
Testing the Challenge Hypothesis in Stumptail Macaque Males: The Role of Testosterone and Glucocorticoid Metabolites in Aggressive and Mating Behavior
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Housing Conditions
2.2. Fecal Sample Collection
2.3. Hormone Extraction and Quantification
2.4. Behavioral Analyses
2.5. Data Analyses
3. Results
3.1. Aggression
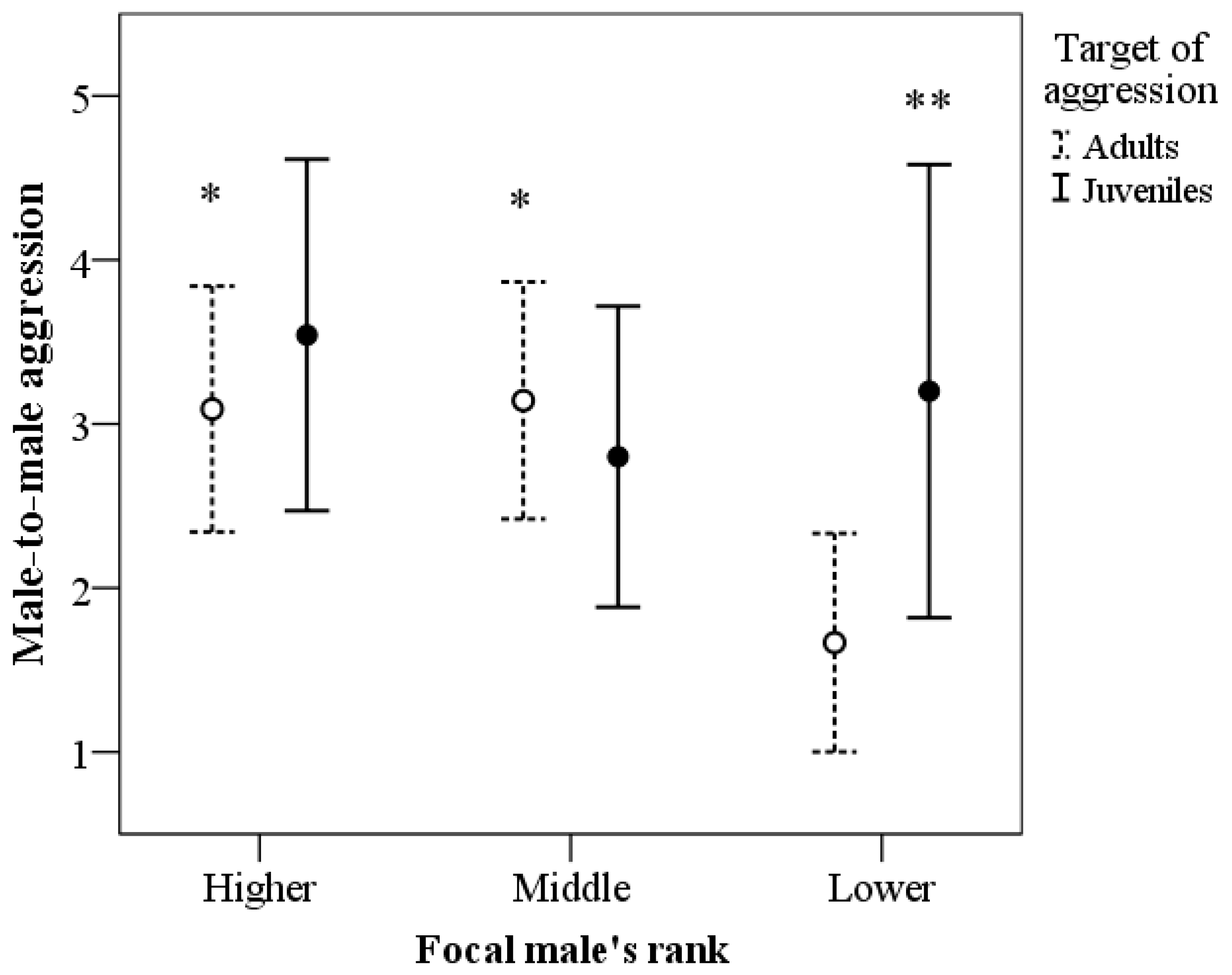
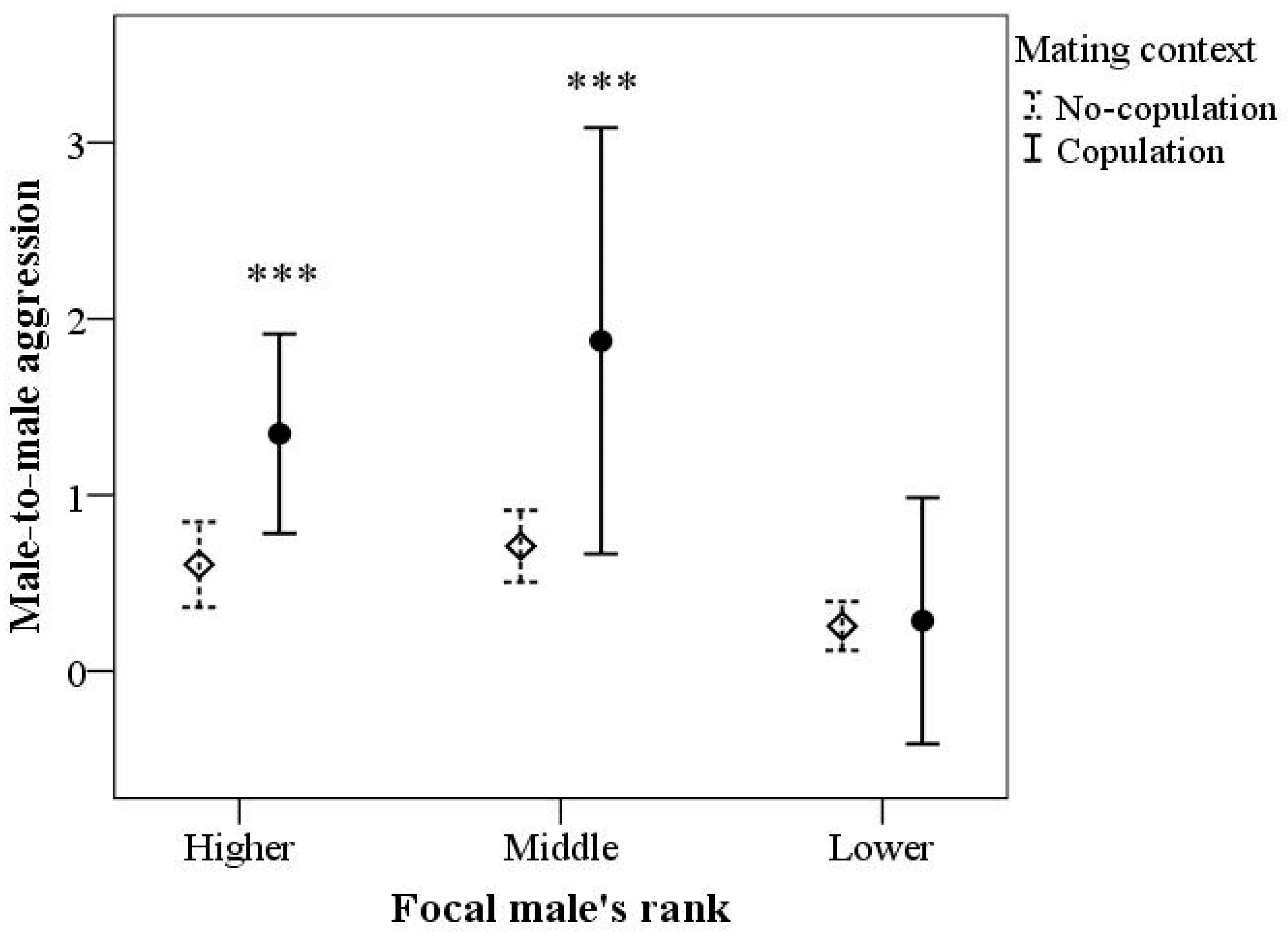
3.2. Male-to-Male Aggression
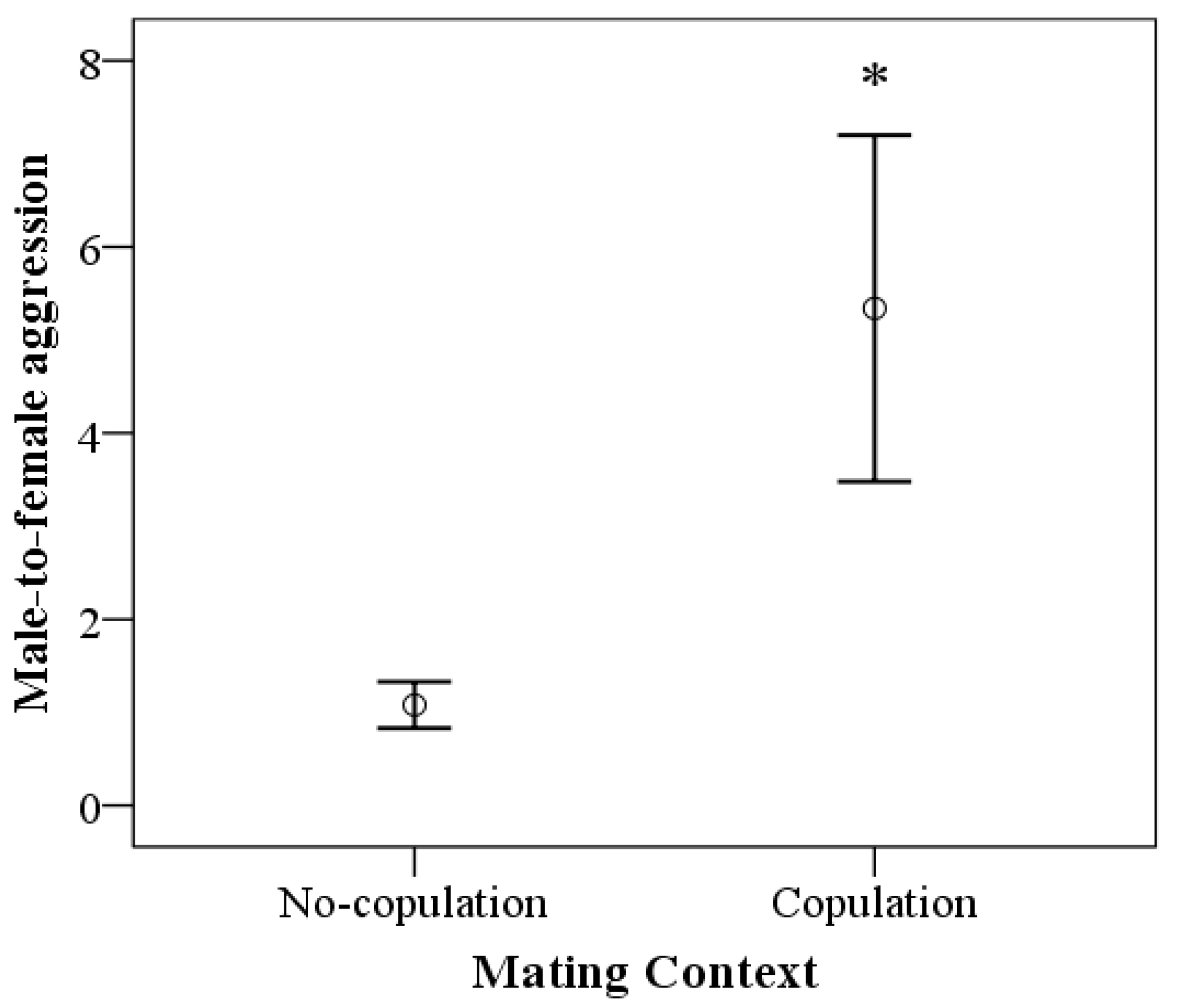
3.3. Male-to-Female Aggression
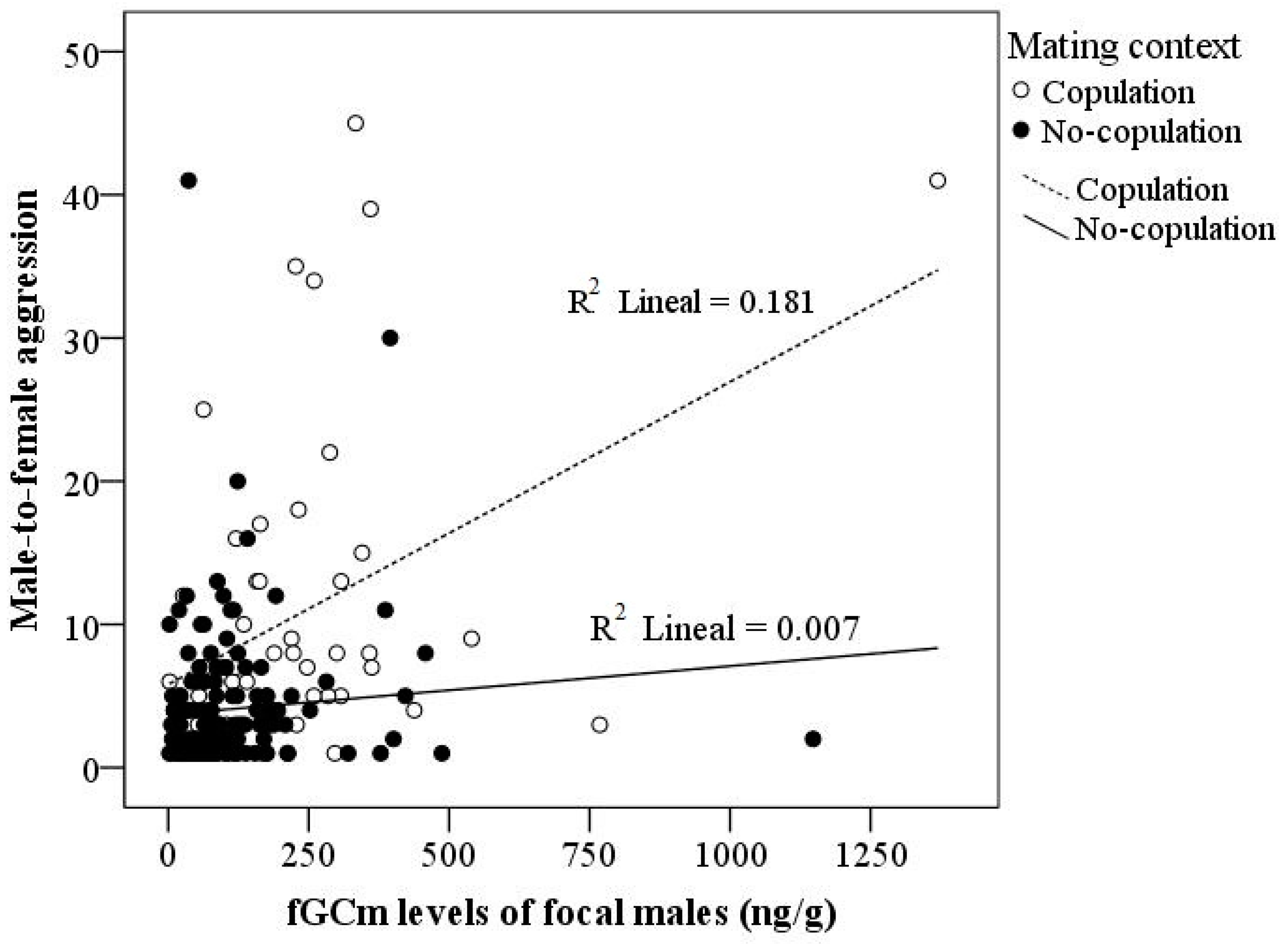
3.4. Predictors of fTm and fGCm Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Honess, P.E.; Marin, C.M. Behavioural and physiological aspects of stress and aggression in nonhuman primates. Neurosci. Biobehav. Rev. 2006, 30, 390–412. [Google Scholar] [CrossRef]
- Georgiev, A.V.; Klimczuk, A.C.E.; Traficonte, D.M.; Maestripieri, D. When violence pays: A cost-benefit analysis of aggressive behavior in animals and humans. Evol. Psychol. 2013, 11, 678–699. [Google Scholar] [CrossRef] [Green Version]
- Silverberg, J.; Gray, J.P. Violence and peacefulness as behavioral potentialities of primates. In Aggression and Peacefulness in Humans and Other Primates; Silverberg, J., Gray, J.P., Eds.; Oxford University Press: New York, NY, USA, 1992; pp. 1–36. [Google Scholar]
- Maynard Smith, J.; Parker, G.A. The logic of asymmetric contest. Anim. Behav. 1976, 24, 159–175. [Google Scholar] [CrossRef]
- Caperos, J.M.; Sánchez, S.; Peláez, F.; Fidalgo, A.; Morcillo, A. The effect of crowding on the social behavior of the cooperatively breeding cotton-top tamarins (Saguinus oedipus). Int. J. Primatol. 2011, 32, 1179–1189. [Google Scholar] [CrossRef]
- Andrade, M.A.; Oliveira, D.; Mazzochini, G.G.; Amado, T.F.; Gouveia, S.F.; Martinez, P.A. Drivers of intraspecific lethal aggression in mammals. J. Biogeogr. 2021, 48, 2445–2454. [Google Scholar] [CrossRef]
- Rosell, D.R.; Siever, L.J. The neurobiology of aggression and violence. CNS Spectr. 2015, 20, 254–279. [Google Scholar] [CrossRef] [PubMed]
