
Specific Neural Mechanisms of Self-Cognition and the Application of Brainprint Recognition

Abstract
:Simple Summary
Abstract
1. Introduction
2. Behavioral Uniqueness of Self-Awareness
2.1. Priority of Self-Awareness
2.2. Sensitivity of Self-Information
2.3. Memory Advantage of Self-Reference
3. fMRI and PET Imaging of Self-Information Processing
4. EEG Study of Self-Information Processing
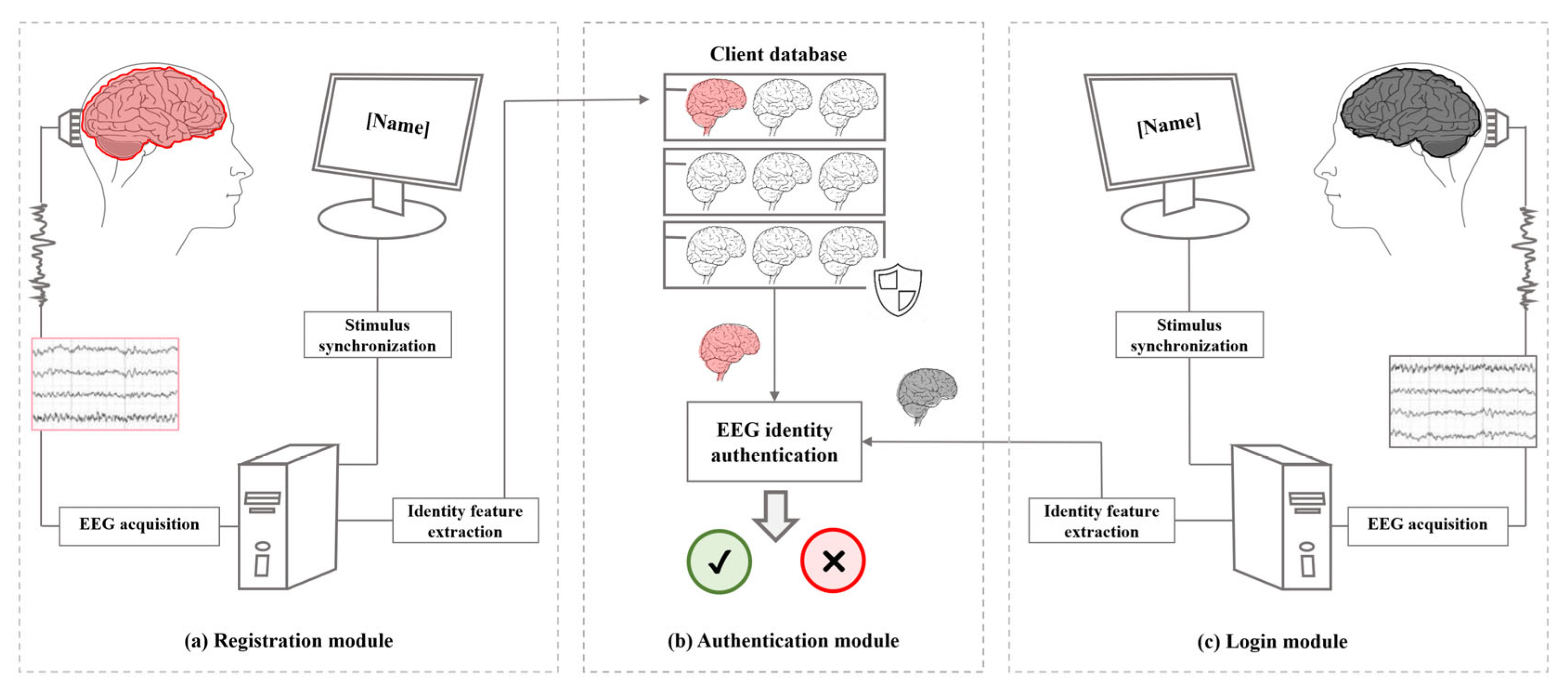
5. Application of Self-Specificity in Brainprint Recognition
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Desebrock, C. Self-Prioritization in Motor Responses. Ph.D. Thesis, University of Oxford, Oxford, UK, 2022. [Google Scholar]
- Maire, H.; Brochard, R.; Zagar, D. A developmental study of the self-prioritization effect in children between 6 and 10 years of age. Child Dev. 2020, 91, 694–704. [Google Scholar] [CrossRef]
- Honda, T.; Nakao, T. Interoception modulates the self-prioritization effect. PsyArXiv 2021. [Google Scholar]
- Veldhuis, A. The Influence of Self-Relevant Information on Attention and Memory. Ph.D. Thesis, Oxford Brookes University, Oxford, UK, 2019. [Google Scholar]
- Gallup, G.G., Jr. Chimpanzees: Self-Recognition. Science 1970, 167, 86–87. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.S. The named and the nameless: Gender and person in Chinese society. Am. Ethnol. 2010, 13, 619–631. [Google Scholar] [CrossRef]
- Desebrock, C.; Spence, C. The self-prioritization effect: Self-referential processing in movement highlights modulation at multiple stages. Atten. Percept. Psychophys. 2021, 83, 2656–2674. [Google Scholar] [CrossRef]
- Ota, C.; Nakano, T. Self-face activates the dopamine reward pathway without awareness. Cereb. Cortex 2021, 31, 4420–4426. [Google Scholar] [CrossRef] [PubMed]
- Orellana-Corrales, G.; Matschke, C.; Schäfer, S.; Wesslein, A.-K. Does an experimentally induced self-association elicit affective self-prioritisation? Q. J. Exp. Psychol. 2022, 17470218221124928. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, S.; Wesslein, A.-K.; Spence, C.; Frings, C. When self-prioritization crosses the senses: Crossmodal self-prioritization demonstrated between vision and touch. Br. J. Psychol. 2021, 112, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Nakayama, K. Robust representations for faces: Evidence from visual search. J. Exp. Psychol. Hum. Percept. Perform. 1999, 25, 1016–1035. [Google Scholar] [CrossRef]
- Moray, N. Attention in dichotic listening: Affective cues and the influence of instructions. Q. J. Exp. Psychol. 1959, 11, 56–60. [Google Scholar] [CrossRef]
- Greenwald, A.G.; Banaji, M.R. The self as a memory system: Powerful, but ordinary. J. Personal. Soc. Psychol. 1989, 57, 41. [Google Scholar] [CrossRef]
- Niu, G.; Yao, L.; Kong, F.; Luo, Y.; Duan, C.; Sun, X.; Zhou, Z. Behavioural and ERP evidence of the self-advantage of online self-relevant information. Sci. Rep. 2020, 10, 20515. [Google Scholar] [CrossRef] [PubMed]
- Deng, N.; Sun, Y.; Chen, X.; Li, W. How does self name influence the neural processing of emotional prosody? An ERP Study. PsyCh J. 2022, 11, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, F.; Rubianes, M.; Jiménez-Ortega, L.; Fondevila, S.; Hernández-Gutiérrez, D.; Sánchez-García, J.; Martínez-de-Quel, Ó.; Casado, P.; Martín-Loeches, M. Spatio-temporal brain dynamics of self-identity: An EEG source analysis of the current and past self. Brain Struct. Funct. 2022, 227, 2167–2179. [Google Scholar] [CrossRef]
