COMT |
| synapse | 4.2 | DBH, GABRA2, DRD3, CNR1, HTR2A, SLC6A4 |
| presynapse | 10.9 | CNR1, HTR2A, SLC6A4 |
| discoidal high-density lipoprotein particle | 286.1 | APOE |
| serotonergic synapse | 286.1 | SLC6A4 |
| cytoplasmic vesicle | 4.1 | NPY, OPRL1, DBH, GABRA2, HTR2A, SLC18A2 |
| Description | Relative Enrichment | Genes |
|---|---|---|
| monoamine transmembrane transporter activity | 172.9 | SLC6A3, SLC6A4, SLC18A2 |
| G protein-coupled receptor activity | 7.1 | ADRB2, CRHR1, OXTR, NPY, OPRL1, DRD3, CNR1, HTR2A, CRHR2, DRD4 |
| corticotropin-releasing hormone receptor activity | 345.9 | CRHR1, CRHR2 |
| dopamine neurotransmitter receptor activity, coupled via Gi/Go | 345.9 | DRD3, DRD4 |
| corticotrophin-releasing factor receptor activity | 345.9 | CRHR1, CRHR2 |
| serotonin:sodium symporter activity | 345.9 | SLC6A4, SLC18A2 |
| norepinephrine binding | 259.5 | ADRB2, DRD4 |
| dopamine neurotransmitter receptor activity | 207.6 | DRD3, DRD4 |
| peptide hormone binding | 36.2 | CRHR1, OXTR, CRHR2 |
| dopamine binding | 148.3 | SLC6A3, DRD4 |
| serotonin binding | 86.5 | HTR2A, SLC6A4 |
| neurotransmitter transmembrane transporter activity | 57.7 | SLC6A3, SLC6A4 |
| neurotransmitter receptor activity | 18.8 | GABRA2, HTR2A, DRD4 |
| signaling receptor binding | 6.1 | SLC6A3, BDNF, NPY, APOE, ADCYAP1 |
| G protein-coupled serotonin receptor activity | 30.6 | HTR2A, DRD4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skolariki, K.; Vrahatis, A.G.; Krokidis, M.G.; Exarchos, T.P.; Vlamos, P. Assessing and Modelling of Post-Traumatic Stress Disorder Using Molecular and Functional Biomarkers. Biology 2023, 12, 1050. https://doi.org/10.3390/biology12081050
Skolariki K, Vrahatis AG, Krokidis MG, Exarchos TP, Vlamos P. Assessing and Modelling of Post-Traumatic Stress Disorder Using Molecular and Functional Biomarkers. Biology. 2023; 12(8):1050. https://doi.org/10.3390/biology12081050
Chicago/Turabian StyleSkolariki, Konstantina, Aristidis G. Vrahatis, Marios G. Krokidis, Themis P. Exarchos, and Panagiotis Vlamos. 2023. "Assessing and Modelling of Post-Traumatic Stress Disorder Using Molecular and Functional Biomarkers" Biology 12, no. 8: 1050. https://doi.org/10.3390/biology12081050