
SerpinB3 Upregulates Low-Density Lipoprotein Receptor-Related Protein (LRP) Family Members, Leading to Wnt Signaling Activation and Increased Cell Survival and Invasiveness
 , ,
, ,  , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.1.1. Primary Human Monocytes
2.1.2. THP-1 Cell Line
2.1.3. HepG2 Cell Line
2.2. Cell Proliferation Assays
2.3. Cell Invasion Assay
2.4. SerpinB3 Quantification by ELISA
2.5. Immunofluorescence
2.5.1. Primary Human Monocytes and THP-1 Cells
2.5.2. HepG2 Cells
2.6. ImageStream Analysis
2.7. RNA Isolation and Quantitative Real-Time PCR
2.8. Western Blot Analysis
2.9. Mouse Model of Liver Carcinogenesis
2.10. Immunohistochemistry
2.11. Patients
2.12. Amino Acid and Structure Alignment
2.13. Statistical Analysis
3. Results
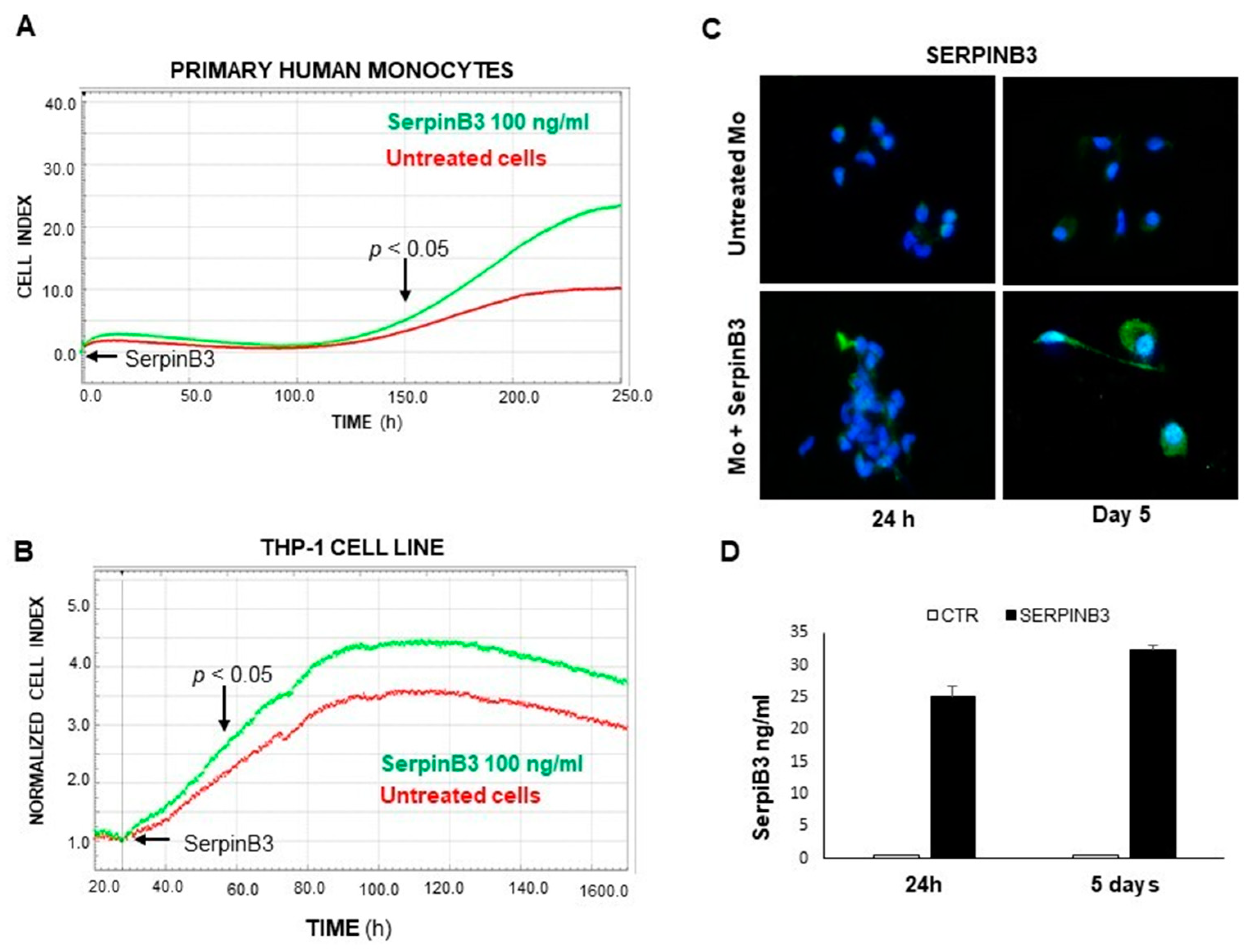
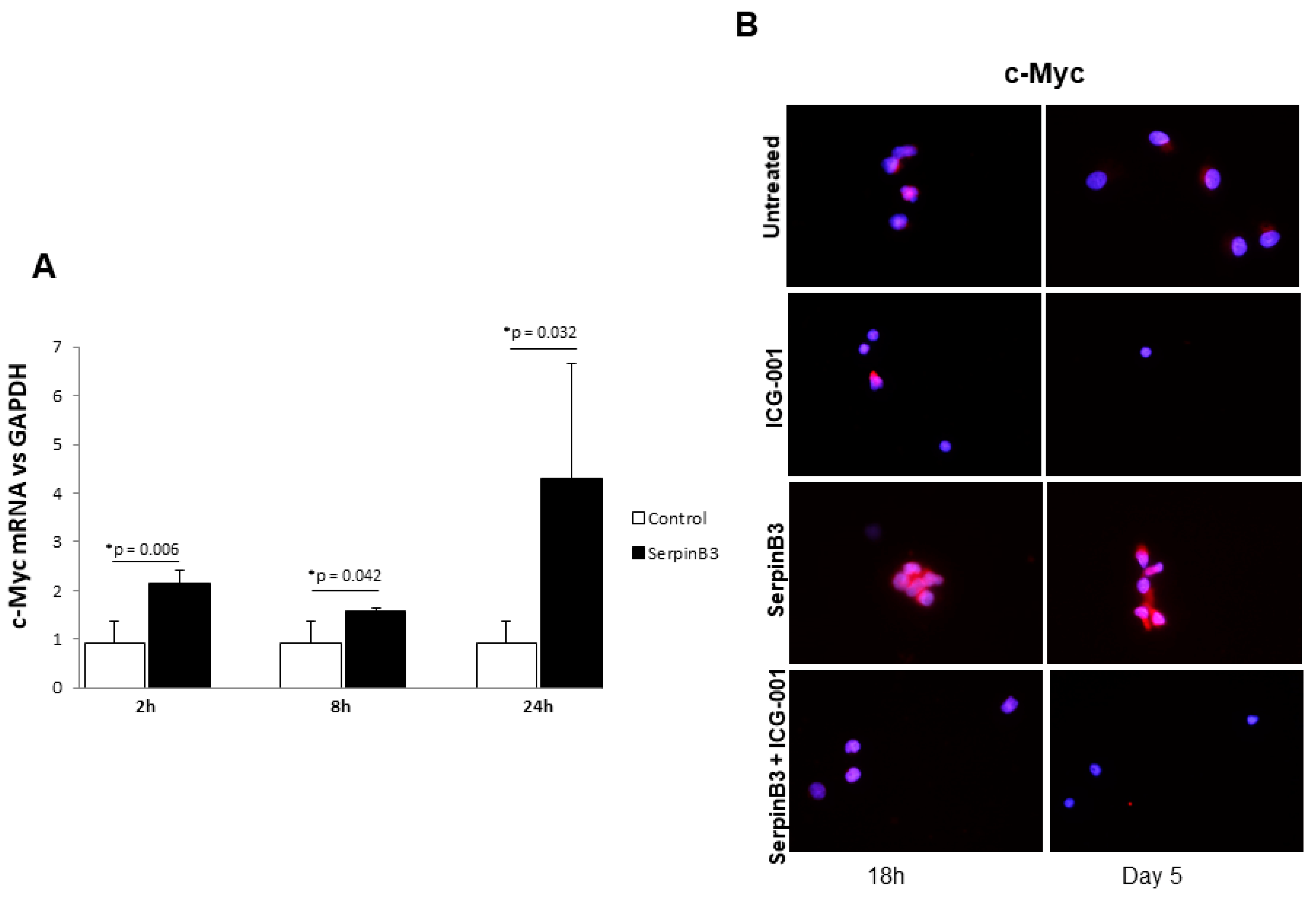
3.1. Exogenous SerpinB3 Increases Monocyte Proliferation and Induces Its Endogenous Expression