- Higley, J.D. Aggression. In Primate Psychology; Maestripieri, D., Ed.; Harvard University Press: Cambridge, MA, USA, 2003; pp. 17–40. [Google Scholar]
- Fanning, J.R.; Keedy, S.; Berman, M.E.; Lee, R.; Coccaro, E.F. Neural correlates of aggressive behavior in real time: A review of fMRI studies of laboratory reactive aggression. Curr. Behav. Neurosci. Rep. 2017, 4, 138–150. [Google Scholar] [CrossRef]
- Archer, J. Testosterone and human aggression: An evaluation of the challenge hypothesis. Neurosci. Biobehav. Rev. 2006, 30, 319–345. [Google Scholar] [CrossRef] [PubMed]
- Higham, J.P.; Heistermann, M.; Maestripieri, D. The endocrinology of male rhesus macaque social and reproductive status: A test of the challenge and social stress hypotheses. Behav. Ecol. Sociobiol. 2013, 67, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Wingfield, J.C.; Hegner, R.E.; Dufty, A.M.; Ball, G.F. The “Challenge hypothesis”: Theoretical implications for patterns of testosterone secretion, mating systems, and breeding strategies. Am. Nat. 1990, 136, 829–846. [Google Scholar] [CrossRef]
- Cavigelli, S.A.; Pereira, M.E. Mating season aggression and fecal testosterone levels in male ring-tailed lemurs (Lemur catta). Horm. Behav. 2000, 37, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.N. Testosterone and reproductive effort in male primates. Horm. Behav. 2017, 91, 36–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapolsky, R. Stress-induced elevation of testosterone concentrations in high-ranking baboons: Role of catecholamines. Endocrinology 1986, 118, 1630–1635. [Google Scholar] [CrossRef]
- Ketterson, E.D. Testosterone and avian life histories effects of experimentally elevated testosterone on behaviour and correlates of fitness in the dark-eye junco (Junco hyemalis). Am. Nat. 1992, 140, 980–999. [Google Scholar] [CrossRef]
- Simon, N.G.; Lu, S.H. Androgens and Aggression. In Biology of Aggression; Nelson, R.J., Ed.; Oxford University Press: New York, NY, USA, 2006; pp. 211–230. [Google Scholar]
