
Long-Tailed Macaque (Macaca fascicularis) Contraception Methods: A Systematic Review
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Methodology
1.2. Information Sources
1.3. Eligibility and Rejection Criteria
1.4. Selection Process
1.5. Method of Data Collection and Synthesis
2. Results
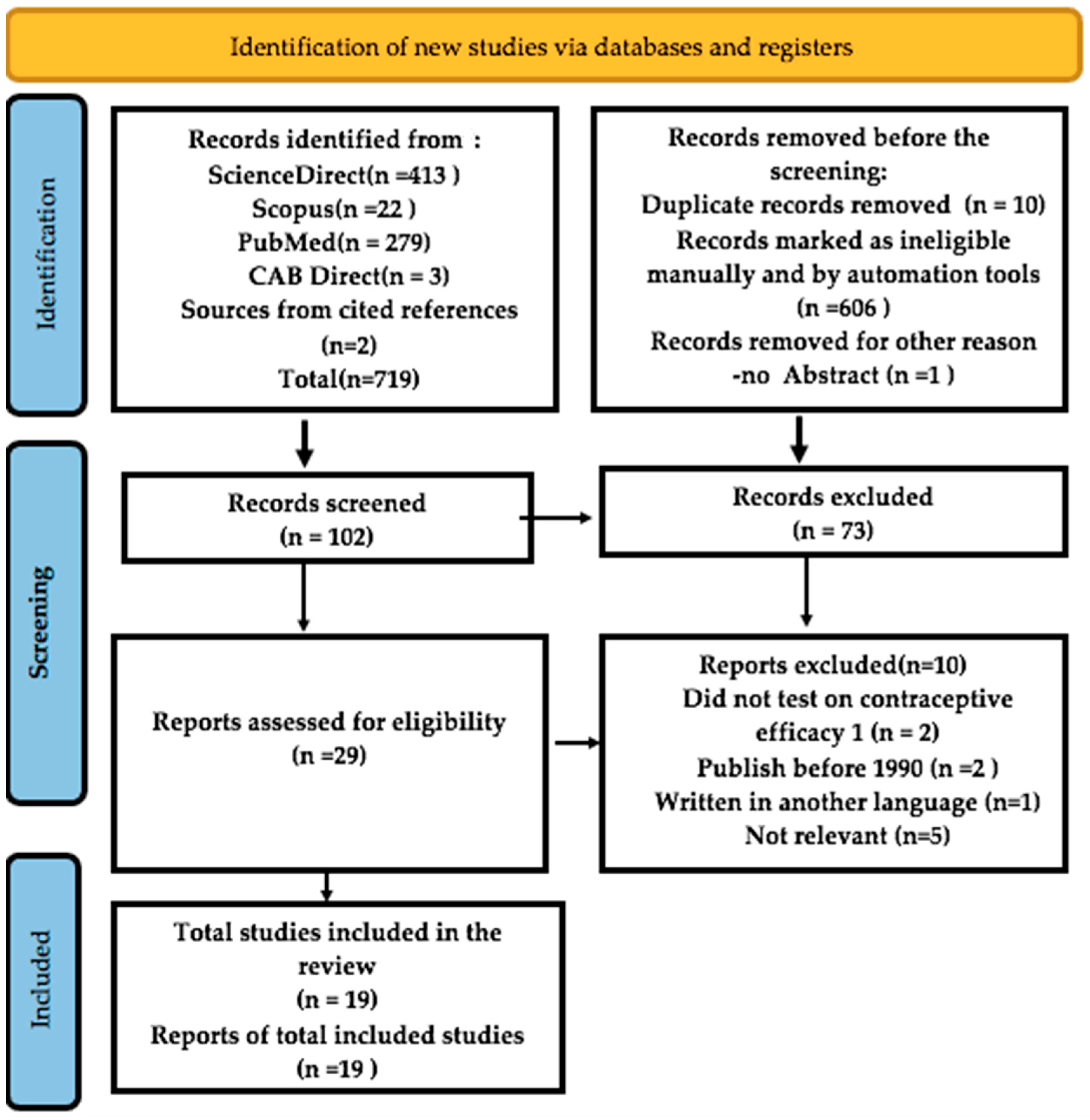
2.1. Study Selection
2.2. Characteristics of the Included Articles
2.3. Hormonal Contraception
2.4. Non-Hormonal Contraception
2.5. Male Non-Hormonal Contraception
2.6. Female Non-Hormonal Contraception

{kind=link}
| Type of Contraception and Reference | Dose, Route of Administration and Duration | Mechanism of Action | Sample Composition | Studies Outcome/Efficacy |
|---|---|---|---|---|
| Female hormonal contraception | ||||
| [64] Levonorgestrel diet | (i) Pellet containing 0, 8, 20, 50 or 125 μg LNG daily from day 2 to 15 of the menstrual cycle (ii) 50 μg LNG Infusion into feeding pellet and orally administered daily on days 9–22, 16–29 or 23–36 of the menstrual cycle | It causes the elevation of FSH and LH and decreases E2 to inhibit ovulation in cynomolgus macaque by affecting the folliculogenesis and hypothalamus | Total sample size n = 22 (i) n = 4 in each treatment group, and control (ii) n = 4 each treatment days | (1). 8/22 (36.3%) treated @ 20 and 50 μg LNG experienced delayed ovulation (2). 4/22 (18.18%) treated @ 125 μg LNG experienced inhibited ovulation. (3). 4/22 (18.18%) treated @ 8 μg LNG failed to inhibit ovulation. (4). 4/22 (18.18%) control had normal ovulation. (5). 16/16 (100%) treated @ 50 μg LNG daily experienced delayed ovulation in all groups. |
| [33] Progesterone (Medroxyprogesterone acetate (MPA)) | Single subcutaneous injection @ 15 mg/kg | Exerts its mechanism of contraception through the inhibition of follicular maturation and or inhibition of ovulation | Total sample n = 8 Treatment group n = 5 Control group n = 3 | (1). 3/8 (37.5%) of control had no changes in the endocrine and menstruation profile. (2). 5/8 (62.5%) of treated showed complete cessation of menstrual cycles. |
| [34] Antiprogestin (RU 486) | Antiprogestin 5 mg/day, dexamethasone I mg/day and 0.5 mL ethanol, SID from cycle day 10 to 12 | (61) indicates that RU 486 exerts its effect by acting as a progesterone antagonist, thereby affecting the physiologic pre-ovulatory gonadotropin surges | Total sample size n = 18 Treatment group (1) n = 5 Treatment group (2) n = 3 Treatment group (3) n = 3 Control group n = 5 | (1). 5/18 (27.7%) of treated group 1 and 3/18 (16.6%) of treated group 2 showed a delayed rise in gonadotropin, delayed folliculogenesis and extended intermenstrual interval. While 3/18 (16.6%) of treated group 3 and 5/18 (27.2%) of control had a normal midcycle surge of gonadotropins. |
| [35] Antiestrogen (Tamoxifen) | A single oral dose of 5 mg/kg on day four post-ovulation | It exerts its antiestrogenic effect by competing with the endogenous estrogen for the estrogen–receptor complex | Total sample size n = 26 Treatment group n = 13 Control group n = 13 | (1). In 6/13 (46%) of the treatment group, the cynomolgus monkey became pregnant while (2). 4/13 (31%) of the control group became pregnant |
| [36] Antiprogestin (Anordrin) | (i) 4.0 and 8.0 mg/kg b.w IM on days 9–13 of the menstrual cycle 0.1 or 0.2 mg/kg b.w IM within the first three days of the menstrual cycle | It exerts its effect through the inhibition of follicular development and subsequently inhibits ovulation | Total sample size n = 32 Control n = 8 (1) Treatment n = 8 (2) Treatment n = 7 (3) Treatment n = 3 (4) Treatment n = 5 | (1). 8/32 (25%) of control values observed within the normal range. (2). 8/32 (25%) of treatment 1 and 7/32 (21.8%) of treatment 2 did not inhibit luteal activity and rather delayed the development of ovarian follicles. (3). 3/32 (9.3%) of treatment 3 observed the absence of menstruation for 5–6 months. (4). 5/32 (15.6%) of treatment 4 showed delayed follicular maturation with an increase in the length of luteal phase. |