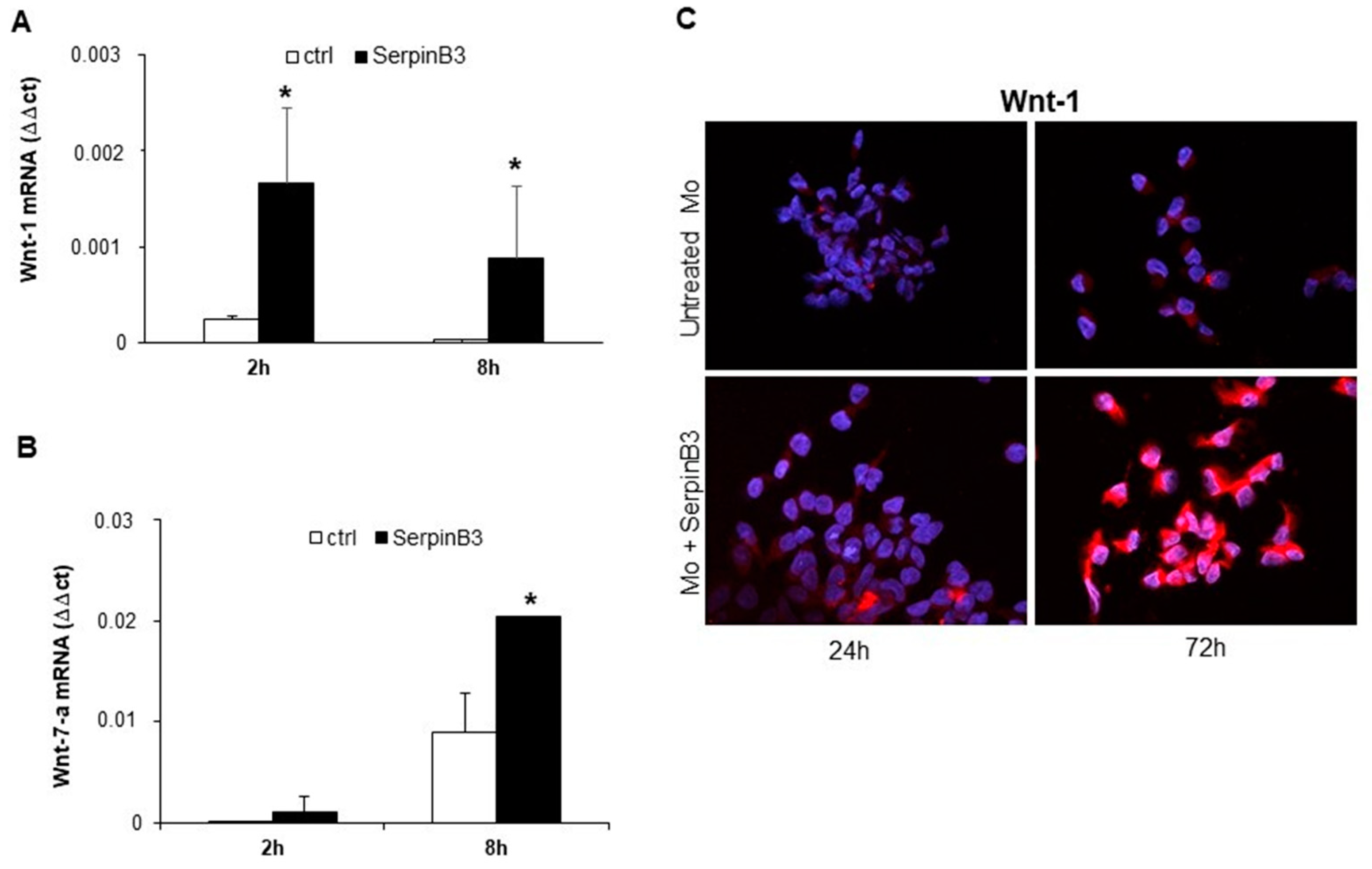
3.2. SerpinB3 Induces Wnt-1 and Wnt-7a
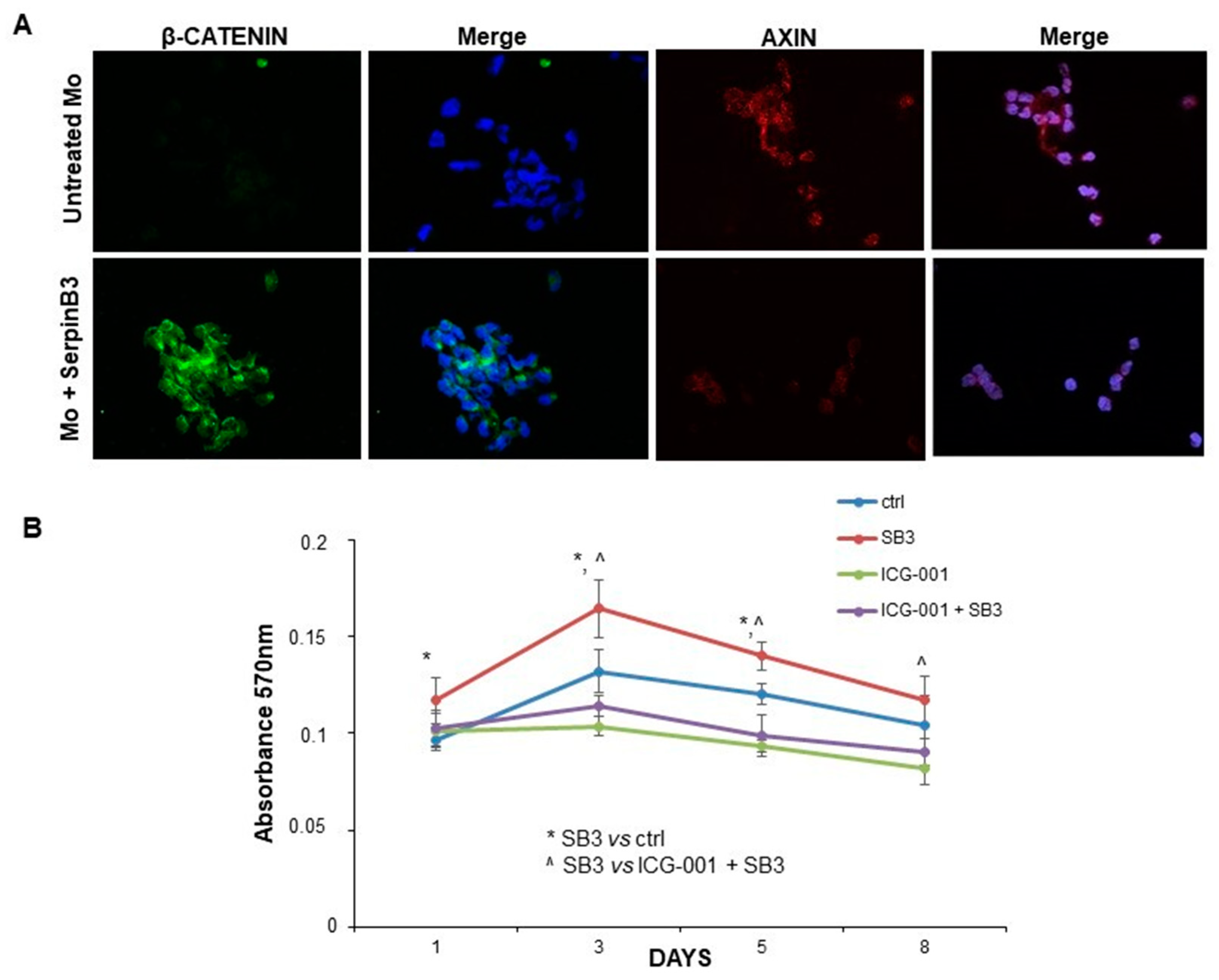
3.3. SerpinB3 Activates the Wnt-Canonical Pathway
