
Oswaldocruzia ukrainae (Nematoda: Molineidae)—A Parasite of European Green Toad Bufotes viridis: Morphological and Molecular Data


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
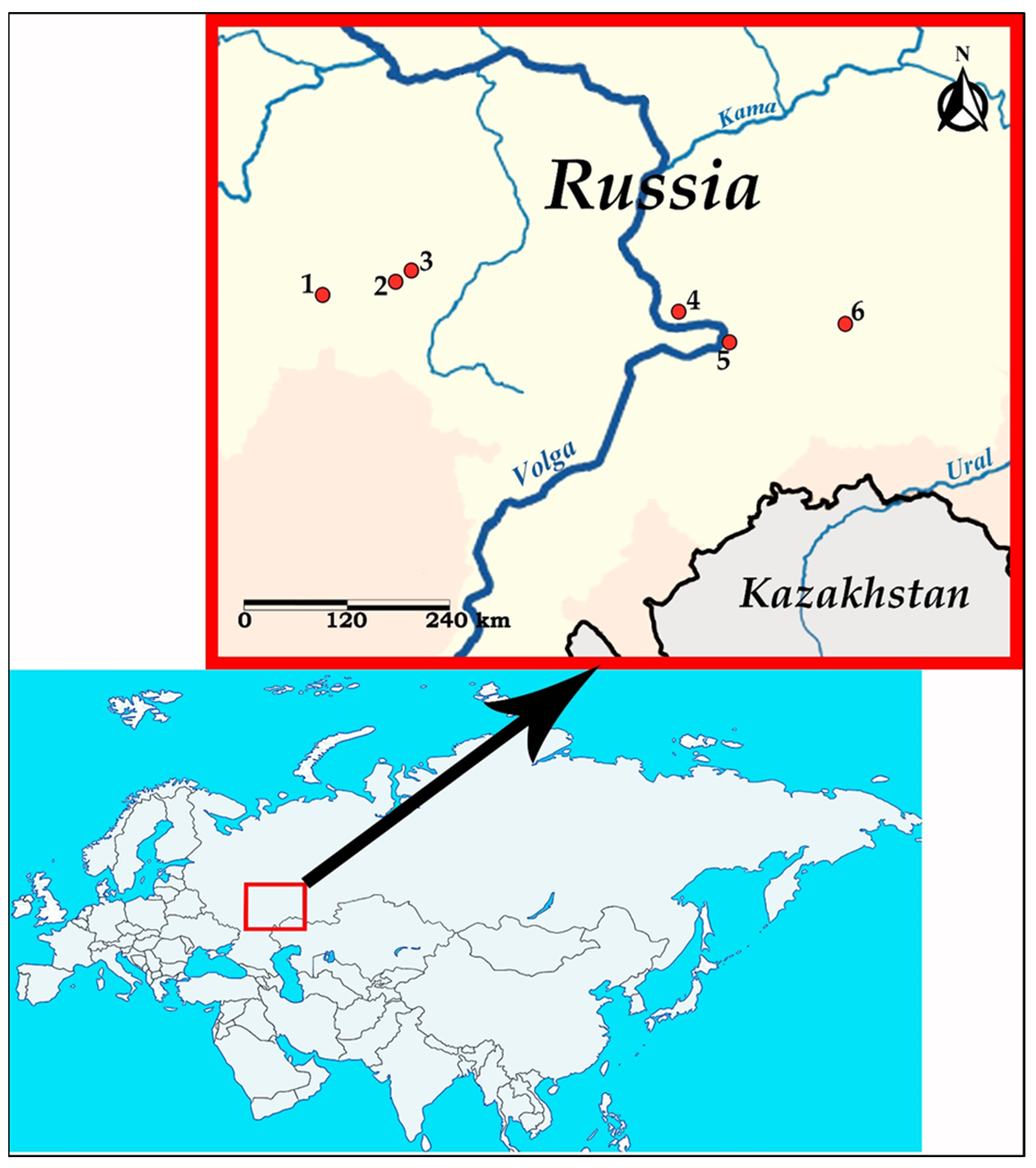
2.1. Nematode Collection and Examination
2.2. DNA Extraction, Amplification, Sequencing, and Phylogenetic Analysis
3. Results
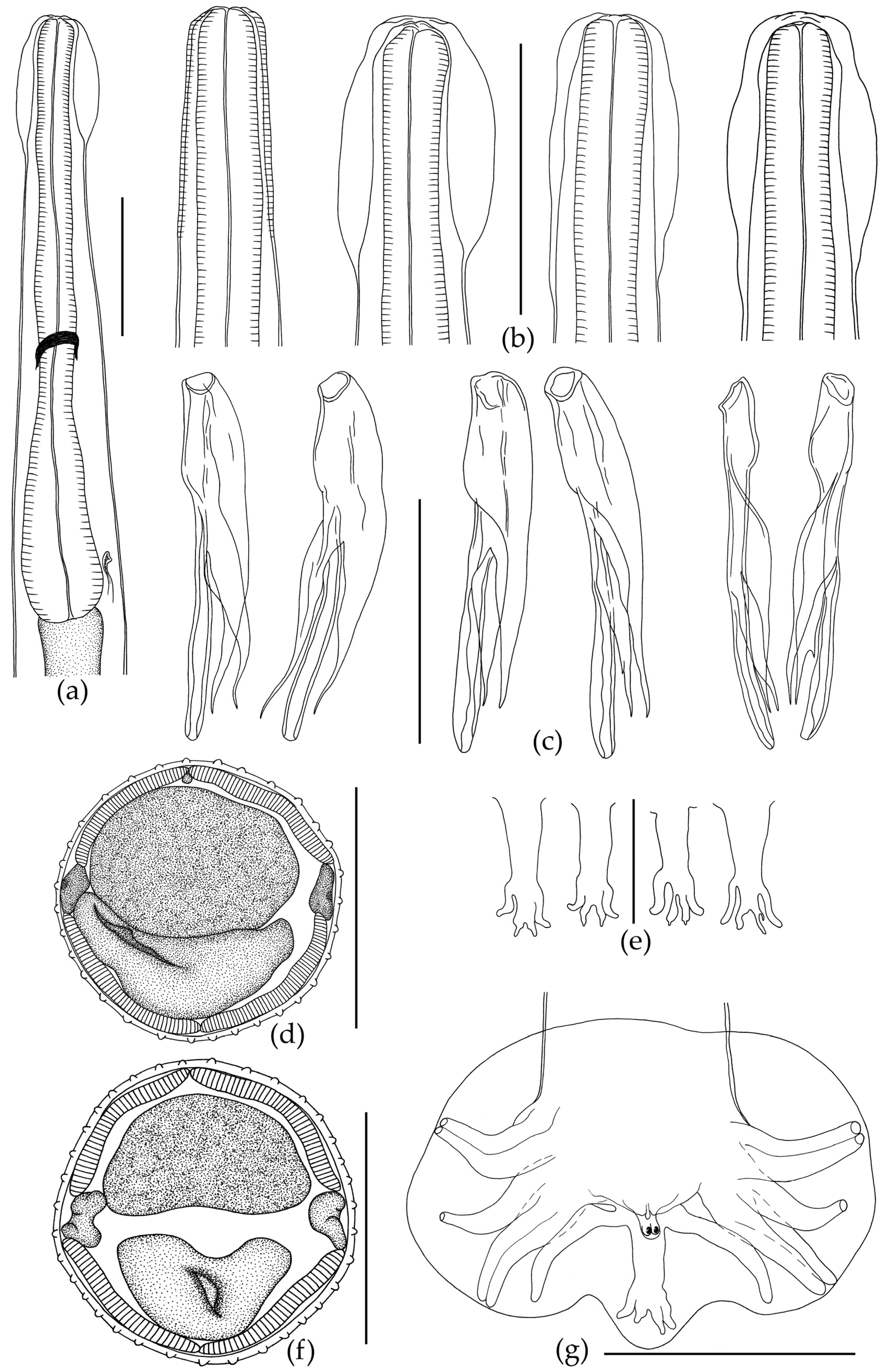