| [38] Antiprogestin RU 486 | 1 mg/kg/day of RU486 IM for three consecutive cycles | RU 486 inhibits ovulation in cynomolgus macaque at the level of hypothalamic–pituitary axis by interfering with the feedback of ovarian periovolatory signals | n = 6 in 3 menstrual cycle (treatment, rest and control) | (1). In 6/6 (100%) of the treatment cycle, ovulation was inhibited during RU486 administration. (2). In 5/6 (83.3%) progesterone was undetected, while 1/6 (16.6%) had increased progesterone. |
| Male hormonal contraception | ||||
| [41] GnRH antagonist plus testosterone (Deterelix) | Silastic capsule implant (crystalline T) SC 5 days before injecting the GnRH antagonist GnRH antagonist injection @ 250 μg/kg, 750 μg/kg day | Exerts its action on cynomolgus monkeys through the reversible suppression of serum T, LH, FSH and inhibin levels and subsequently causes azoospermia due to combination of testosterone (Deterelix) | Total sample size n = 22 1. Treatment 1 n = 5 @ 250 μg/kg + Sham Implant 2. Treatment 2 n = 5@ 250 μg/kg+ T 3. Treatment 3 n = 5@ 750 μg/kg + T 4. Control n = 7 | In animals treated with an antagoninst, 14/15 representing 93% became azoospermic including 9/10 of treated with antagonist plus T representing 90%. No changes observed in the control group. |
| [42] Testosterone bucilate (TB) | Control received diluent Treatment group 1 received TB@ 10 mg/kg Treatment group 2 received TB @ 20 mg/kg IM | Testicular involution is achieved through the complete suppression of FSH secretion | Total sample size n = 15 Control group n = 5 Treatment 1 group n = 5 Treatment 2 group n = 5 | 1. 5/15 (33.3%) of control had average LH levels and serum testosterone was within the normal range. 2. In 10/15 (66.6%) of both treated groups, the LH level dropped while 5/15 (33.3%) of the treated had a rise in serum testosterone level up to 4-fold and 5/15 (33.3%) of the treated had up to a 5-fold rise in serum testosterone. |
| Type of Contraception and Reference | Dose, Route of Administration, Duration | Mechanism of Action | Sample Size | Studies Outcomes |
|---|---|---|---|---|
| Male non-hormonal contraception | ||||
| [43] Triptonide (Tripterygium wilfordii) | 0.1 mg/kg BW single oral dose | Exert its mechanism of action at its target site, the plakoglobin junction, during spermiogenesis disrupt its interaction with SPEM1. It causes deformed sperm, with compromised forward motility and low sperm count after 5 week of treatment | Total sample size n = 4 Control n = 2 + 2 female Treatment n = 2 + 2 female Each group was housed with 2 proven fertile females | 2/4 (50%) of the treated had no pregnancy. 2/4 (50%) of the control had two pregnancies. |
| [47] EP055 | IV infusion of a single dose of 63.25 mg/kg | It exerts its effect by targeting the sperm protein EPPIN on the surface of the spermatozoa to inhibit sperm motility | Total sample size n = 5 Infusion (IV)Treatment n = 3 Tissue level n = 2 | 3/5 (60%) of the infusion had 10.6 min half-life for EP055. 2/5 (40%) of the tissue demonstrated EP055 in the testis and epididymis 2–6 h after infusion. |
| Female non-hormonal contraception | ||||
| [26] Surgical approach | NA | Through laparoscopy. To carry out tubectomy | n = 140 | No new pregnancy was recorded among the sterilized females, with a 96.3% survival rate. |