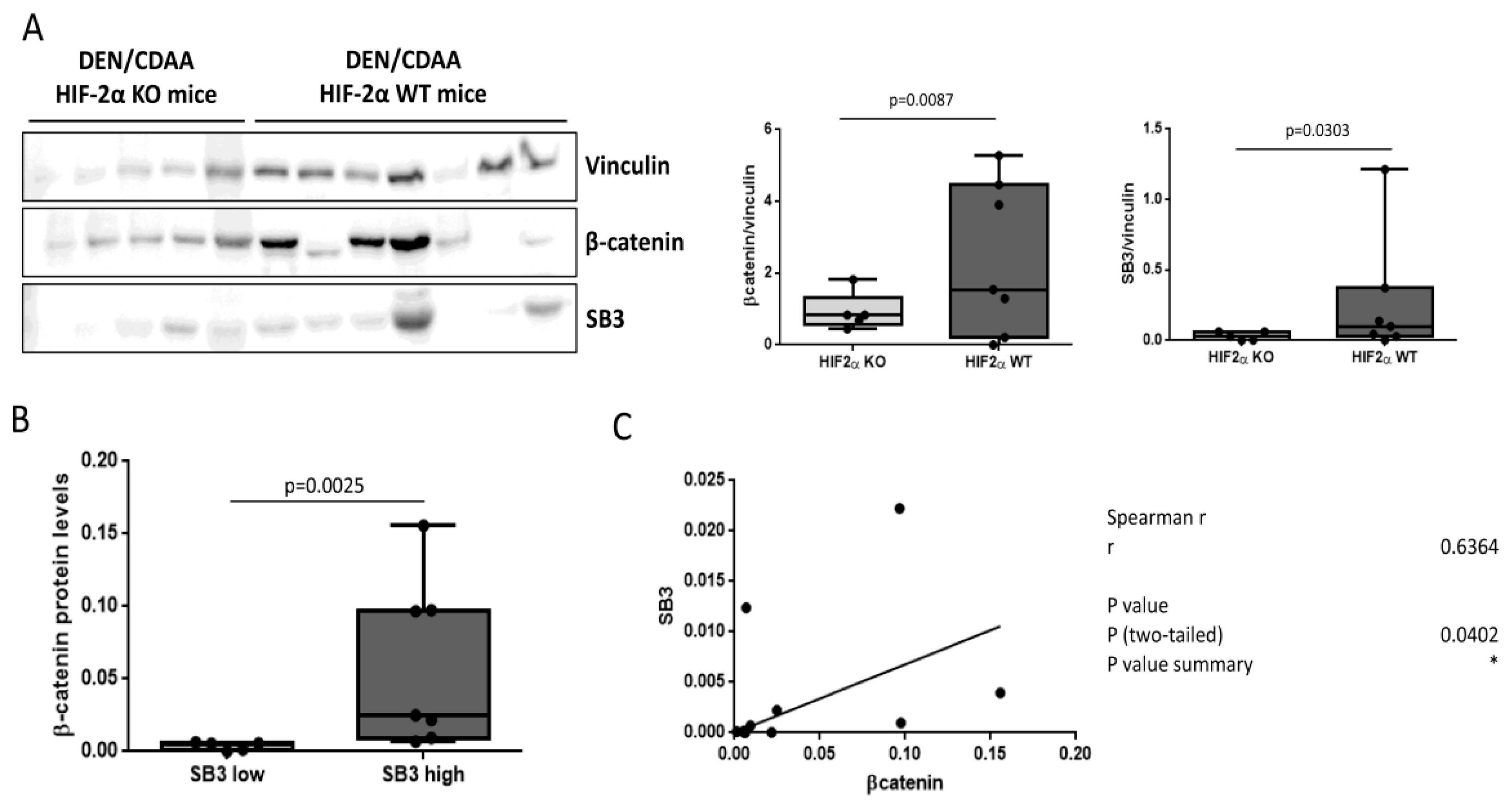
3.4. SerpinB3 and Wnt/β-Catenin Axis in an Experimental Model of Liver Carcinogenesis
3.5. SerpinB3 and Wnt Family Members in Human Liver Tumors