- Girard-Buttoz, C.; Heistermann, M.; Krummel, S.; Engelhardt, A. Seasonal and social influences on fecal androgen and glucocorticoid excretion in wild male long-tailed macaques (Macaca fascicularis). Physiol. Behav. 2009, 98, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Ostner, J.; Heistermann, M.; Schülke, O. Male competition and its hormonal correlates in Assamese macaques (Macaca assamensis). Horm. Behav. 2011, 59, 105–113. [Google Scholar] [CrossRef]
- Rangel-Negrin, A.; Dias, P.A.; Chavira, R.; Canales-Espinosa, D. Social modulation of testosterone levels in male black howlers (Alouatta pigra). Horm. Behav. 2011, 59, 159–166. [Google Scholar] [CrossRef]
- Xia, D.P.; Li, J.H.; Sun, B.H.; Weed, J.L.; Kyes, R.C. Evaluation of fecal testosterone, rank and copulatory behavior in wild male Macaca thibetana at Huangshan, China. Pak. J. Zool. 2015, 47, 1445–1454. [Google Scholar]
- Muller, M.N.; Wrangham, R.W. Dominance, aggression and testosterone in wild chimpanzees: A test of the ‘challenge’ hypothesis. Anim. Behav. 2004, 67, 113–123. [Google Scholar] [CrossRef]
- Muller, M.N.; Wrangham, R.W. Dominance, cortisol and stress in wild chimpanzees (Pan troglodytes schweinfurthii). Behav. Ecol. Sociobiol. 2004, 55, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.L.; Maestripieri, D.; Wallen, K. Estrogen increases female sexual initiation independent of male responsiveness in rhesus monkeys. Horm. Behav. 1998, 33, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Gettler, L.T.; McDade, T.W.; Feranil, A.B.; Kuzawa, C.W. Longitudinal evidence that fatherhood decreases testosterone in human males. Proc. Natl. Acad. Sci. USA 2011, 108, 16194–16199. [Google Scholar] [CrossRef] [Green Version]
- Bales, K.L.; French, J.A.; McWilliams, J.; Lake, R.A.; Dietz, J.M. Effects of social status, age, and season on androgen and cortisol levels in wild male golden lion tamarins (Leontopithecus rosalia). Horm. Behav. 2006, 49, 88–95. [Google Scholar] [CrossRef]