| [48] NSAIDs (Meloxicam) | Oral administration of meloxicam @ 0.5 mg/kg body weight/day in food for five days at mid-follicular, late follicular and periovulatory phases | Meloxicam in this study is thought to exert its contraceptive action by inhibiting the activity of ovarian prostaglandin, which affects ovulation by preventing follicular rupture and prevents the release of oocytes | Total sample size n = 24 Cycle 1(vehicle treatment) n = 4 × 3 group Cycle 2 (meloxicam treatment) n = 4 × 3 group Cycle 3 (vehicle oocyte) n = 4 × 3 group Cycle 4 (meloxicam oocytes) n = 3, n = 4 and n = 3 | In both cycles 1 and 2, no significant differences were observed. In cycle 3, no oocytes in the luteinized follicles were observed. In cycle 4 8/11, 72.7% of oocytes were observed in the luteinized follicles. |
| [53] Phosphodiesterase (PDE) 3 inhibitor ORG 9935 | Given orally @ 150 mg/kg SID or depot injection @ 150 mg/kg weekly for seven ovulatory cycles | ORG 9935 exerts its reversible contraceptive effect at a serum concentration above 300 nmol/L by inhibiting meiotic maturation of the oocyte without affecting ovulation or the normal function of the corpus luteum | Total sample size n = 16 Control n = 8 Treatment n = 8 | In the control group, 7/8 become pregnant, representing 88% of the pregnancy rate. In the treatment group, 4/8 become pregnant, representing 50% of the pregnancy rate. |
| [25] Surgical approach | NA | Via laparoscopy to carry out tubectomy | N = 1343 comprising both cynomolgus and rhesus macaque | During the monitoring period from 2009 to 2019, none of the sterilized animals became pregnant, which was evidenced by the decrease in the population growth from under 60% to 30%. |
| [56] NSAID (Meloxicam) | Oral administration @ 0.5 mg/kg SID for five days after mating | Through the inhibition of prostaglandin synthesis, which interrupts the release of oocyte necessary for ovulation | Total sample size for emergency model n = 11 46 meloxicam treatment cycle 15 control cycle | In the oral vehicle, no meloxicam, the pregnancy rate was 5/15 (33%), while in meloxicam treatment group, the pregnancy rate was 3/46 (6.6%). |
| [57] Prostaglandin E2 receptor antagonist (PTGER2)/BAY06 | Subcutaneously @ 10 mg/kg in 0.5 mL castor oil given BID for six months | BAY06 is thought to exert its effect through the inhibition of cumulus–oocyte expansion | Total sample size n = 19 Treatment group n = 9 Control n = 10 | In the control, the pregnancy rate was 8/10, representing 80% pregnancy, while in the treated group, the pregnancy rate was 2/9, representing 22% pregnancy. |
| [58] Pueraria Mirifica (A Thai herb) | Suspension @ 10, 100, and 1000 mg/5 mL of distilled water/individual/day in 3 treatment group of macaque | The mechanism of its action involves the mediation of the estrogen receptor at the hypothalamic–pituitary–gonadal axis and the uterus, which reduces FSH and LH levels in the hypothalamus through a negative feedback mechanism. These factors subsequently influence the menstrual cycle and inhibit ovulation | Total sample size n = 9 Treatment group 1 @ 10 mg/5 mL n = 3 Treatment group 2 @ 100 mg/5 mL n = 3 Treatment group 3 @ 1000 mg/5 mL n = 3 | 1. 6/9 (66.6%) of treatment groups 1 and 2 had extended menstrual cycle length with recovery of hormones levels post-treatment. However, group 2 showed suppression of FSH and LH. 2. 3/9 (33.3%) of treatment group 3 showed complete cessation of the menstrual cycle and suppression of FSH and LH. |
| [60] Lignosulfonic acid | Involve treatment of sperm with 1.5 mg/mL before washing or after capacitation to evaluate sperm–oocyte interaction with a zona pellucida binding assay and invitro fertilization | It exerts its potential as a vaginal contraceptive by binding to the head of capacitated sperm, thereby inhibiting the binding of sperm to the zona pellucida | NA | 92.5% of treatment with LSA inhibited sperm zona binding post-capacitation, while 82.5% inhibited sperm zona binding before washing. For in vitro fertilization, 19/19 (100%) of pre and post-capacitation treatment of sperm with LSA prevented fertilization following on the 19 oocytes, while 11/18 (61.1%) of control oocytes were fertilized |