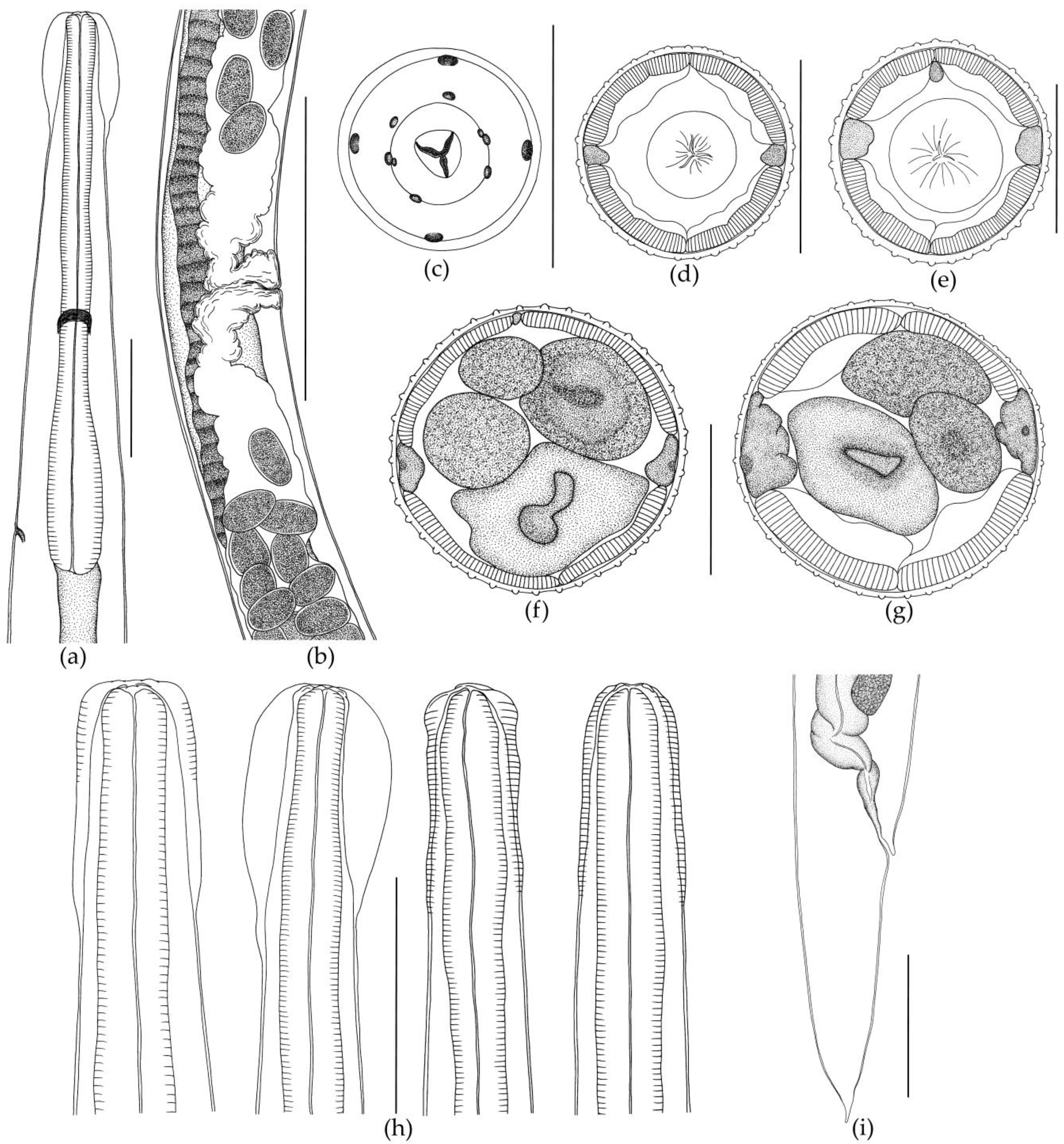
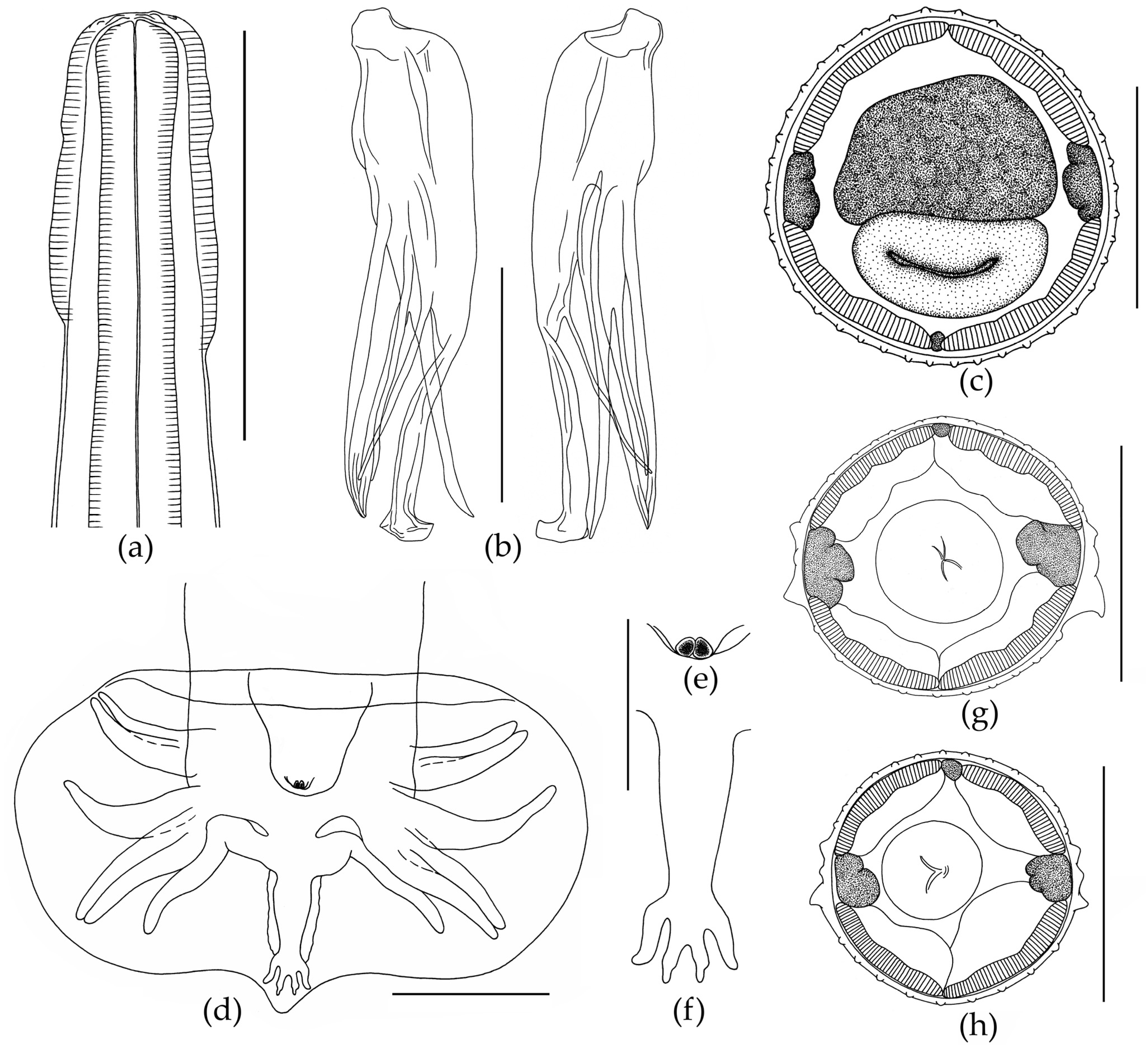
3.1. Morphological Description of Nematodes
3.1.1. Morphology of Oswaldocruzia ukrainae
3.1.2. Morphology of Oswaldocruzia filiformis Male
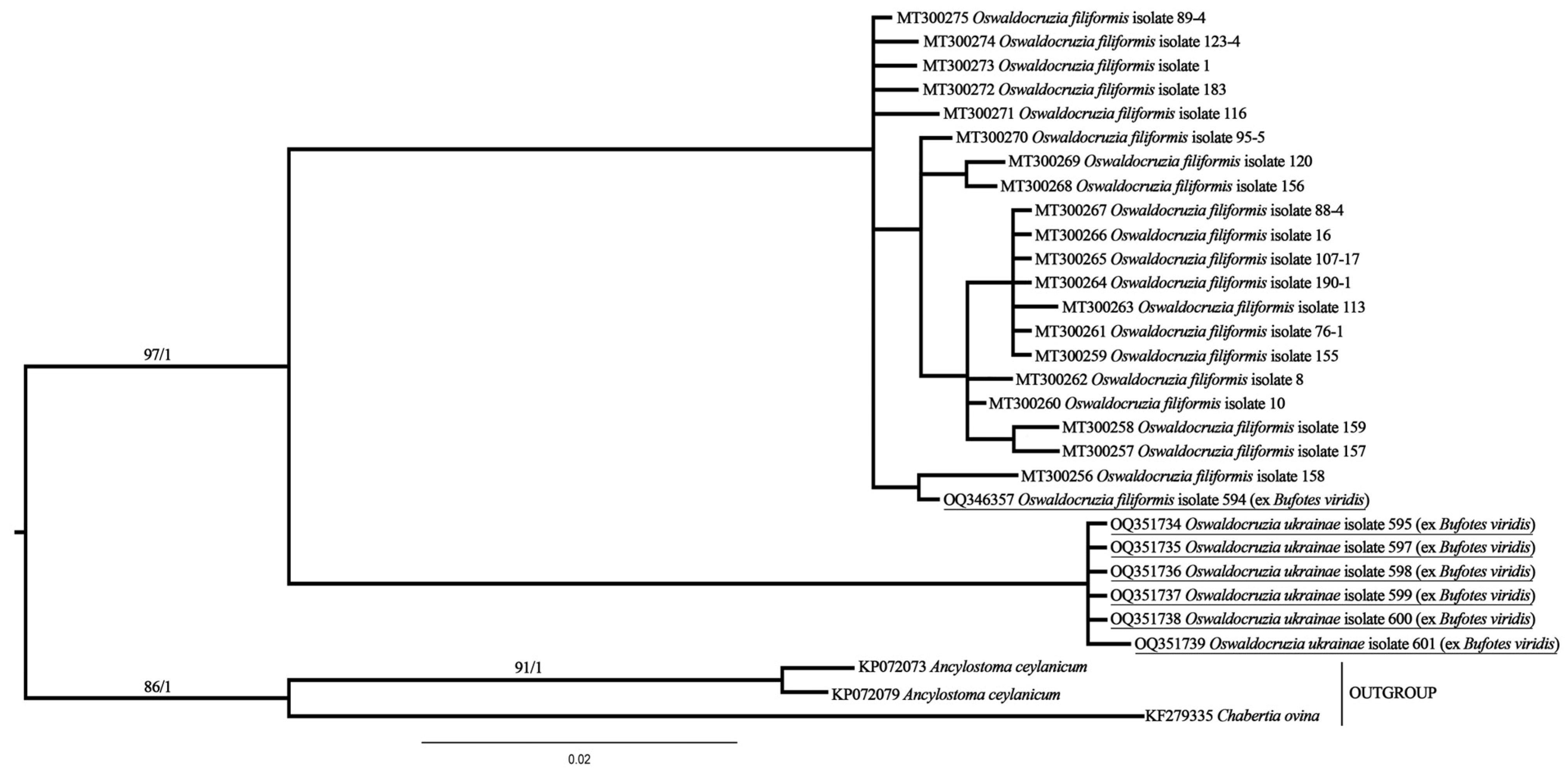
3.2. Molecular Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dufresnes, C.; Mazepa, G.; Jablonski, D.; Oliveira, R.C.; Wenseleers, T.; Shabanov, D.A.; Auer, M.; Ernst, R.; Koch, C.; Ramírez-Chaves, H.E.; et al. Fifteen shades of green: The evolution of Bufotes toads revisited. Mol. Phylogenet. Evol. 2019, 141, 106615. [Google Scholar] [CrossRef] [PubMed]
- Bufotes viridis. Amphibian Species of the World 6.1, an Online Reference. American Museum of Natural History. Available online: https://amphibiansoftheworld.amnh.org/ (accessed on 22 January 2023).