- Alatorre-Cruz, G.C.; Downs, H.; Hagood, D.; Sorensen, S.T.; Williams, D.K.; Larson-Prior, L. Effect of obesity on inhibitory control in preadolescents during stop-signal task. An event-related potentials study. Int. J. Psychophysiol. 2021, 165, 56–67. [Google Scholar] [CrossRef]
- Kelleyw, M.; Macraec, N.; Wylandc, L.; Caglar, S.; Inati, S.; Heathertont, F. Finding the self? An event-related FMRI study. J. Cogn. Neurosci. 2002, 14, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Koch, C. What is consciousness? Nature 2018, 557, S8–S12. [Google Scholar] [CrossRef] [Green Version]
- Leszkowicz, E.; Maio, G.R.; Linden, D.E.; Ihssen, N. Neural coding of human values is underpinned by brain areas representing the core self in the cortical midline region. Soc. Neurosci. 2021, 16, 486–499. [Google Scholar] [CrossRef]
- Martínez-Pérez, V.; Campoy, G.; Palmero, L.B.; Fuentes, L.J. Examining the dorsolateral and ventromedial prefrontal cortex involvement in the self-attention network: A randomized, sham-controlled, parallel group, double-blind, and multichannel HD-TDCS study. Front. Neurosci. 2020, 14, 683. [Google Scholar] [CrossRef]
- Perrin, F.; Maquet, P.; Peigneux, P.; Ruby, P.; Degueldre, C.; Balteau, E.; Fiore, G.D.; Moonen, G.; Luxen, A.; Laureys, S. Neural mechanisms involved in the detection of our first name: A combined ERPs and PET study. Neuropsychologia 2005, 43, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Jublie, A.; Kumar, D. Early capture of attention by self-face: Investigation using a temporal order judgment task. I-Perception 2021, 12, 20416695211032990. [Google Scholar] [CrossRef]
- Smith, D.; Wolff, A.; Wolman, A.; Ignaszewski, J.; Northoff, G. Temporal continuity of self: Long autocorrelation windows mediate self-specificity. NeuroImage 2022, 257, 119305. [Google Scholar] [CrossRef] [PubMed]
- Caughey, S.; Falbén, J.K.; Tsamadi, D.; Persson, L.M.; Golubickis, M.; Neil Macrae, C. Self-prioritization during stimulus processing is not obligatory. Psychol. Res. 2021, 85, 503–508. [Google Scholar] [CrossRef] [Green Version]
- McCrackin, S.D.; Lee, C.M.; Itier, R.J.; Fernandes, M.A. Meaningful faces: Self-relevance of semantic context in an initial social encounter improves later face recognition. Psychon. Bull. Rev. 2021, 28, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Rotshtein, P. Self-prioritization and the attentional systems. Curr. Opin. Psychol. 2019, 29, 148–152. [Google Scholar] [CrossRef]
- Schäfer, S.; Frings, C. Understanding self-prioritisation: The prioritisation of self-relevant stimuli and its relation to the individual self-esteem. J. Cogn. Psychol. 2019, 31, 813–824. [Google Scholar] [CrossRef]
- Golubickis, M.; Falbén, J.K.; Ho, N.S.; Sui, J.; Cunningham, W.A.; Macrae, C.N. Parts of me: Identity-relevance moderates self-prioritization. Conscious. Cogn. 2020, 77, 102848. [Google Scholar] [CrossRef]
- Żochowska, A.; Jakuszyk, P.; Nowicka, M.M.; Nowicka, A. the self and a close-other: Differences between processing of faces and newly acquired information. Cereb. Cortex 2023, 33, 2183–2199. [Google Scholar] [CrossRef]
- Falbén, J.K.; Golubickis, M.; Tamulaitis, S.; Caughey, S.; Tsamadi, D.; Persson, L.M.; Svensson, S.L.; Sahraie, A.; Macrae, C.N. Self-relevance enhances evidence gathering during decision-making. Acta Psychol. 2020, 209, 103122. [Google Scholar] [CrossRef]
- Liu, S.; Jia, Y.; Zheng, S.; Feng, S.; Zhu, H.; Wang, R.; Jia, H. An experimental study of subliminal self-face processing in depersonalization–derealization disorder. Brain Sci. 2022, 12, 1598. [Google Scholar] [CrossRef]