| [63] Immunocontraception (Porcine Zona Pellucida) | 360 μg IM on cycle day 26, followed by a booster of 180 μg IM at week two after the first two doses and four weeks apart from the rest | Immunocontraception with porcine zona pellucida in cynomolgus monkeys exerts contraception only when there is a sustained serum antibody level for several weeks | Total sample size n = 12 All animals were immunized with porcine zona pellucida | 6/12 (50%) of immunized animals conceived, while 6/12 (50%) did not conceive. |
3. Discussion
4. Study Limitation
5. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liedigk, R.; Kolleck, J.; Böker, K.O.; Meijaard, E.; Md-Zain, B.M.; Abdul-Latiff, M.A.; Ampeng, A.; Lakim, M.; Abdul-Patah, P.; Tosi, A.J.; et al. Mitogenomic phylogeny of the common long-tailed macaque (Macaca fascicularis fascicularis). BMC Genom. 2015, 16, 222. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, Y.; Su, H.H. Macaques as seed dispersal agents in Asian forests: A review. Int. J. Primatol. 2018, 39, 356–376. [Google Scholar] [CrossRef]
- Fauzi, R.; Wuryanto, T.; Suarmadi, F.; Tomonob, A. Distribution of long-tailed macaque (Macaca fascicularis) in Kelimutu National Park. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 591, p. 012041. [Google Scholar]
- Fooden, J. Comparative review of fascicularis-group species of macaques (primates: Macaca). Fieldiana Zool. 2006, 107, 1–43. [Google Scholar]
- Fooden, J. Systematic review of Southeast Asia long-tail macaques, Macaca fascicularis (Raffles, 1821). Fieldiana Zool. 1995, 81, 1–206. [Google Scholar]
- Karuppannan, K.; Saaban, S.; Mustapa, A.R.; Zainal Abidin, F.A.; Azimat, N.A.; Keliang, C.J. Population status of long-tailed macaque (Macaca fascicularis) in Peninsular Malaysia. J. Primatol. 2014, 3, 34–37. [Google Scholar]
- Kaewchot, S.; Tangsudjai, S.; Sariya, L.; Mongkolphan, C.; Saechin, A.; Sariwongchan, R.; Suksai, P. Zoonotic pathogens survey in free-living long-tailed macaques in Thailand. Int. J. Vet. Sci. Med. 2022, 10, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Bushmitz, M.; Lecu, A.; Verreck, F.; Preussing, E.; Rensing, S.; Mätz-Rensing, K. Guidelines for the prevention and control of tuberculosis in non-human primates: Recommendations of the European Primate Veterinary Association Working Group on Tuberculosis. J. Med. Primatol. 2009, 38, 59–69. [Google Scholar] [CrossRef]
- Paungpin, W.; Thongdee, M.; Ketchim, N.; Chaiwattanarungruengpaisan, S.; Saechin, A.; Sariya, L.; Wiriyarat, W. Evidence of Influenza A Virus Infection in Cynomolgus Macaques, Thailand. Vet. Sci. 2022, 9, 132. [Google Scholar] [CrossRef]
- Barua, M.; Bhagwat, S.A.; Jadhav, S. The hidden dimensions of human-wildlife conflict: Health impacts, opportunity and transaction costs. Biol. Conserv. 2013, 157, 309–316. [Google Scholar] [CrossRef]
- Nyhus, P.J. Human-wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 2016, 41, 143–171. [Google Scholar] [CrossRef] [Green Version]
- Linnell, J.D.C.; Rondeau, D.; Reed, D.H.; Williams, R.; Altwegg, R.; Raxworthy, C.J.; Austin, J.D.; Hanley, N.; Fritz, H.; Evans, D.M.; et al. Confronting the costs and conflicts associated with biodiversity. Anim. Conserv. 2010, 13, 429–431. [Google Scholar] [CrossRef]
- Plowman, A.B.; Jordan, N.R.; Anderson, N.; Condon, E.; Fraser, O. Welfare implications of captive primate population management: Behavioural and psychosocial effects of female-based contraception, oestrus and male removal in hamadryas baboons (Papio hamadryas). Appl. Anim. Behav. Sci. 2005, 90, 155–165. [Google Scholar] [CrossRef]
- Al-Jazeera. Massive Malaysian Monkey Cull Spurs Concerns. Available online: https://www.aljazeera.com/in-depth/features/2013/04/2013422143220434441.Html (accessed on 1 November 2022).