- Ryzhikov, K.M.; Sharpilo, V.P.; Shevchenko, N.N. Helminths of Amphibians in the USSR Fauna; Nauka: Moscow, Russia, 1980; pp. 1–279. [Google Scholar]
- Baker, M.R. On three Oswaldocruzia spp. (Trichostrongyloidea: Molineidea) in amphibians from Africa. Can. J. Zool. 1981, 59, 246–251. [Google Scholar] [CrossRef]
- Durette-Desset, M.C.; Batcharov, A.; Ben Slimane, B.; Chabaud, A. Some Oswaldocruzia (Nematoda: Trichostrongyloidea) parasites of Amphibia in Bulgaria. Redescription of Oswaldocruzia bialata (Molin, 1860). Helminthologia 1993, 30, 99–104. [Google Scholar]
- Ben Slimane, B.; Durette-Desset, M.C. Quatre nouvelles espéces du genre Oswaldocruzia Travassos, 1917 (Nematoda: Trichostrongyloidea) parasites d’Amphibiens d’ Equateur. Rev. Suiss. Zool. 1993, 100, 113–136. [Google Scholar] [CrossRef]
- Ben Slimane, B.; Lluch, J.; Durette-Desset, M.C. Two new species of the genus Oswaldocruzia Travassos, 1917 (Nematode: Trichostrongylina: Molineidae) parasitizing Spanish amphibians. Res. Rev. Parasitol. 1995, 55, 209–215. [Google Scholar]
- Moravec, F.; Kaiser, H. Helminth parasites from West Indian frogs, with descriptions of two new species. Carib. J. Sci. 1995, 31, 252–268. [Google Scholar]
- Ben Slimane, B.; Durette-Desset, M.-C. Four new species of Oswaldocruzia (Nematoda: Trichostrongylina, Molineoidea) Parasitizing amphibians and lizards from Ecuador. Mem. Inst. Osw. Cruz 1996, 91, 317–328. [Google Scholar] [CrossRef]
- Ben Slimane, B.; Durette-Desset, M.-C. New Oswaldocruzia (Nematoda, Trichostrongylina, Molineoidea) parasites of amphibians from French Guyana and Ecuador. Misc. Zool. 1996, 19, 55–66. [Google Scholar]
- Ben Slimane, B.; Chabaud, A.G.; Durette-Desset, M.-C. Les Nématodes Trichostrongylina parasites d’Amphibiens et de Reptiles: Problèmes taxonomiques, phylétiques et biogéographiques. Syst. Parasitol. 1996, 35, 179–206. [Google Scholar] [CrossRef]
- Ben Slimane, B.; Guerrero, R.; Durette-Desset, M.C. Oswaldocruzia venezuelensis sp. n. (Nematoda: Trichostrongylina, Molineoidea), a parasite of Bufo marinus from Venezuela. Folia Parasitol. 1996, 43, 297–300. [Google Scholar]
- Vashetko, E.V.; Siddikov, H.B. The effect of the ecology of toads on the distributions of helmints. Turk. J. Zool. 1999, 23, 107–110. [Google Scholar]
- Yildirimhan, H.S. Researches on parasitic helminths of Bufo viridis Laurenti, 1768 (Anura: Amphibia). Turk. J. Zool. 1999, 23, 177–195. [Google Scholar]
- Kirillov, A.A. Helminth fauna of reptiles from Samara region. Proc. Sam. Sci. Cent. RAS 2000, 2, 324–329. [Google Scholar]
- Galli, P.; Crosa, G.; Gentilli, A.; Santagostino, M. New geographical records of parasitic nematodes from Bufo bufo in Italy. Parassitologia 2001, 43, 147–149. [Google Scholar] [PubMed]
- Bursey, C.R.; Goldberg, S.R. Cosmocerca vrcibradici n. sp. (Ascaridida: Cosmocercidae), Oswaldocruzia vitti n. sp. (Strongylida: Molineoidae), and other helminths from Prionodactylus eigenmanni and Prionodactylus oshaughnessyi (Sauria: Gymnophthalmidae) from Brazil and Ecuador. J. Parasitol. 2004, 90, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Bursey, C.R.; Goldberg, S.R. New species of Oswaldocruzia (Nematoda: Molineoidae), new species of Rhabdias (Nematoda: Rhabdiasidae), and other helminths in Rana cf. forreri (Anura: Ranidae) from Costa Rica. J. Parasitol. 2005, 91, 600–605. [Google Scholar] [CrossRef]
- Bursey, C.R.; Goldberg, S.R.; Vitt, L.J. New species of Oswaldocruzia (Nematoda: Molineidae) in Ameiva festiva (Squamata: Teiidae) from Nicaragua. J. Parasitol. 2006, 92, 350–352. [Google Scholar] [CrossRef]
- Durette-Desset, M.C.; Alves dos Anjos, L.; Vrcibradic, D. Three new species of the genus Oswaldocruzia Travassos, 1917 (Nematoda, Trichostrongylina, Molineoidea) parasites of Enyalius spp. (Iguanidae) from Brazil. Parasite 2006, 13, 115–125. [Google Scholar] [CrossRef]
- Bursey, C.R.; Goldberg, S.R.; Telford, S.R. Gastrointestinal helminths of 14 species of lizards from Panama with descriptions of five new species. Comp. Parasitol. 2007, 74, 108–140. [Google Scholar] [CrossRef]
- Murvanidze, L.; Nikolaishvili, K.; Lomidze, T. The annotated list of amphibians of Georgia. Proc. Inst. Zool. 2008, 23, 43–49. [Google Scholar]
- Santos, J.N.; Giese, E.G.; Maldonado, A.J.; Lanfredi, R.M. A new species of Oswaldocruzia (Molineidae: Nematoda) in Chaunus marinus (Amphibian: Bufonidae) (Linneaus, 1758) from Brazil. J. Parasitol. 2008, 94, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Schotthoefer, A.M.; Bolek, M.G.; Cole, R.; Beasley, V.R. Parasites of the Mink Frog (Rana septentrionalis) from Minnesota, U.S.A. Comp. Parasitol. 2009, 76, 240–246. [Google Scholar] [CrossRef]
- Düşen, S.; Oğuz, M.C. Metazoan endoparasites of three species of anurans collected from the Middle Black Sea Region of Turkey. Helminthologia 2010, 47, 226–232. [Google Scholar] [CrossRef]
- Dusen, S.; Oguz, M.C.; Barton, D.P.; Aral, A.; Sulekoglu, S.; Tepe, Y. Metazoan parasitological research on three species of anurans collected from Çanakkale Province, Northwestern Turkey. North-West. J. Zool. 2010, 6, 25–35. [Google Scholar]
- Mohammad, M.K.; Al-Moussawi, A.A.; Jasim, S.Y. Helminth parasites of the Green Toad Bufo viridis Laurenti, 1768 in Baghdad Area, Central Iraq. Egypt. Acad. J. Biol. Sci. 2010, 2, 17–25. [Google Scholar] [CrossRef]
- Bursey, C.R.; Goldberg, S.R. New species of Oswaldocruzia (Nematoda: Molineidae) and other helminths in Bolitoglossa subpalmata (Caudata: Plethodontidae) from Costa Rica. J. Parasitol. 2011, 97, 286–292. [Google Scholar] [CrossRef]