- Svensson, S.L.; Golubickis, M.; Maclean, H.; Falbén, J.K.; Persson, L.M.; Tsamadi, D.; Caughey, S.; Sahraie, A.; Macrae, C.N. More or less of me and you: Self-relevance augments the effects of item probability on stimulus prioritization. Psychol. Res. 2022, 86, 1145–1164. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, S.; Wentura, D.; Frings, C. Creating a network of importance: The particular effects of self-relevance on stimulus processing. Atten. Percept. Psychophys. 2020, 82, 3750–3766. [Google Scholar] [CrossRef]
- Keenan, J.P.; Gorman, J. The causal role of the right hemisphere in self-awareness: It is the brain that is selective. Cortex 2007, 43, 1074–1082. [Google Scholar] [CrossRef]
- Sui, J.; Zhu, Y.; Han, S. Self-face recognition in attended and unattended conditions: An event-related brain potential study. Neuroreport 2006, 17, 423–427. [Google Scholar] [CrossRef]
- Gray, H.M.; Ambady, N.; Lowenthal, W.T.; Deldin, P. P300 as an index of attention to self-relevant stimuli. J. Exp. Soc. Psychol. 2004, 40, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Han, S. Why we respond faster to the self than to others? An implicit positive association theory of self-advantage during implicit face recognition. J. Exp. Psychol. Hum. Percept. Perform. 2010, 36, 619–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.R.; Pashler, H.E.; Coburn, P. Moray revisited: High-priority affective stimuli and visual search. Q. J. Exp. Psychol. Sect. A 2004, 57, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Tamaki, M.; Shibata, K.; Worden, M.S.; Yamada, T.; Kawato, M.; Sasaki, Y.; Watanabe, T. Visual perceptual learning of a primitive feature in human V1/V2 as a result of unconscious processing, revealed by decoded FMRI neurofeedback (DecNef). J. Vis. 2020, 21, 24. [Google Scholar] [CrossRef]
- Tacikowski, P.; Brechmann, A.; Marchewka, A.; Jednoróg, K.; Dobrowolny, M.; Nowicka, A. Is it about the self or the significance? An FMRI study of self-name recognition. Soc. Neurosci. 2011, 6, 98–107. [Google Scholar] [CrossRef]
- Keenan, J.P.; Nelson, A.; O’connor, M.; Pascual-Leone, A. Alvaro self-recognition and the right hemisphere. Nature 2001, 409, 305. [Google Scholar] [CrossRef]
- Yamada, Y.; Kawabe, T.; Miura, K. One’s own name distorts visual space. Neurosci. Lett. 2012, 531, 96–98. [Google Scholar] [CrossRef]
- Wang, L.; Li, Z.; Wang, L.; Tong, L.; Zeng, Y.; Yan, B. Edge-assistant visual objects decoding using sparse representation. In Proceedings of the 7th International Conference on Biomedical Engineering and Informatics, Dalian, China, 14–16 October 2014; pp. 654–658. [Google Scholar]
- Alexopoulos, T.; Muller, D.; Ric, F.; Marendaz, C.I. Me, Mine: Automatic attentional capture by self-related stimuli. Eur. J. Soc. Psychol. 2012, 42, 770–779. [Google Scholar] [CrossRef]
- Callan, D.E.; Tsytsarev, V.; Hanakawa, T.; Callan, A.M.; Katsuhara, M.; Fukuyama, H.; Turner, R. Song and speech: Brain regions involved with perception and covert production. Neuroimage 2006, 31, 1327–1342. [Google Scholar] [CrossRef] [PubMed]
- Pfister, R.; Pohl, C.; Kiesel, A.; Kunde, W. Your unconscious knows your name. PLoS ONE 2012, 7, e32402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, K.L.; Caldwell, J.; Sorensen, R.E. Personal names and the attentional blink: A visual “cocktail party” effect. J. Exp. Psychol. Hum. Percept. Perform. 1997, 23, 504. [Google Scholar] [CrossRef]
- Pannese, A.; Hirsch, J. Self-specific priming effect. Conscious. Cogn. 2010, 19, 962–968. [Google Scholar] [CrossRef]