- IPPL. International Primate Protection League Vol. 2018. Available online: https://www.ippl.org/gibbon/blog/Malaysia-monkey-war-take-action (accessed on 1 November 2022).
- Retnam, L.; Chatikavanij, P.; Kunjara, P.; Paramastri, Y.A.; Goh, Y.M.; Hussein, F.N.; Mutalib, A.R.; Poosala, S. Laws, Regulations, Guidelines and Standards for Animal Care and Use for Scientific Purposes in the Countries of Singapore, Thailand, Indonesia, Malaysia, and India. ILAR J. 2017, 57, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, K. Birth control in female Japanese macaques at Iwatayama Monkey Park, Arashiyama. In The Monkeys of Stormy Mountain: 60 Years of Primatological Research on the Japanese Macaques of Arashiyama; Cambridge University Press: Cambridge, UK, 2012; Volume 61, p. 435. [Google Scholar]
- Gray, M.E.; Cameron, E.Z. Does contraceptive treatment in wildlife result in side effects? A review of quantitative and anecdotal evidence. Reproduction 2010, 139, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Hecht, L. The importance of considering age when quantifying wild animals’ welfare. Biol. Rev. 2021, 96, 2602–2616. [Google Scholar] [CrossRef]
- Rhyan, J.C.; Miller, L.A.; Fagerstone, K.A. The use of contraception as a disease management tool in wildlife. J. Zoo Wildl. Med. 2013, 44, S135–S137. [Google Scholar] [CrossRef] [Green Version]
- Padodara, R.; Singh, V.; Odedara, A.; Vasava, A.; Sharma, A.; Mehta, V. Modern approaches to contraception in domestic and wild animals: A review. J. Glob. Ecol. Environ. 2022, 16, 14–25. [Google Scholar]
- Nya-Ngatchou, J.J.; Amory, J.K. New approaches to male non-hormonal contraception. Contraception 2013, 87, 296–299. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 88, 105906. [Google Scholar]
- Hong, Q.N.; Fàbregues, S.; Bartlett, G.; Boardman, F.; Cargo, M.; Dagenais, P.; Gagnon, M.P.; Griffiths, F.; Nicolau, B.; O’Cathain, A.; et al. The Mixed Methods Appraisal Tool (MMAT) version 2018 for information professionals and researchers. Educ. Inf. 2018, 34, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Martelli, P.; Krishnasamy, K.; Kwan, A.; Wong, A. Permanent contraception by laparoscopic tubectomy with ovarian conservation in Hong Kong macaques. Jpn. J. Vet. Res. 2020, 68, 209–215. [Google Scholar]
- Deleuze, S.; Brotcorne, F.; Polet, R.; Soma, G.; Rigaux, G.; Giraud, G.; Cloutier, F.; Poncin, P.; Wandia, N.; Huynen, M.C. Tubectomy of Pregnant and Non-pregnant Female Balinese Macaques (Macaca Fascicularis) with Post-operative Monitoring. Front. Vet. Sci. 2021, 8, 688656. [Google Scholar] [CrossRef] [PubMed]
- Henzl, M.R. Contraceptive hormones and their clinical use. In Reproductive Endocrinology. Physiology, Pathophysiology and Clinical Management; Yen, S.S.C., Jaffe, R.B., Eds.; WB Saunders Company: Philadelphia, PA, USA, 1991; pp. 807–829. [Google Scholar]
- Henderson, J.A.; Shively, C.A. Triphasic oral contraceptive treatment alters the behavior and neurobiology of female cynomolgus monkeys. Psychoneuroendocrinology 2004, 29, 21–34. [Google Scholar] [CrossRef]
- Zimmerman, Y.; Eijkemans, M.J.C.; Coelingh Bennink, H.J.T.; Blankenstein, M.A.; Fauser, B.C.J.M. The effect of combined oral contraception on testosterone levels in healthy women: A systematic review and meta-analysis. Hum. Reprod. 2014, 20, 76–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aden, U.; Jung-Hoffmann, C.; Kuhl, H. A randomized cross-over study on various hormonal parameters of two triphasic oral contraceptives. Contraception 1998, 58, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Carr, B.R.; Griffin, J.E. Fertility control and its complications. In Williams Textbook of Endocrinology, 9th ed.; Wilson, J.D., Foster, D.W., Kronenberg, H.M., Larsen, P.R., Eds.; W.B. Saunders: Philadelphia, PA, USA, 1998; pp. 901–926. [Google Scholar]