- Svitin, R.; Kuzmin, Y. Oswaldocruzia duboisi (Nematoda, Molineidae): Morphology, Hosts and Distribution in Ukraine. Vest. Zool. 2012, 46, 1–9. [Google Scholar] [CrossRef]
- Guerrero, R. Two new species of Oswaldocruzia (Nematoda: Trichostrongylina: Molineoidea) parasites of the cane toad Rhinella marina (Amphibia: Anura) from Peru. Acta Parasitol. 2013, 58, 30–36. [Google Scholar] [CrossRef]
- Svitin, R.S. New Data on the morphology and distribution of Oswaldocruzia skrjabini (Nematoda, Molineidae). Vest. Zool. 2015, 46, 195–203. [Google Scholar] [CrossRef]
- Herczeg, D.; Vörös, J.; Végvári, Z.; Kuzmin, Y.; Brooks, D.R. Helminth Parasites of the Pelophylax esculentus Complex (Anura: Ranidae) in Hortobágy National Park (Hungary). Comp. Parasitol. 2016, 83, 36–48. [Google Scholar] [CrossRef]
- Svitin, R. Nematodes of the genus Oswaldocruzia Travassos, 1917 of Western Palaearctic. Ph.D. Thesis, Institute of Zoology, Kyiv, Ukraine, 2016. [Google Scholar]
- Willkens, Y.; Maldonado, A.; dos Santos, J.N.; Maschio, G.F.; Melo, F.T.D.V. Redescription of Oswaldocruzia chambrieri (Strongylida: Molineidae) from Rhinella margaritifera (Anura: Bufonidae) in Caxiuanã National Forest, Brazil. Acta Parasitol. 2016, 61, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Kirillov, A.A.; Kirillova, N.Y. Overview of helminths in reptiles of the National Park «Samarskaya Luka» (Russia). Nat. Conserv. Res. 2018, 3 (Suppl. 1), 73–82. [Google Scholar] [CrossRef]
- Larrat, Y.M.; Melo, F.T.D.V.; Gomes, T.F.F.; Wilkens, Y.; dos Santos, J.N. Oswaldocruzia lanfrediae n. sp. (Strongylida: Molineidae), a parasite of Leptodactylus paraensis Heyer (Anura: Leptodactylidae) in Brazil. Syst. Parasitol. 2018, 95, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U.; Heneberg, P.; Těšínský, M.; Balczun, C.; Scheid, P. Helminth endoparasites of the smooth newt Lissotriton vulgaris: Linking morphological identification and molecular data. J. Helminthol. 2019, 93, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Andreev, V.Y. To the helminth fauna of the green toad (Bufo viridis Laur.). In Ecological and Biological Problems of the Caspian Sea Basin: Materials of VIII International Conference (11–12 October 2005); Pilipenko, V.N., Ed.; Astrakhan University: Astrakhan, Russia, 2005; pp. 3–5. [Google Scholar]
- Kalmykov, A.P.; Semenova, N.N.; Ivanov, V.M. Helminths in the Volga Delta Ecosystem. Volume 2. Nematodes; Print: Izhevsk, Russia, 2017; pp. 1–350. [Google Scholar]
- Kirillova, N.Y.; Kirillov, A.A.; Shchenkov, S.V.; Chikhlyaev, I.V. Oswaldocruzia filiformis sensu lato (Nematoda: Molineidae) from amphibians and reptiles of European Russia: Morphological and molecular data. Nat. Cons. Res. 2020, 5, 41–56. [Google Scholar] [CrossRef]
- Kirillova, N.Y.; Kirillov, A.A. Morphological variability of Oswaldocruzia filiformis (Nematoda: Molineidae) in reptiles inhabiting the protected areas of the Republic of Mordovia (Russia). IOP Conf. Ser. Earth Environ. Sci. 2020, 607, 012007. [Google Scholar] [CrossRef]
- Kirillova, N.Y.; Kirillov, A.A.; Chikhlyaev, I.V. Morphological variability of Oswaldocruzia filiformis (Nematoda: Molineidae) in amphibians from European Russia. IOP Conf. Ser. Earth Environ. Sci. 2021, 818, 012018. [Google Scholar] [CrossRef]
- Moravec, F.; Vojtkova, L. Variabilität von zwei Nematodenarten Oswaldocruzia filiformis (Goeze, 1782) und Oxysomatium brevicaudatum (Zeder, 1800). In Der Gemeinsamen Parasiten der Europäischen Amphibien und Reptilien. Scripta Fac. Sci. Natur. Univ. Purk. Brun. Biol. 1975, 2, 61–76. [Google Scholar]
- Shaldybin, S.L. To the parasite fauna of anurans in the Volga-Kama reserve. In Issues of Herpetology; Darevsky, I.S., Ed.; Nauka: Leningrad, Russia, 1977; pp. 228–230. [Google Scholar]
- Smirnova, M.I.; Gorshkov, P.K.; Sizova, V.G. Helminth Fauna of Tailless Amphibians in Tatarstan Republic; Institute of Biology, Kazan Branch of Academy of Sciences of the USSR: Kazan, Russia, 1987; pp. 3–19. [Google Scholar]
- Ayupov, K.V.; Valiullin, S.M.; Khaziev, G.Z.; Bayanov, M.G.; Kazadaev, V.I.; Antonov, N.P. Helminths of animals, humans and plants in the Bashkirian ASSR. In Helminths of Animals, Humans and Plants in the Southern Urals; Bayanov, M.G., Ed.; Bashkiria Branch of Academy of Sciences of the USSR: Ufa, Russia, 1974; pp. 8–29. [Google Scholar]
- Bayanov, M.G.; Yumagulova, G.R. Helminths in anurans from various habitats. Res. Biol. Res. 2000, 6, 153–155. [Google Scholar]
- Petrova, S.V.; Bayanov, M.G. Helminths in toads (Amphibia, Bufonidae) in Bashkiria. Res. Biol. Res. 2000, 6, 155–157. [Google Scholar]
- Yumagulova, G.R. Helminths of amphibians in the Southern Urals. Ph.D. Thesis, Bashkirian State University, Ufa, Russia, 2000. [Google Scholar]
- Ravkovskaya, E.A.; Polyakova, N.A.; Terekhina, M.S.; Pyatova, M.V.; Lada, G.A. The first data about the helminths in the green toad Bufotes viridis (Laurenti, 1768) in the Tambov region. In Recent Problems of Parasitology and Ecology: Readings Dedicated to the Memory of S.S. Shulman; Kirillov, A.A., Ed.; Anna: Togliatti, Russia, 2018; pp. 223–228. [Google Scholar]
- Vekhnik, V.P.; Golovatyuk, L.V.; Goreslavets, I.N.; Dyuzhaeva, I.V.; Zharikov, V.V.; Zinchenko, T.D.; Kirillov, A.A.; Kirillova, N.Y.; Krasnobaev, Y.P.; Krasnobaeva, T.P.; et al. Cadastre of Invertebtates of the Samarskaya Luka; Ofort: Samara, Russia, 2007; pp. 1–471. [Google Scholar]
- Fayzulin, A.I.; Chikhlyaev, I.V.; Kuzovenko, A.E. Amphibians of the Samara Region; Kassandra: Togliatti, Russia, 2013; pp. 1–140. [Google Scholar]