- Xiao, L.; Lei, G.; Qiang-Lin, Z. The relative uniqueness of personal names: Evidences from the RSVP. J. Psychol. Sci. 2013, 36, 72. [Google Scholar]
- Brédart, S.; Delchambre, M.; Laureys, S. Short article: One’s own face is hard to ignore. Q. J. Exp. Psychol. 2006, 59, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Devue, C.; Brédart, S. Attention to self-referential stimuli: Can I ignore my own face? Acta Psychol. 2008, 128, 290–297. [Google Scholar] [CrossRef]
- Devue, C.; Stigchel, S.; Brédart, S.; Theeuwes, J. You do not find your own face faster; you just look at it longer. Cognition 2009, 111, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Wolford, G.; Morrison, F. Processing of unattended visual information. Mem. Cogn. 1980, 8, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Perrin, F.; Bastuji, H.; Mauguière, F.; García-Larrea, L. Functional dissociation of the early and late portions of human K-complexes. Neuroreport 2000, 11, 1637. [Google Scholar] [CrossRef]
- Oswald, I.; Taylor, A.M.; Treisman, M. Discriminative responses to stimulation during human sleep. Brain A J. Neurol. 1960. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.M.; Coleman, M.R.; Boly, M.; Davis, M.H.; Laureys, S.; Pickard, J.D. Detecting awareness in the vegetative state. Science 2006, 313, 1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, D.; Trapp, C.; Kieny, M.; Wassmer, J.; Mugnaioni, M.; Pack, A.; Hoff, E. Study of recovery and the post-anaesthetic period (Author’s Translation). Rev. D’electroencephalogr. Neurophysiol. Clin. 1977, 7, 62–69. [Google Scholar] [CrossRef]
- Fishback, D.B. Mental status questionnaire for organic brain syndrome, with a new visual counting test. J. Am. Geriatr. Soc. 1977, 25, 167. [Google Scholar] [CrossRef]
- Tsytsarev, V. Methodological aspects of studying the mechanisms of consciousness. Behav. Brain Res. 2022, 419, 113684. [Google Scholar] [CrossRef]
- Kliemann, D.; Jacoby, N.; Anzellotti, S.; Saxe, R.R. decoding task and stimulus representations in face-responsive cortex. Cogn. Neuropsychol. 2016, 33, 362–377. [Google Scholar] [CrossRef] [Green Version]
- Bower, G.H.; Gilligan, S.G. Remembering information related to one’s self. J. Res. Personal. 1979, 13, 420–432. [Google Scholar] [CrossRef]
- Dewhurst, S.A.; Anderson, R.J.; Grace, L.; Boland, J. Survival processing versus self-reference: A memory advantage following descriptive self-referential encoding. J. Mem. Lang. 2017, 94, 291–304. [Google Scholar] [CrossRef]
- Rogers, T.B.; Kuiper, N.A.; Kirker, W.S. Self-reference and the encoding of personal information. J. Personal. Soc. Psychol. 1977, 35, 677. [Google Scholar] [CrossRef]
- Symons, C.S.; Johnson, B.T. The self-reference effect in memory: A meta-analysis. Psychol. Bull. 1997, 121, 371–394. [Google Scholar] [CrossRef]
- D’Argembeau, A.; Comblain, C. Affective valence and the self-reference effect: Influence of retrieval conditions. Br. J. Psychol. 2011, 96, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Tacikowski, P.; Brechmann, A.; Nowicka, A. Cross-modal pattern of brain activations associated with the processing of self- and significant other’s name. Hum. Brain Mapp. 2013, 34, 2069–2077. [Google Scholar] [CrossRef]