- Nayak, N.R.; Ghosh, D.; Lasley, B.L.; Sengupta, J. Anti-implantation activity of luteal phase mifepristone administration is not mimicked by prostaglandin synthesis inhibitor or prostaglandin analog in the rhesus monkey. Contraception 1997, 55, 103–114. [Google Scholar] [CrossRef]
- Shimizu, K.; Takenoshita, Y.; Mitsunaga, F.; Nozaki, M. Suppression of ovarian function and successful contraception in macaque monkeys following a single injection of medroxyprogesterone acetate. J. Reprod. Dev. 1996, 42, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Collins, R.L.; Hodgen, G.D. Blockade of the Spontaneous Midcycle Gonadotropin Surge in Monkeys by RU 486: A Progesterone Antagonist or Agonist. J. Clin. Endocrinol. Metab. 1986, 63, 1270–1276. [Google Scholar] [CrossRef]
- Tarantal, A.F.; Hendrickx, A.G.; Matlin, S.A.; Lasley, B.L.; Gu, Q.Q.; Thomas, C.A.; Vince, P.M.; Van Look, P.F. Tamoxifen as an antifertility agent in the long-tailed macaque (Macaca fascicularis). Contraception 1993, 47, 307–316. [Google Scholar] [CrossRef]
- Mehta, R.R.; Chatterton, R.T. Antiovulatory action of anordrin in the cynomolgus monkey (Macaca fascicularis). Adv. Contracept 1991, 7, 21–28. [Google Scholar] [CrossRef]
- Hu, C.-Y.; Wang, W.-C.; Hwang, T.-M.; Chen, C.-H.; Lu, S.-H.; Hu, H.-E.; Char, S.-S.; Fan, B.-C.; Hew, W.-L. Endometrial and hormonal changes induced by anordrin. In WHO Symposium on Steroid Contraception and Mechanisms of Endometrial Bleeding; Diczfalusy, I.S., Frazer, I.S., Webb, F.T., Eds.; Pitman Press Ltd.: Bath, UK, 1980; pp. 191–200. [Google Scholar]
- Heikinheimo, O.; Gordon, K.; Lähteenmäki, P.; Williams, R.F.; Hodgen, G.D. D. Antiovulatory actions of RU 486: The pituitary is not the primary site of action in vivo. J. Clin. Endocrinol. Metab. 1995, 80, 1859–1868. [Google Scholar] [PubMed]
- Ledger, W.; Sweeting, V.; Hillier, H.; Baird, D. Inhibition of ovulation by low dose mifepristone (RU 486). Hum. Reprod. 1992, 7, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Batista, M.; Cartledge, T.; Zellmer, A.; Nieman, L.; Merriam, G.; Loriaux, L. Evidence for the critical role of progesterone in the regulation of the midcycle gonadotropin surge and ovulation. J. Clin. Endocrinol. Metab. 1992, 74, 565–570. [Google Scholar] [PubMed]
- Bremner, W.J.; Bagatell, C.J.; Steiner, R.A. Gonadotropin-releasing hormone antagonist plus testosterone: A potential male contraceptive. J. Clin. Endocrinol. Metab. 1991, 73, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, G.F.; Schlatt, S.; Walter, V.; Nieschlag, E. Testosterone-induced inhibition of spermatogenesis is more closely related to the suppression of FSH than to testicular androgen levels in the cynomolgus monkey model (Macaca fascicularis). J. Endocrinol. 2001, 168, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Qin, W.; Zheng, H.; Schegg, K.; Han, L.; Liu, X.; Wang, Y.; Wang, Z.; McSwiggin, H.; Peng, H.; et al. Triptonide is a reversible non-hormonal male contraceptive agent in mice and non-human primates. Nat. Commun. 2021, 12, 1253. [Google Scholar] [CrossRef] [PubMed]
- Matlin, S.A.; Belenguer, A.; Stacey, V.E.; Qlan, S.Z.; Xu, Y.; Zhang, J.W.; Sanders, J.K.; Amor, S.R.; Pearce, C.M. Male Antifertility Compd. Tripterygium Wilfordii Hook. Contraception 1993, 47, 387–400. [Google Scholar] [CrossRef]
- Bao, J.; Dai, S.M. A Chinese herb Tripterygium wilfordii Hook F in the treatment of rheumatoid arthritis: Mechanism, efficacy, and safety. Rheumatol. Int. 2011, 31, 1123–1129. [Google Scholar] [CrossRef]
- Tao, X.; Lipsky, P.E. The Chinese anti-inflammatory and immunosuppressive herbal remedy Tripterygium wilfordii Hook F. Rheum. Dis. Clin. N. Am. 2000, 26, 29–50. [Google Scholar]
- O’Rand, M.G.; Hamil, K.G.; Adevai, T.; Zelinski, M. Inhibition of sperm motility in male macaques with EP055, a potential non-hormonal male contraceptive. PLoS ONE 2018, 13, e0195953. [Google Scholar] [CrossRef] [Green Version]