- Hoffman, C.L.; Ruiz-Lambides, A.V.; Davila, E.; Maldonado, E.; Gerald, M.S.; Maestripieri, D. Sex differences in survival costs of reproduction in a promiscuous primate. Behav. Ecol. Sociobiol. 2008, 62, 1711–1718. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R. Endocrine aspects of social instability in the olive baboon (Papio anubis). Am. J. Primatol. 1983, 5, 365–379. [Google Scholar] [CrossRef]
- Sapolsky, R. The physiology of dominance in stable versus unstable social hierarchies. In Primate Social Conflict; Mason, W.A., Mendoza, S.P., Eds.; New York Press: New York, NY, USA, 1993; pp. 171–204. [Google Scholar]
- Sapolsky, R.M. The influence of social hierarchy on primate health. Science 2005, 308, 648–652. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive suppressive, stimulatory and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [Green Version]
- Higham, J.P.; Maestripieri, D. Revolutionary coalitions in male rhesus macaques. Behaviour 2010, 147, 1889–1908. [Google Scholar] [CrossRef] [Green Version]
- Kalbitzer, U.; Heistermann, M.; Cheney, D.; Seyfarth, R.; Fischer, J. Social behavior and patterns of testosterone and glucocorticoid levels differ between male chacma and Guinea baboons. Horm. Behav. 2015, 75, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostner, J.; Heistermann, M.; Schülke, O. Dominance, aggression and physiological stress in wild male Assamese macaques (Macaca assamensis). Horm. Behav. 2008, 54, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Gesquiere, L.R.; Learn, N.H.; Simao, M.C.M.; Onyango, P.O.; Alberts, S.C.; Altmann, J. Life at the Top: Rank and Stress in Wild Male Baboons. Science 2011, 333, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, C.; Mevis, L.; Malaivijitnond, S.; Schülke, O.; Ostner, J. Social relationships in free-ranging male Macaca arctoides. Int. J. Primatol. 2009, 30, 625–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuijsen, K.; de Neef, K.J.; Van der Werff ten Bosch, J.J.; Slob, A.K. Testosterone, testis size, seasonality and behavior in group-living stumptail macaques (Macaca arctoides). Horm. Behav. 1985, 21, 153–169. [Google Scholar] [CrossRef] [PubMed]