- Johnson, S.C.; Baxter, L.C.; Wilder, L.S.; Pipe, J.G.; Heiserman, J.E.; Prigatano, G.P. Neural correlates of self-reflection. Brain 2002, 125, 1808–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninomiya, H.; Onitsuka, T.; Chen, C.H.; Sato, E.; Tashiro, N. P300 in response to the subject’s own face. Psychiatry Clin. Neurosci. 1998, 52, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Kawashima, R.; Nakamura, K.; Okada, K.; Kato, T.; Nakamura, A.; Hatano, K.; Itoh, K.; Kojima, S.; Fukuda, H. Passive and active recognition of one’s own face. NeuroImage 2000, 11, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Gusnard, D.A.; Akbudak, E.; Shulman, G.L.; Raichle, M.E. Medial prefrontal cortex and self-referential mental activity: Relation to a default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 4259–4264. [Google Scholar] [CrossRef] [Green Version]
- Shulman, G.L.; Corbetta, M.; Buckner, R.L.; Fiez, J.A.; Miezin, F.M.; Raichle, M.E.; Petersen, S.E. C Common blood flow changes across visual tasks: I. Increases in subcortical structures and cerebellum but not in nonvisual cortex. J. Cogn. Neurosci. 1997, 9, 624–647. [Google Scholar] [CrossRef] [Green Version]
- Craik, F.; Moroz, T.M.; Moscovitch, M.; Stuss, D.T.; Winocur, G.; Tulving, E.; Kapur, S. In search of the self: A positron emission tomography study. Psychol. Sci. 2010, 10, 26–34. [Google Scholar] [CrossRef]
- Ackerly, S.S.; Benton, A.L. Report of case of bilateral frontal lobe defect. Res. Publ.-Assoc. Res. Nerv. Ment. Dis. 1948, 27, 479–504. [Google Scholar]
- Platek, S.M.; Keenan, J.P.; Gallup, G.G.; Mohamed, F.B. Where am I? The neurological correlates of self and other. Cogn. Brain Res. 2004, 19, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.T.; Aziz-Zadeh, L.; Uddin, L.Q.; Iacoboni, M. The self across the senses: An FMRI study of self-face and self-voice recognition. Soc. Cogn. Affect. Neurosci. 2008, 3, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northoff, G.; Bermpohl, F. Cortical midline structures and the self. Trends Cogn. Sci. 2004, 8, 102–107. [Google Scholar] [CrossRef] [PubMed]
- RongKai, Z.; Yan, B.; Ying, Z.; Li, T.; Jun, S.; GuangCheng, B. EEG-based identity authentication using subliminal name stimuli. Int. J. Psychophysiol. 2021, 168, S193. [Google Scholar] [CrossRef]
- Caharel, S.; Courtay, N.; Bernard, C.; Lalonde, R.; Rebaie, M. Familiarity and emotional expression influence an early stage of face processing: An electrophysiological study. Brain Cogn. 2005, 59, 96–100. [Google Scholar] [CrossRef]
- Caharel, S.; Poiroux, S.; Bernard, C.; Thibaut, F.; Lalonde, R.; Rebai, M. ERPs associated with familiarity and degree of familiarity during face recognition. Int. J. Neurosci. 2002, 112, 1499–1512. [Google Scholar] [CrossRef]
- Keyes, H.; Brady, N.; Reilly, R.B.; Foxe, J.J. My face or yours? Event-related potential correlates of self-face processing. Brain Cogn. 2010, 72, 244–254. [Google Scholar] [CrossRef]
- Rossion, B.; Jacques, C. Does physical interstimulus variance account for early electrophysiological face sensitive responses in the human brain? Ten Lessons on the N170. NeuroImage 2008, 39, 1959–1979. [Google Scholar] [CrossRef]
- Müller, H.M.; Kutas, M. What’s in a name? Electrophysiological differences between spoken nouns, proper names and one’s own name. Neuroreport 1996, 8, 221–225. [Google Scholar] [CrossRef]
- Eimer, M. Event-related brain potentials distinguish processing stages involved in face perception and recognition. Clin. Neurophysiol. 2000, 111, 694–705. [Google Scholar] [CrossRef]