- Hester, K.E.; Harper, M.J.; Duffy, D.M. Oral administration of the cyclooxygenase-2 (COX-2) inhibitor meloxicam blocks ovulation in non-human primates when administered to simulate emergency contraception. Hum. Reprod. 2010, 25, 360–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendonca, L.L.; Khamashta, M.A.; Nelson-Piercy, C.; Hunt, B.J.; Hughes, G.R. Non-steroidal anti-inflammatory drugs as a possible cause of reversible infertility. Rheumatology 2000, 39, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Pall, M.; Friden, B.E.; Brannstrom, M. Induction of delayed follicular rupture in humans by the selective COX-2 inhibitor rofecoxib: A randomized, double-blind study. Hum. Reprod. 2001, 16, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Bata, M.S.; Al-Ramahi, M.; Salhab, A.S.; Gharaibeh, M.N.; Schwartz, J. Delay of ovulation by meloxicam in healthy cycling volunteers: A placebo-controlled, double-blind, crossover study. J. Clin. Pharmacol. 2006, 46, 925–932. [Google Scholar] [CrossRef]
- Jensen, J.T.; Zelinski, M.B.; Stanley, J.E.; Fanton, J.W.; Stouffer, R.L. The phosphodiesterase 3 inhibitor ORG 9935 inhibits oocyte maturation in the naturally selected dominant follicle in rhesus macaques. Contraception 2008, 77, 303–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesan, C.; Salvatierra, A.M.; Schwartz, J.; Croxatto, H.B. Suppression of follicular rupture with meloxicam, a cyclooxygenase inhibitor: Potential for emergency contraception. Hum. Reprod. 2010, 25, 368–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, J.T.; Schwinof, K.M.; Zelinski-Wooten, M.B.; Conti, M.; DePaolo, L.V.; Stouffer, R.L. Phosphodiesterase 3 inhibitors selectively block the spontaneous resumption of meiosis by macaque oocytes in vitro. Hum. Reprod. 2002, 17, 2079–2084. [Google Scholar] [CrossRef]
- Nogueira, D.; Albano, C.; Adriaenssens, T.; Cortvrindt, R.; Bourgain, C.; Devroey, P.; Smitz, J. Human oocytes reversibly arrested in prophase I by phosphodiesterase type 3 inhibitor in vitro. Biol. Reprod. 2003, 69, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- McCann, N.C.; Lynch, T.J.; Kim, S.O.; Duffy, D.M. The COX-2 inhibitor meloxicam prevents pregnancy when administered as an emergency contraceptive to non-human primates. Contraception 2013, 88, 744–748. [Google Scholar] [CrossRef]
- Peluffo, M.C.; Stanley, J.; Braeuer, N.; Rotgeri, A.; Fritzemeier, K.H.; Fuhrmann, U.; Stouffer, R.L. A prostaglandin E2 receptor antagonist prevents pregnancies during a preclinical contraceptive trial with female macaques. Hum. Reprod. 2014, 29, 1400–1412. [Google Scholar] [CrossRef] [Green Version]
- Trisomboon, H.; Malaivijitnond, S.; Watanabe, G.; Taya, K. Ovulation block by Pueraria mirifica: A study of its endocrinological effect in female monkeys. Endocrine 2005, 26, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Chansakaow, S.; Ishikawa, T.; Sekine, K.; Okada, M.; Higuchi, Y.; Kudo, M.; Chaichantipyuth, C. Isoflavonoids from Pueraria Mirifica and their estrogenic activity. Planta Med. 2000, 66, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Tollner, T.L.; Overstreet, J.W.; Li, M.W.; Meyers, S.; Yudin, A.I.; Salinas, E.R.; Cherr, G.N. Lignosulfonic acid blocks in vitro fertilization of macaque oocytes when sperm are treated either before or after capacitation. J. Androl. 2002, 23, 889–898. [Google Scholar] [PubMed]
- Fraser, L.R. Sperm capacitation and the acrosome reaction. Hum. Reprod. 1998, 13 (Suppl. S1), 9–19. [Google Scholar] [CrossRef] [Green Version]
- Youssef, H.M.; Doncel, G.F.; Bassiouni, B.A.; Acosta, A.A. Effect of sperm viability, plasmalemma integrity, and capacitation on patterns of ex- pression of mannose-binding sites on human sperm. Arch. Androl. 1997, 38, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Gulyas, B.J.; Gwatkin, R.B.; Yuan, L.C. Active immunization of cynomolgus monkeys (Macaca fascicularis) with porcine zonae pellucidae. Gamete Res. 1983, 7, 299–307. [Google Scholar] [CrossRef]