- López-Vergara, L.; Santillan-Doherty, A.M.; Mayagoitia, L.; Mondragon-Ceballos, R. Self and social grooming in stump-tailed macaques: Effects of kin presence or absence within the group. Behav. Process. 1989, 18, 99–106. [Google Scholar] [CrossRef]
- Pineda-Galindo, E.; Cerda-Molina, A.L.; Mayagoitia-Novales, L.; Matamoros-Trejo, G. Biological validations of fecal glucocorticoid, testosterone, and progesterone metabolite measurements in captive stumptail macaques (Macaca arctoides). Int. J. Primatol. 2017, 38, 985–1001. [Google Scholar] [CrossRef]
- Maestripieri, D. Social communication among captive stump-tailed macaques (Macaca arctoides). Int. J. Primatol. 1996, 17, 785–802. [Google Scholar] [CrossRef]
- Maestripieri, D. Gestural communication in three species of macaques (Macaca mulatta, M. nemestrina, M. arctoides): Use of signals in relation to dominance and social context. Gesture 2005, 5, 57–73. [Google Scholar] [CrossRef]
- de la O-Rodríguez, C.; Mevis, L.; Richter, C.; Malaivijitnond, S.; Ostner, J.; Schülke, O. Reconciliation in Male Stump-tailed Macaques (Macaca arctoides): Intolerant Males Care for Their Social Relationships. Ethology 2013, 119, 39–51. [Google Scholar] [CrossRef]
- Thierry, B. Unity in diversity: Lessons from macaque societies. Evol. Anthropol. 2007, 16, 224–238. [Google Scholar] [CrossRef]
- Estrada, A.; Estrada, R. Birth and breeding cyclicity in an outdoor living stumptail macaque (Macaca arctoides) group. Primates 1976, 17, 225–231. [Google Scholar] [CrossRef]
- Smith, E.O. Non seasonal breeding patterns in Stumptail Macaques (Macaca arctoides). Primates 1984, 25, 117–122. [Google Scholar] [CrossRef]
- Cerda-Molina, A.L.; Mayagoitia-Novales, L.; de la O-Rodríguez, C.; Borráz-León, J.I.; Matamoros-Trejo, G. Socio-sexual behaviors and fecal hormone metabolites but not age predict female aggressive interactions in Macaca arctoides. Behav. Ecol. Sociobiol. 2022, 76, 16. [Google Scholar] [CrossRef]
- Fooden, J. The bear macaque, Macaca arctoides: A systematic review. J. Hum. Evol. 1990, 19, 607–686. [Google Scholar] [CrossRef]