- Bentin, S.; Deouell, L.Y. Structural encoding and identification in face processing: ERP evidence for separate mechanisms. Cogn. Neuropsychol. 2000, 17, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Perrin, F.; García-Larrea, L.; Mauguière, F.; Bastuji, H. A differential brain response to the subject’s own name persists during sleep. Clin. Neurophysiol. 1999, 110, 2153–2164. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, J.W.; Curran, T.; Porterfield, A.L.; Collins, D. Activation of preexisting and acquired face representations: The N250 event-related potential as an index of face familiarity. J. Cogn. Neurosci. 2006, 18, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Folmer, R.L.; Yingling, C.D. Auditory P3 responses to name stimuli. Brain Lang. 1997, 56, 306–311. [Google Scholar] [CrossRef]
- Webb, S.J.; Dawson, G.; Bernier, R.; Panagiotides, H. erp evidence of atypical face processing in young children with autism. J. Autism Dev. Disord. 2006, 36, 881–890. [Google Scholar] [CrossRef]
- Signorino, M.; D’Acunto, S.; Angeleri, F.; Pietropaoli, P. Eliciting P300 in comatose patients. Lancet 1995, 345, 255–256. [Google Scholar] [CrossRef]
- Fischer, C.; Dailler, F.; Morlet, D. Novelty P3 elicited by the subject’s own name in comatose patients. Clin. Neurophysiol. Shannon 2008, 119, 2224–2230. [Google Scholar] [CrossRef]
- Mu, Y.; Han, S. neural oscillations involved in self-referential processing. Neuroimage 2010, 53, 757–768. [Google Scholar] [CrossRef]
- Wang, M.; Hu, J.; Abbass, H.A. BrainPrint: EEG biometric identification based on analyzing brain connectivity graphs. Pattern Recognit. 2020, 105, 107381. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Behavior | Author | Experiment | Control Group | Conclusion |
|---|---|---|---|---|
| Keys | Tong [11] | Vision: face orientation recognition | Stranger face. | Own face stable faster than stranger. |
| Keenan [35] | Vision: face recognition | Familiar colleagues, stranger face. | Upright and inverted faces, self-faces were significantly faster than familiars and stranger faces. | |
| Sui [36] | Vision: face reaction | Familiar, stranger face. | Recognition speed: self (551 ms), familiar (596 ms), unfamiliar face (588 ms). | |
| Jie [37] | Vision: face recognition | Familiar, disordered face. | Recognition speed: own face stable faster than stranger.Recognition accuracy: own face is higher than familiar and unfamiliar faces. | |
| Ma [38] | Vision: face orientation recognition | Familiars, disordered face. | The speed of self is faster than that of acquaintances and disordered faces, and it has the advantage of rapid recognition in both explicit and implicit experiments. | |
| Harris [39] | Vision: Name search | Interference name, ordinary words. | The names of 60 subjects were detected faster than other identities. The author believed that the subjects’ names were a ‘high-priority‘ text stimulus. | |
| Eye track | Wang [40] | Vision: Name search | Mother, celebrity, common name. | The average number of saccades of one’s own name is 1.1 times less than that of a mother’s and a celebrity’s name, and the first fixation of one’s own name is 170 ms faster than that of other names. |
| EEG | Tacikowski [41] | Vision: name recognition | Celebrity, stranger name. | The P300 of one’s own name has a shorter latency period. |
| State | Author | Experiment | Control Group | Conclusion |
|---|---|---|---|---|
| Distraction | Moray [4] | Auditory: name recognition by the following ear | None | The brain can process self-names in a distracted state. |
| Alexopoulos [23] | Auditory: name recognition | Acquaintance, stranger name | Self-attention is automatic, unintentional, unconscious, and uncontrolled. | |
| Pfister [24] | Vision: name recognition | Unrelated nouns, non-words | Self-names attract attention and were prioritized. | |