- Nakama, K.; Akune, A.; Kawate, N.; Takahashi, M.; Inaba, T.; Sameshima, H.; Tamada, H. Delay of ovulation due to diets containing levonorgestrel in cynomolgus monkeys (Macaca fascicularis). J. Vet. Med. Sci. 2012, 74, 1453–1460. [Google Scholar] [CrossRef] [Green Version]
- Wallace, P.Y.; Asa, C.S.; Agnew, M.; Cheyne, S.M. A review of population control methods in captive-housed primates. Anim. Welf. 2016, 25, 7–20. [Google Scholar] [CrossRef]
- Kirkpatrick, J.F.; Turner, J.W. Reversible fertility in non-domestic animals. J. Zoo Wildl. Med. 1991, 22, 392–408. [Google Scholar]
- Bomford, M.; O’Brien, P. Potential use of contraception for managing wildlife pests in Australia. In Contraception in Wildlife Management. USD-APHIS Technical Bulletin 1853; Kreeger, T.J., Ed.; USDA-APHIS: Washington, DC, USA, 1997; pp. 205–214. [Google Scholar]
- Fraker, M.A.; Brown, R.G.; Gaunt, G.E.; Kerr, J.A.; Pohajdak, B. Long-lasting, single-dose immunocontraception of feral fallow deer in British Columbia. J. Wildl. Manag. 2002, 1141–1147. [Google Scholar] [CrossRef]
- Turck, D.; Roth, W.; Busch, U. A review of the clinical pharmacokinetics of meloxicam. Br. J. Rheumatol. 1996, 35, 13–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeNicola, A.J.; VerCauteren, K.C.; Curtis, P.D.; Hygnstrom, S.E. (Eds.) Managing White-tailed Deer in Suburban Environments: A Technical Guide; Cornell Cooperative Extension: Ithaca, NY, USA, 2000. [Google Scholar]
- Delsink, A.K.; van Altena, J.J.; Grobler, D.; Bertschinger, H.; Kirkpatrick, J.; Slotow, R. Implementing immunocontraception in free-ranging African elephants at Makalali Conservancy. J. S. Afr. Vet. Assoc. 2007, 78, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delsink, A.K.; Kirkpatrick, J. (Eds.) Free-Ranging African Elephant Immunocontraception; Trident Press: Cape Town, South Africa, 2012. [Google Scholar]
- Kirkpatrick, J.F.; Rowan, A.; Lamberski, N.; Wallace, R.; Frank, K.; Lyda, R. The practical side of immunocontraception: Zona proteins and wildlife. J. Reprod. Immunol. 2009, 83, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Fagerstone, K.A.; Miller, L.A.; Bynum, K.S.; Eisemann, J.D.; Yoder, C.A. When, Where and for What Wildlife Species Will Contraception Be a Useful Management Approach? Proc. Vertebr. Pest Conf. 2006, 22, 22. [Google Scholar] [CrossRef] [Green Version]
- Hampton, J.O.; Hyndman, T.H.; Barnes, A.; Collins, T. Is wildlife fertility control always humane? Animals 2015, 5, 1047–1071. [Google Scholar] [CrossRef]
- Kirkpatrick, J.F. Measuring the effects of wildlife contraception: The argument for comparing apples with oranges. Reprod. Fertil. Dev. 2007, 19, 548–552. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, J.F.; Turner, J.W. Chemical Fertility Control and Wildlife Management. BioScience 1985, 35, 485–491. [Google Scholar] [CrossRef]
- Cowan, D.P.; Massei, G. Wildlife contraception, individuals, and populations: How much fertility control is enough? Proc. Vertebr. Pest Conf. 2008, 23, 23. [Google Scholar] [CrossRef] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikail, M.; Azizan, T.R.P.T.; Noor, M.H.M.; Hassim, H.A.; Che’Amat, A.; Latip, M.Q.A. Long-Tailed Macaque (Macaca fascicularis) Contraception Methods: A Systematic Review. Biology 2023, 12, 848. https://doi.org/10.3390/biology12060848
Mikail M, Azizan TRPT, Noor MHM, Hassim HA, Che’Amat A, Latip MQA. Long-Tailed Macaque (Macaca fascicularis) Contraception Methods: A Systematic Review. Biology. 2023; 12(6):848. https://doi.org/10.3390/biology12060848
Chicago/Turabian StyleMikail, Muhammed, Tengku Rinalfi Putra Tengku Azizan, Mohd Hezmee Mohd Noor, Hasliza Abu Hassim, Azlan Che’Amat, and Mohd Qayyum Ab Latip. 2023. "Long-Tailed Macaque (Macaca fascicularis) Contraception Methods: A Systematic Review" Biology 12, no. 6: 848. https://doi.org/10.3390/biology12060848