- Nunn, C.L. The evolution of exaggerated sexual swellings in primates and the graded-signal hypothesis. Anim. Behav. 1999, 58, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Cerda-Molina, A.L.; Hernández-López, L.; Chavira, R.; Cardenas, M.; Paez-Ponce, D.; Cervantes-De la luz, H.; Mondragón-Ceballos, R. Endocrine changes in male stumptailed macaques (Macaca arctoides) as a response to odor stimulation with vaginal secretions. Horm. Behav. 2006, 49, 81–87. [Google Scholar] [CrossRef]
- Cerda-Molina, A.; Hernández-López, L.; Rojas-Maya, S.; Murcia-Mejía, C.; Mondragón-Ceballos, R. Male-induced sociosexual behavior by vaginal secretions in Macaca arctoides. Int. J. Primatol. 2006, 26, 791–807. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P. Measuring Behaviour. An Introductory Guide, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Bertrand, M. The behavioral repertoire of the stumptail macaque: A descriptive and comparative study. Bibl. Primatol. 1969, 11, 1–273. [Google Scholar]
- Santillán-Doherty, A.M.; Díaz, J.L.; Mondragón-Ceballos, R. Synergistic effects of kinship, sex and rank in the behavioural interactions of captive stump-tailed macaques. Folia Primatol. 1991, 56, 177–189. [Google Scholar] [CrossRef]
- de Vries, H.; Stevens, J.; Vervaecke, H. Measuring and testing the steepness of dominance hierarchies. Anim. Behav. 2006, 71, 585–592. [Google Scholar] [CrossRef] [Green Version]
- de Vries, H. An improved test of linearity in dominance hierarchies containing unknown or tied relationships. Anim. Behav. 1995, 50, 1375–1389. [Google Scholar] [CrossRef]
- de Vries, H. Finding a dominance order most consistent with a linear hierarchy: A new procedure and review. Anim. Behav. 1998, 55, 827–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curley, J.P. Compete: Organizing and Analyzing Social Dominance Hierarchy Data. R Package Version 0.1. 2016. Available online: https://www.r-pkg.org/pkg/compete (accessed on 1 November 2018).
- R Development Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://cran.rproject.org/bin/windows/base/old/3.5.1/ (accessed on 1 November 2018).