- Chikhlyaev, I.V.; Fayzulin, A.I.; Kuzovenko, A.E. The analysis of helminthes fauna of a green toad Bufotes viridis (Laurenti, 1768) on the urbanized territories of the Samara region. Proc. Sam. Sci Cent. RAS 2017, 19, 178–184. [Google Scholar]
- Chikhlyaev, I.V.; Kirillova, N.Y.; Kirillov, A.A. Overview of helminths of amphibians (Amphibia) from Samara region. Proc. Sam. Sci. Cent. RAS 2018, 20, 385–400. [Google Scholar]
- Kirillov, A.A.; Kirillova, N.Y.; Chikhlyaev, I.V. Parasites of Vertebrates of the Samara Region; Polyar: Togliatti, Russia, 2018; pp. 1–304. [Google Scholar]
- Kostyunin, V.M. Helminth Fauna of Land Vertebrates in the Middle Volga Region; Nizhny Novgorod State Pedagogical University: Nizhny Novgorod, Russia, 2010; pp. 1–225. [Google Scholar]
- Davletbakova, G.M.; Yumagulova, G.R. Helminths of tailless amphibians in the Orenburg region. In Ecological Collection Papers: Proceedings of Young Scientists of the Volga Region; Saxonov, S.V., Ed.; Cassandra: Togliatti, Russia, 2013; pp. 31–34. [Google Scholar]
- Burakova, A.V.; Vershinin, V.L. Analysis of the parasite fauna of syntopically inhabiting representatives of anurans. Vest. St. Petersb. Univ. Ser. 3. Biol. 2016, 3, 31–36. [Google Scholar] [CrossRef]
- Shimalov, V.V.; Shimalov, V.T. Helminth fauna of toads in Belorussian Polesie. Parasitol. Res. 2001, 87, 84. [Google Scholar] [CrossRef] [PubMed]
- Shimalov, V.V. Helminth fauna of amphibians (Vertebrata: Amphibia) in the Republic of Belarus. Parazitologiia 2009, 43, 118–129. [Google Scholar]
- Buchvarov, G.K. Catalog of Helminths of Amphibians in Bulgaria; Plovdiv University, P. Hilendarski: Plovdiv, Bulgaria, 1977; pp. 1–53. [Google Scholar]
- Schad, G.A.; Kuntz, R.E.; Wells, W.H. Nematode Parasites from Turkish Vertebrates: An Annotated List. Can. J. Zool. 1960, 38, 949–963. [Google Scholar] [CrossRef]
- Ikromov, E.F. Some patterns of infection of amphibians with helminths depending on age. World Sci. Discov. 2010, 9, 33–36. [Google Scholar]
- Ivanitsky, S.V. Materials for the helminth fauna of Ukrainian vertebrates (fauna of cestodes, nematodes and acanthocephalans). Proc. Kharkov Vet. Inst. 1940, 19, 129–155. [Google Scholar]
- Sudarikov, V.E. Helminth fauna of vertebrates in the Middle Volga region. Proc. Helminthol. Lab. Acad. Sci. USSR 1951, 5, 326–330. [Google Scholar]
- Kozak, A. Helminth fauna of frogs from the surroundings of Kosice. Biologia 1966, 21, 606–611. [Google Scholar]
- Chikhlyaev, I.V.; Ruchin, A.B.; Fayzulin, A.I. The helminth fauna study of European common toad in the Volga Basin. Nat. Environ. Poll. Tech. 2016, 15, 1103–1109. [Google Scholar]
- Chikhlyaev, I.V.; Ruchin, A.B. An Overview of the Helminths of Moor Frog Rana arvalis Nilsson, 1842 (Amphibia: Anura) in the Volga Basin. Diversity 2021, 13, 61. [Google Scholar] [CrossRef]
- Chikhlyaev, I.V.; Ruchin, A.B. Ecological Analysis and Biodiversity of the Helminth Community of the Pool Frog Pelophylax lessonae (Amphibia: Anura) from Floodplain and Forest Water Bodies. Diversity 2022, 14, 247. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, G.-H.; Zhao, G.-H.; Cai, J.-Z.; Zhu, X.-Q.; Qian, A.-D. Genetic differences between Chabertia ovina and C. erschowirevealed by sequence analysis of four mitochondrial genes. Mitochondrial DNA 2013, 26, 167–170. [Google Scholar] [CrossRef]
- Hu, W.; Yu, X.; Wu, S.; Tan, L.; Song, M.; Abdulahi, A.; Wang, Z.; Jiang, B.; Li, G. Levels of Ancylostoma infections and phylogenetic analysis of cox 1 gene of A. ceylanicum in stray cat faecal samples from Guangzhou, China. J. Helminthol. 2016, 90, 392–397. [Google Scholar] [CrossRef]
- Durette-Desset, M.-C. Trichostrongyloid Nematodes and their Vertebrate Hosts: Reconstruction of the Phylogeny of a Parasitic Group. Adv. Parasitol. 1985, 24, 239–306. [Google Scholar] [CrossRef]
- Durette-Desset, M.-C.; Chabaud, A.G. Nouvel essai de classification des Nématodes Trichostrongyloidea. Ann. Parasitol. Hum. Comp. 1981, 56, 297–312. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; McManus, D. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.r-project.org/index.html (accessed on 22 January 2023).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 45–52. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Gu, Z. Complex heatmap visualization. Imeta 2022, 1, 1–15. [Google Scholar] [CrossRef]
- Stöck, M.; Roth, P.; Podloucky, R.; Grossenbacher, K. Wechselkröten unter Berücksichtigung von Bufo viridis viridis Laurenti, 1768; Bufo variabilis (Pallas, 1769); Bufo boulengeri Lataste, 1879; Bufo balearicus Böttger, 1880 und Bufo siculus Stöck, Sicilia, Belfiore, Lo Brutto, Lo Valvo und Arculeo, 2008. In Handbuch der Reptilien und Amphibien Europas, Band 5/II, Froschlurche (Anura) II (Hylidae, Bufonidae); Grossenbacher, K., Ed.; AULA-Verlag: Wiebelsheim, Germany, 2009; pp. 413–498. [Google Scholar]
- Kovacs, E.-H.; Sas, I. Aspects of breeding activity of Bufo viridis in an urban habitat: A case study in Oradea, Romania. Biharean Biol. 2010, 4, 73–77. [Google Scholar]
- Kuzmin, S.L. Amphibians of the Former USSR, 2nd ed.; KMK: Moscow, Russia, 2012; pp. 1–370. [Google Scholar]
- Kaczmarski, M.; Szala, K.; Kloskowski, J. Early onset of breeding season in the green toad Bufotes viridis in Western Poland. Herpetozoa 2019, 32, 109–112. [Google Scholar] [CrossRef]
- Konowalik, A.; Najbar, A.; Konowalik, K.; Dylewski, Ł.; Frydlewicz, M.; Kisiel, P.; Starzecka, A.; Zaleśna, A.; Kolenda, K. Amphibians in an urban environment: A case study from a central European city (Wrocław, Poland). Urban Ecosyst. 2019, 23, 235–243. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nematode Species, Isolate No. | Locality | Coordinates | Host | GenBank Acc. No. | Museum and Laboratory Specimen No. | Source |