| Subliminal | Shapiro [25] | Vision: name recognition | Other names, common nouns | Own name shows stronger anti-interference ability. |
| Shelley [26] | Vision: Own name masked probability | Other symbols | In the masking task, the probability of own names was identified is higher. | |
| Vision: The interference target of its own names. | Other symbols | Self-information strongly affects the distribution of attention in both conscious and unconscious states. | ||
| Pannese [27] | Vision: Face and gender matching. | Acquaintance, celebrity, stranger face | Self-information acquires special cognitive processing, and self-face benefits from the early processing of the brain. | |
| Sun [28] | Vision: Gender judgment of name | Celebrity, stranger name | The accuracy of gender judgment of one’s own name under low load and short delay conditions is significantly better than that of other names. | |
| Interferent | S Brédart [29] | Vision: facial interference | Friend name | Self-face has stronger inhibition ability to target stimulus, and the interference effect of self-face is much stronger than that of friends. |
| Devue [30] | Vision: facial interference | Friend, stranger face | The own face can temporarily attract attention within the focus of attention. | |
| Devue [31] | Vision: facial interference | Friend, stranger face | The reaction time of the own face interference is longer, and the own face is more difficult to distract attention. | |
| Yamada [32] | Vision: Name interference | Other’s name | The distortion of vision space caused by the own name leads to the spatial distribution deviation of attention. | |
| Wolford [34] | Vision: Name interference and decision | Same surname string, blank word. | The strong interference of one’s own name to the main task has a lower perception threshold. | |
| Minimum consciousness | Ian [35] | Auditory: Name of sleep state | Classmate name | The extraordinary ability to remember the own name in sleep. |
| Perrin [36] | Auditory: name of sleep state | Other’s name | Own names cause N2, P3 to appear more frequently. | |
| Kurtz [38] | Auditory: Patient’s names during recovery and post-anesthesia | Noise | The patient‘s reaction to their name is more obvious. | |
| Fishback [39] | Questionnaire: Cognitive tests of the elderly at different stages of AD | Time, place, count words. | With the development of dementia, the elderly finally forget their own name. | |
| Perrin [21] | Auditory: Name of sleep state | Other’s name | The brain’s cognitive response to one’s own name during sleep is similar to that during wakefulness. | |
| Pratt [40] | Auditory: Name of sleep state | Unrelated words | The feature of own names ERP show significant differences. | |
| Perrin | Auditory: the name of the minimally consciousness patient | Stranger name | The significant stimulus materials of own names have semantic processing. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Zeng, Y.; Tong, L.; Yan, B. Specific Neural Mechanisms of Self-Cognition and the Application of Brainprint Recognition. Biology 2023, 12, 486. https://doi.org/10.3390/biology12030486
Zhang R, Zeng Y, Tong L, Yan B. Specific Neural Mechanisms of Self-Cognition and the Application of Brainprint Recognition. Biology. 2023; 12(3):486. https://doi.org/10.3390/biology12030486
Chicago/Turabian StyleZhang, Rongkai, Ying Zeng, Li Tong, and Bin Yan. 2023. "Specific Neural Mechanisms of Self-Cognition and the Application of Brainprint Recognition" Biology 12, no. 3: 486. https://doi.org/10.3390/biology12030486