- Pan, W. Akaike’s information criterion in generalized estimating equations. Biometrics 2001, 57, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Pekár, S.; Brabec, M. Generalized estimating equations: A pragmatic and flexible approach to the marginal GLM modelling of correlated data in the behavioural sciences. Ethology 2018, 124, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Tweedie, M.C.K. An index which distinguishes between some important exponential families. In Statistics: Applications and New Directions, Proceedings of the Indian Statistical Institute Golden Jubilee International Conference; Ghosh, J.K., Roy, J., Eds.; Indian Statistical Institute: Calcutta, India, 1984; pp. 579–604. [Google Scholar]
- Van Schaik, C.P.; Pradhan, G.R.; van Noordwijk, M.A. Mating conflict in primates: Infanticide, sexual harassment and female sexuality. In Sexual Selection in Primates: New and Comparative Perspectives; Kappeler, P.M., Van Schaik, C.P., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 131–150. [Google Scholar]
- Peckett, A.J.; Wright, D.C.; Riddell, M.C. The effects of glucocorticoids on adipose tissue lipid metabolism. Metab. Clin. Exp. 2011, 60, 1500–1510. [Google Scholar] [CrossRef]
- Schoof, V.A.; Jack, K.M.; Ziegler, T.E. Male response to female ovulation in white-faced capuchins (Cebus capucinus): Variation in fecal testosterone, dihydrotestosterone, and glucocorticoids. Int. J. Primatol. 2014, 35, 643–660. [Google Scholar] [CrossRef]
- Beehner, J.C.; Gesquiere, L.; Seyfarth, R.M.; Cheney, D.L.; Alberts, S.C.; Altmann, J. Testosterone related to age and life-history stages in male baboons and geladas. Horm. Behav. 2009, 56, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Reed, C.; O’brien, T.G.; Kinnaird, M.F. Male social behavior and dominance hierarchy in the Sulawesi crested black macaque (Macaca nigra). Int. J. Primatol. 1997, 18, 247–260. [Google Scholar] [CrossRef]
- Kimura, K.; Shimizu, K.; Hayashi, M.; Ishikawa, T.; Ago, Y. Pituitary–adrenocortical responses to the first dyadic encounters in male rhesus monkeys: Effect of dominance relationship. Am. J. Primatol. 2000, 50, 247–256. [Google Scholar] [CrossRef]
- Setchell, J.M.; Smith, T.; Wickings, E.J.; Knapp, L.A. Stress, social behaviour, and secondary sexual traits in a male primate. Horm. Behav. 2010, 58, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Bergman, T.J.; Beehner, J.C.; Cheney, D.L.; Seyfarth, R.M.; Whitten, P.L. Correlates of stress in free-ranging male chacma baboons, Papio hamadryas ursinus. Anim. Behav. 2005, 70, 703–713. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Age in Years (Class) | NormDS 2010 h’ = 0.75 a | NormDS 2011-1 h’ = 0.83 a | NormDS 2011-2 h’ = 0.63 b | No. of Fecal Samples |
|---|---|---|---|---|---|
| DF 1 | 10 (adult) | 9.62 | 9.62 | 8.61 | 104 |