|---|---|---|---|---|---|---|
| Oswaldocruzia ukrainae, 599, 600 | Nikolaevka village, suburb of Saransk, Republic of Mordovia | 54°8′34″ N, 45°8′55″ E | Bufotes viridis | OQ351737 OQ351738 | Nem-Os333-599, Nem-Os-334-600 | This study |
| Oswaldocruzia ukrainae, 597, 598 | Lykovshchina village, Republic of Mordovia | 54°18′45″ N, 45°29′13″ E | Bufotes viridis | OQ351735 OQ351736 | Nem-Os-331-597, Nem-Os-332-598 | This study |
| Oswaldocruzia ukrainae, 595, 601 | Samara city | 53°12′27″ N, 50°9′18″ E | Bufotes viridis | OQ351734 OQ351739 | Nem-Os-329-595, Nem-Os-335-601 | This study |
| Oswaldocruzia filiformis, 594 | Vicinity of Spassk city, Penza region | 53°54′49″ N, 43°12′36″ E | Bufotes viridis | OQ346357 | Nem-Os-328-594 | This study |
| Oswaldocruzia filiformis, 116 | Samarskaya Luka National Park, Samara region | 53°10′34″ N, 49°26′12″ E | Pelophylax ridibundus | MT300271 | Nem-Os-116 | [40] |
| Oswaldocruzia filiformis, 120 | Samarskaya Luka National Park, Samara region | 53°10′34″ N, 49°26′12″ E | Pelophylax ridibundus | MT300269 | Nem-Os-120 | [40] |
| Oswaldocruzia filiformis, 183 | Samarskaya Luka National Park, Samara region | 53°10′34″ N, 49°26′12″ E | Pelophylax ridibundus | MT300272 | Nem-Os-183 | [40] |
| Oswaldocruzia filiformis, 155 | Smolny National Park, Republic of Mordovia | 55°50′07″ N, 45°22′43″ E | Bufo bufo | MT300259 | Nem-Os-155 | [40] |
| Oswaldocruzia filiformis, 156 | Smolny National Park, Republic of Mordovia | 55°45′36″ N, 45°24′21″ E | Rana arvalis | MT300268 | Nem-Os-156 | [40] |
| Oswaldocruzia filiformis, 159 | Zvenigorod Biol. Station of Moscow University, Moscow region | 55°42′02″ N, 36°43′20″ E | Rana temporaria | MT300258 | Nem-Os-159 | [40] |
| Oswaldocruzia filiformis, 157 | Uzola floodplain, Nizhny Novgorod region | 56°36′12″ N, 43°35′06″ E | Rana arvalis | MT300257 | Nem-Os-157 | [40] |
| Oswaldocruzia filiformis, 158 | Ural floodplain, Orenburg region | 51°13′05″ N, 58°33′22″ E | Rana arvalis | MT300256 | Nem-Os-158 | [40] |
| Oswaldocruzia filiformis, 16 | Mordovia Nature Reserve, Republic of Mordovia | 54°42′49″ N, 43°13′39″ E | Lacerta agilis | MT300266 | Nem-Os-16 | [40] |
| Oswaldocruzia filiformis, 8 | Mordovia Nature Reserve, Republic of Mordovia | 54°42′49″ N, 43°13′39″ E | Lacerta agilis | MT300262 | Nem-Os-18 | [40] |
| Oswaldocruzia filiformis, 10 | Mordovia Nature Reserve, Republic of Mordovia | 54°42′49″ N, 43°13′39″ E | Zootoca vivipara | MT300260 | Nem-Os-10 | [40] |
| Oswaldocruzia filiformis, 16_1 | Mordovia Nature Reserve, Republic of Mordovia | 54°42′49″ N, 43°13′39″ E | Zootoca vivipara | MT300273 | Nem-Os-16_1 | [40] |
| Oswaldocruzia filiformis, 88_4 | Smolny National Park, Republic of Mordovia | 54°44′49″ N, 45°15′46″ E | Zootoca vivipara | MT300267 | Nem-Os-88_4 | [40] |
| Oswaldocruzia filiformis, 107_17 | Smolny National Park, Republic of Mordovia | 54°43′58″ N, 45°16′16″ E | Lacerta agilis | MT300265 | Nem-Os-107_17 | [40] |
| Oswaldocruzia filiformis, 95_5 | Smolny National Park, Republic of Mordovia | 54°43′58″ N, 45°16′16″ E | Lacerta agilis | MT300270 | Nem-Os-95_5 | [40] |
| Oswaldocruzia filiformis, 89_4 | Smolny National Park, Republic of Mordovia | 54°44′49″ N, 45°15′46″ E | Lacerta agilis | MT300275 | Nem-Os-89_4 | [40] |
| Oswaldocruzia filiformis, 190_1 | Smolny National Park, Republic of Mordovia | 54°44′41″ N, 45°30′08″ E | Anguis fragilis | MT300264 | Nem-Os-190_1 | [40] |
| Oswaldocruzia filiformis, 76_1 | Smolny National Park, Republic of Mordovia | 54°44′41″ N, 45°30′08″ E | Natrix natrix | MT300261 | Nem-Os-76_1 | [40] |
| Oswaldocruzia filiformis, 113 | Smolny National Park, Republic of Mordovia | 54°44′49″ N, 45°15′46″ E | Vipera berus | MT300263 | Nem-Os-113 | [40] |
| Oswaldocruzia filiformis, 123_4 | Smolny National Park, Republic of Mordovia | 54°44′49″ N, 45°15′46″ E | Vipera berus | MT300274 | Nem-Os-123_4 | [40] |
| Chabertia ovina | Jingyang County, China 1 | 34°31′45″ N, 108°50′24″ E | Capra aegagrus hircus | KF279335 | – | [70] |
| Ancylostoma ceylanicum | Guangzhou, Guangdong, China 1 | 23°07′49″ N, 113°15′33″ E | Felis catus (faeces) | KP072073 | – | [71] |
| Ancylostoma ceylanicum | Guangzhou, Guangdong, China 1 | 23°07′49″ N, 113°15′33″ E | Felis catus (faeces) | KP072079 | – | [71] |
| Characteristics | Togliatti (This Study) | Oktyabrskiy (This Study) | Samara (This Study) | Nikolaevka (This Study) | Lykovshchina (This Study) | Ivanitsky [64] | Svitin [32] | Ryzhikov et al. [3] |
|---|---|---|---|---|---|---|---|---|
| n 1 | 7 | 8 | 3 | 6 | 1 | 1 | 21 | – |
| Body length | 6.20–7.10 (6.71) | 6.35–7.83 (7.00) | 5.00–5.80 (5.35) | 5.20–5.94 (5.58) | 5.00 | 3.94 | 3.53–5.59 (4.79) | 3.8–5.0 |
| Body width | 0.110–0.130 (0.119) | 0.126–0.160 (0.144) | 0.108–0.123 (0.115) | 0.111–0.126 (0.118) | 0.104 | 0.140 | 0.080–0.170 (0.118) | 0.088–0.099 |
| Length of cephalic vesicle | 0.077–0.085 (0.081) | 0.081–0.089 (0.085) | 0.079–0.094 (0.087) | 0.095–0.111 (0.104) | 0.083 | 0.100 | 0.075–0.108 (0.091) | 0.088–0.099 |
| Width of cephalic vesicle | 0.035–0.039 (0.036) | 0.038–0.043 (0.040) | 0.036–0.040 (0.038) | 0.051–0.059 (0.056) | 0.041 | 0.041 | 0.038–0.048 (0.040) | 0.038–0.044 |
| Length of esophagus | 0.341–0.377 (0.362) | 0.362–0.446 (0.401) | 0.346–0.360 (0.353) | 0.422–0.454 (0.441) | 0.344 | 0.444 | 0.358–0.415 (0.380) | 0.360–0.390 |