| AL 1 | 20 (adult) | 8.44 | 8.60 | 7.85 | 117 |
| JI 2 | 16 (adult) | 6.58 | 7.21 | 5.59 | 110 |
| ES 2 | 20 (adult) | 5.60 | 6.42 | 5.57 | 129 |
| PO 2 | 22 (adult) | 5.03 | 4.44 | 5.10 | 57 |
| RP | 3 (juvenile) | 4.91 | 3.37 | 4.34 | |
| PL | 1 (infant) | 3.88 | 3.79 | 4.04 | |
| FO | 3 (juvenile) | 3.72 | 3.53 | 4.18 | |
| GA 3 | 16 (adult) | 3.40 | 3.27 | 3.95 | 87 |
| RE | 3 (juvenile) | 3.06 | 3.48 | 3.43 | |
| DW 3 | 23 (adult) | 0.75 | 1.29 | 2.33 | 96 |
| Focal Male-to-Male Aggression | Focal Male-to-Female Aggression | |||||
|---|---|---|---|---|---|---|
| Predictor | Wald χ2 | d.f. | p | Wald χ2 | d.f. | p |
| Interception | 0.464 | 1 | 0.496 | 0.341 | 1 | 0.560 |
| Mating context | 0.854 | 1 | 0.355 | 3.826 | 1 | 0.050 |
| Target of aggression | 68.257 | 1 | <0.001 | 8.724 | 1 | 0.003 |
| Focal male’s fTm | 2.597 | 1 | 0.107 | 2.832 | 1 | 0.092 |
| Focal male’s fGCm | 0.049 | 1 | 0.825 | 33.349 | 1 | <0.001 |
| Mating × rank | 14.113 | 2 | 0.001 | 1.851 | 2 | 0.829 |
| Mating × fTm | 1.239 | 1 | 0.266 | 0.000 | 1 | 0.985 |
| Mating × fGCm | 0.041 | 1 | 0.839 | 8.716 | 1 | 0.003 |
| Target of aggression × rank | 31.398 | 2 | <0.001 | 0.376 | 2 | 0.829 |
| Fecal Testosterone Metabolites | Fecal Glucocorticoid Metabolites | |||||
|---|---|---|---|---|---|---|
| Variable | Wald χ2 | d.f. | p | Wald χ2 | d.f. | p |
| Interception | 488.608 | 1 | 0.000 | 104.595 | 1 | 0.000 |
| Age | 0.901 | 1 | 0.343 | 0.151 | 1 | 0.697 |
| Rank | 3.112 | 2 | 0.211 | 48.422 | 2 | <0.001 |
| Mating Activity | 8.248 | 1 | 0.004 | 31.594 | 1 | <0.001 |
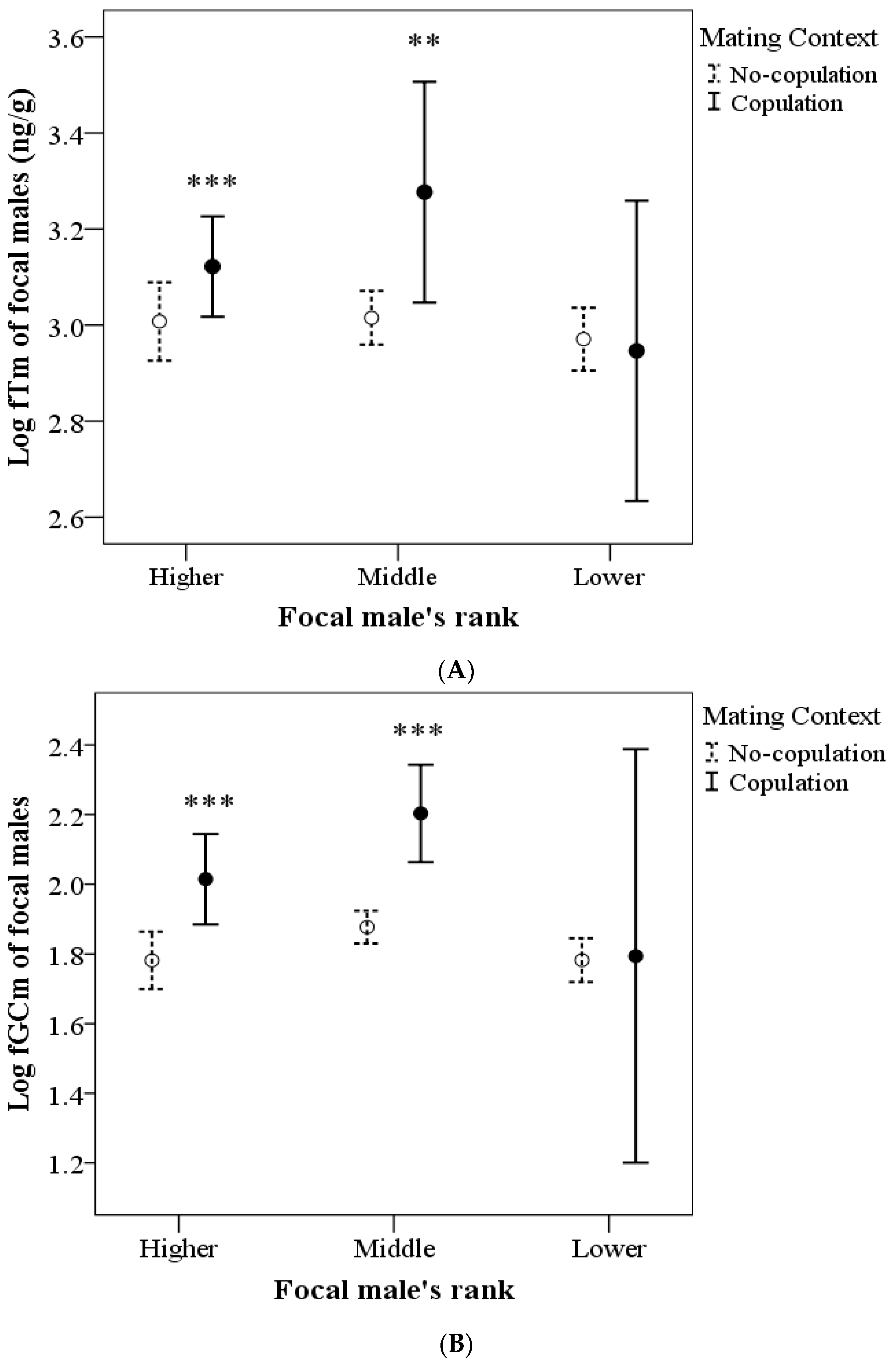
| Mating × rank | 6.072 | 2 | 0.042 | 9.305 | 2 | 0.010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerda-Molina, A.L.; Borráz-León, J.I.; Matamoros-Trejo, G.; de la O, C.; Estudillo-Mendoza, G.R.; Mayagoitia-Novales, L.; Maestripieri, D. Testing the Challenge Hypothesis in Stumptail Macaque Males: The Role of Testosterone and Glucocorticoid Metabolites in Aggressive and Mating Behavior. Biology 2023, 12, 813. https://doi.org/10.3390/biology12060813
Cerda-Molina AL, Borráz-León JI, Matamoros-Trejo G, de la O C, Estudillo-Mendoza GR, Mayagoitia-Novales L, Maestripieri D. Testing the Challenge Hypothesis in Stumptail Macaque Males: The Role of Testosterone and Glucocorticoid Metabolites in Aggressive and Mating Behavior. Biology. 2023; 12(6):813. https://doi.org/10.3390/biology12060813
Chicago/Turabian StyleCerda-Molina, Ana Lilia, Javier I. Borráz-León, Gilberto Matamoros-Trejo, Claudio de la O, Gema R. Estudillo-Mendoza, Lilian Mayagoitia-Novales, and Dario Maestripieri. 2023. "Testing the Challenge Hypothesis in Stumptail Macaque Males: The Role of Testosterone and Glucocorticoid Metabolites in Aggressive and Mating Behavior" Biology 12, no. 6: 813. https://doi.org/10.3390/biology12060813