| Width of esophageal bulb | 0.045–0.051 (0.048) | 0.051–0.059 (0.055) | 0.037–0.041 (0.039) | 0.039–0.052 (0.045) | 0.040 | – | 0.035–0.055 (0.047) | – |
| Distance from anterior end of body to nerve ring | 0.150–0.173 (0.160) | 0.160–0.183 (0.173) | 0.165–0.177 (0.172) | 0.230–0.250 (0.240) | 0.169 | – | 0.160–0.220 (0.192) | – |
| Distance from anterior end of body to excretory pore | 0.255–0.324 (0.309) | 0.320–0.389 (0.349) | 0.287–0.310 (0.297) | 0.382–0.397 (0.394) | 0.330 | – | 0.250–0.395 (0.346) | – |
| Tail length | 0.102–0.112 (0.106) | 0.106–0.118 (0.111) | 0.087–0.098 (0.092) | 0.092–0.100 (0.097) | 0.089 | – | 0.073–0.113 (0.086) | – |
| Length of spicules | 0.155.167 (0.160) | 0.158.170 (0.163) | 0.156–0.163 (0.160) | 0.148–0.158 (0.154) | 0.152, 0.154 | 0.141–0.160 | 0.135–0.168 (0.150) | 0.143–0.159 |
| Number of crests at mid-body level | 33–36 | 34–37 | 32–34 | 33–35 | 32 | 22–30 | 30 | – |
| Characteristics | Togliatti (This Study) | Oktyabrskiy (This Study) | Samara (This Study) | Nikolaevka (This Study) | Lykovshchina (This Study) | Svitin [32] | Ryzhikov et al. [3] |
|---|---|---|---|---|---|---|---|
| n | 9 | 11 | 5 | 6 | 2 | 29 | – |
| Body length | 9.95–11.05 (10.47) | 10.05–11.55 (10.81) | 9.25–10.00 (9.66) | 9.25–10.95 (10.11) | 9.95–10.48 (10.22) | 4.92–12.41 (7.74) | 8.00–9.20 |
| Body width | 0.161–0.209 (0.183) | 0.181–0.217 (0.201) | 0.163–0.189 (0.177) | 0.184–0.200 (0.192) | 0.186–0.198 (0.192) | 0.090–0.240 (0.173) | 0.130–0.150 |
| Length of cephalic vesicle | 0.083–0.102 (0.090) | 0.085–0.091 (0.087) | 0.083–0.108 (0.092) | 0.104–0.113 (0.108) | 0.110–0.117 (0.114) | 0.070–0.110 (0.091) | 0.088–0.100 |
| Width of cephalic vesicle | 0.039–0.043 (0.041) | 0.041–0.047 (0.044) | 0.040–0.044 (0.042) | 0.059–0.064 (0.061) | 0.056–0.061 (0.059) | 0.040–0.053 (0.046) | 0.044–0.049 |
| Length of esophagus | 0.352–0.413 (0.393) | 0.359–0.445 (0.400) | 0.350–0.407 (0.374) | 0.451–0.476 (0.463) | 0.413–0.444 (0.429) | 0.355–0.440 (0.395) | 0.400–0.450 |
| Width of esophageal bulb | 0.051–0.061 (0.056) | 0.059–0.067 (0.063) | 0.046–0.052 (0.049) | 0.045–0.065 (0.053) | 0.048–0.52 (0.050) | 0.045–0.063 (0.053) | – |
| Distance from anterior end of body to excretory pore | 0.177–0.205 (0.190) | 0.158–0.198 (0.179) | 0.170–0.200 (0.184) | 0.234–0.256 (0.248) | 0.227–0.242 (0.235) | 0.143–0.238 (0.191) | – |
| Distance from anterior end of body to vulva | 0.320–0.366 (0.348) | 0.295–0.369 (0.335) | 0.331–0.379 (0.351) | 0.420–0.458 (0.444) | 0.389–0.422 (0.406) | 0.265–0.450 (0.350) | – |
| Distance from anterior end of body to vulva | 6.00–6.75 (6.33) | 6.05–7.80 (6.79) | 6.17–6.54 (6.38) | 6.50–7.10 (6.80) | 6.13–6.38 (6.26) | 2,87–7.29 (4.61) | – |
| Tail length | 0.172–0.197 (0.185) | 0.189–0.217 (0.201) | 0.148–0.170 (0.158) | 0.165–0.209 (0.191) | 0.172–0.180 (0.176) | 0.128–0.238 (0.173) | 0.140–0.170 |
| Length of eggs | 0.081–0.089 (0.085) | 0.083–0.091 (0.087) | 0.085–0.095 (0.089) | 0.085–0.108 (0.096) | 0.085–0.098 (0.090) | 0.085–0.090 | 0.099–0.104 |
| Width of eggs | 0.045–0.050 (0.047) | 0.047–0.051 (0.049) | 0.047–0.053 (0.049) | 0.050–0.058 (0.054) | 0.047–0.055 (0.051) | 0.050–0.055 | |
| Number of crests at level of mid-body | 42–47 | 43–48 | 40–44 | 40–46 | 42–46 | 30 | – |
| Characters | Spassk (This Study) | Kirillova et al. [42] | Ivanitsky [64] | Ryzhikov et al. [3] | Kirillova et al. [42] | Ryzhikov et al. [3] |
|---|---|---|---|---|---|---|
| males | females | |||||
| n | 1 | 14 | 2 | |||
| Body length | 8.65 | 6.70–9.25 (7.92) | 4.00–7.32 | 5.9–6.2 | 16.55–17.20 (16.88) | 12.0–20.0 |
| Body width | 0.163 | 0.130–0.181 (0.149) | 0.125–0.187 | 0.130–0.160 | 0.240–0.256 (0.248) | 0.230–0.280 |
| Length of cephalic vesicle | 0.083 | 0.075–0.083 (0.079) | 0.084 | 0.077–0.082 | 0.094–0.098 (0.096) | 0.077–0.082 |
| Width of cephalic vesicle | 0.041 | 0.035–0.039 (0.037) | 0.050 | 0.049 | 0.047–0.051 (0.049) | 0.049–0.055 |
| Length of esophagus | 0.425 | 0.378–0.488 (0.431) | – | 0.380–0.420 | 0.559–0.587 (0.573) | 0.450–0.490 |
| Width of esophageal bulb | 0.051 | – | – | – | – | – |
| Distance from anterior end of body to nerve ring | 0.196 | – | – | – | – | – |
| Distance from anterior end of body to excretory pore | 0.364 | – | – | – | – | – |
| Distance from anterior end of body to vulva | – | – | – | – | 11.06–11.30 (11.18) | – |
| Tail length | 0.139 | 0.126–0.146 (0.134) | – | – | 0.276–0.297 (0.287) | 0.260–0.330 |
| Length of spicules | 0.228–0.230 | 0.192–0.236 (0.215) | 0.230 | 0.193–0.321 | – | – |
| Length of eggs | – | – | – | – | 0.079–0.087 (0.084) | 0.099–0.104 |
| Width of eggs | – | – | – | – | 0.039–0.047 (0.044) | 0.055 |
| Number of crests at level of mid-body | 40 | 33–40 | 30–50 | – | 49–50 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirillova, N.Y.; Kirillov, A.A.; Shchenkov, S.V.; Chikhlyaev, I.V. Oswaldocruzia ukrainae (Nematoda: Molineidae)—A Parasite of European Green Toad Bufotes viridis: Morphological and Molecular Data. Biology 2023, 12, 772. https://doi.org/10.3390/biology12060772
Kirillova NY, Kirillov AA, Shchenkov SV, Chikhlyaev IV. Oswaldocruzia ukrainae (Nematoda: Molineidae)—A Parasite of European Green Toad Bufotes viridis: Morphological and Molecular Data. Biology. 2023; 12(6):772. https://doi.org/10.3390/biology12060772
Chicago/Turabian StyleKirillova, Nadezhda Yu., Alexander A. Kirillov, Sergei V. Shchenkov, and Igor V. Chikhlyaev. 2023. "Oswaldocruzia ukrainae (Nematoda: Molineidae)—A Parasite of European Green Toad Bufotes viridis: Morphological and Molecular Data" Biology 12, no. 6: 772. https://doi.org/10.3390/biology12060772