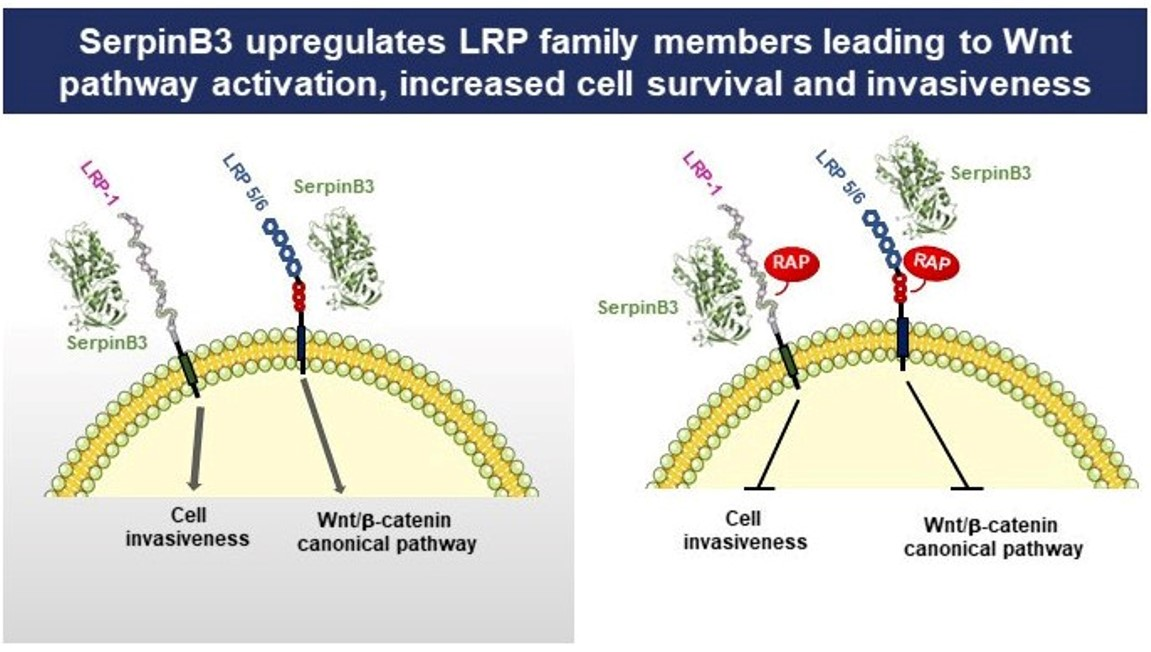
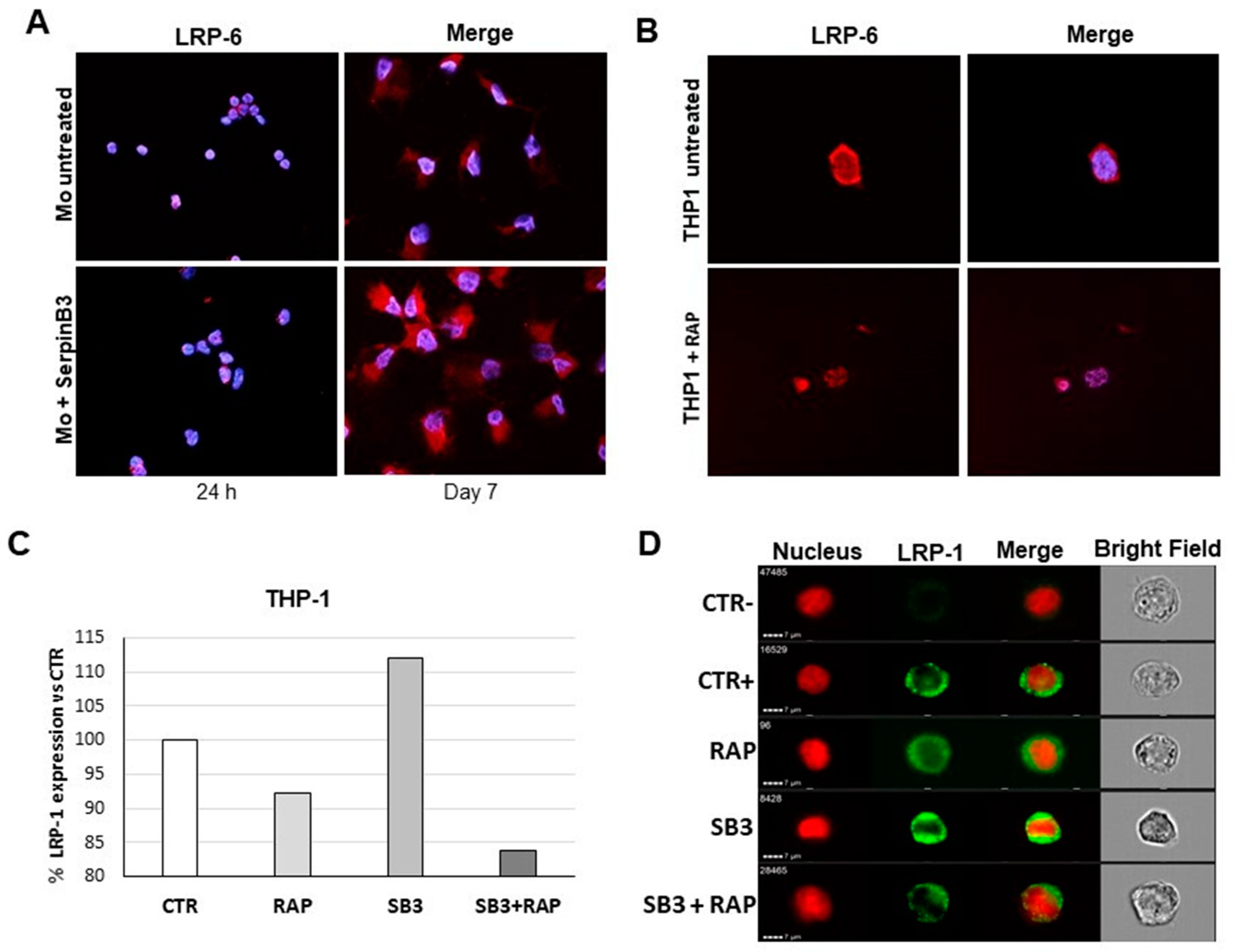
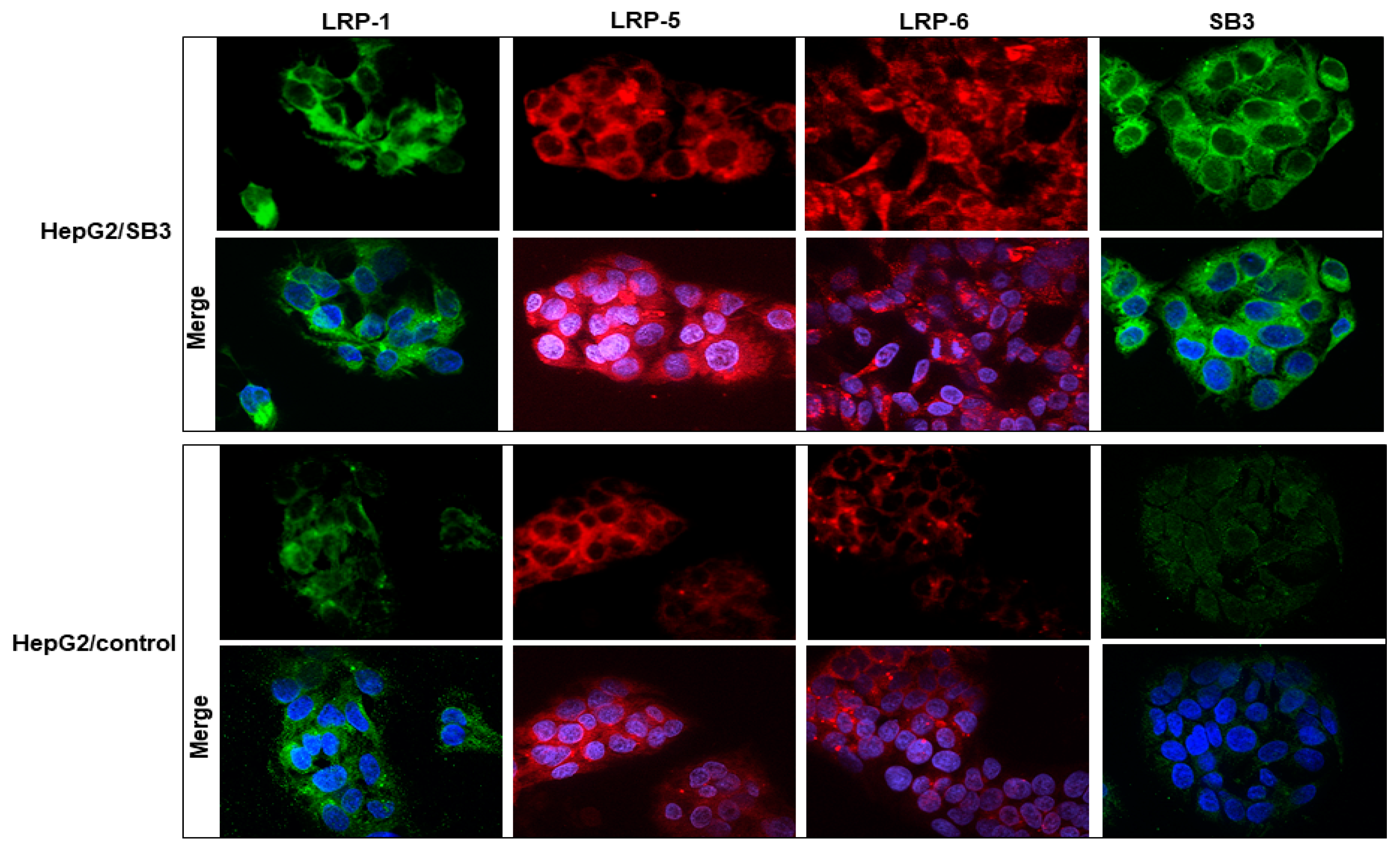
3.6. SerpinB3 Upregulates Low-Density Lipoprotein Receptors
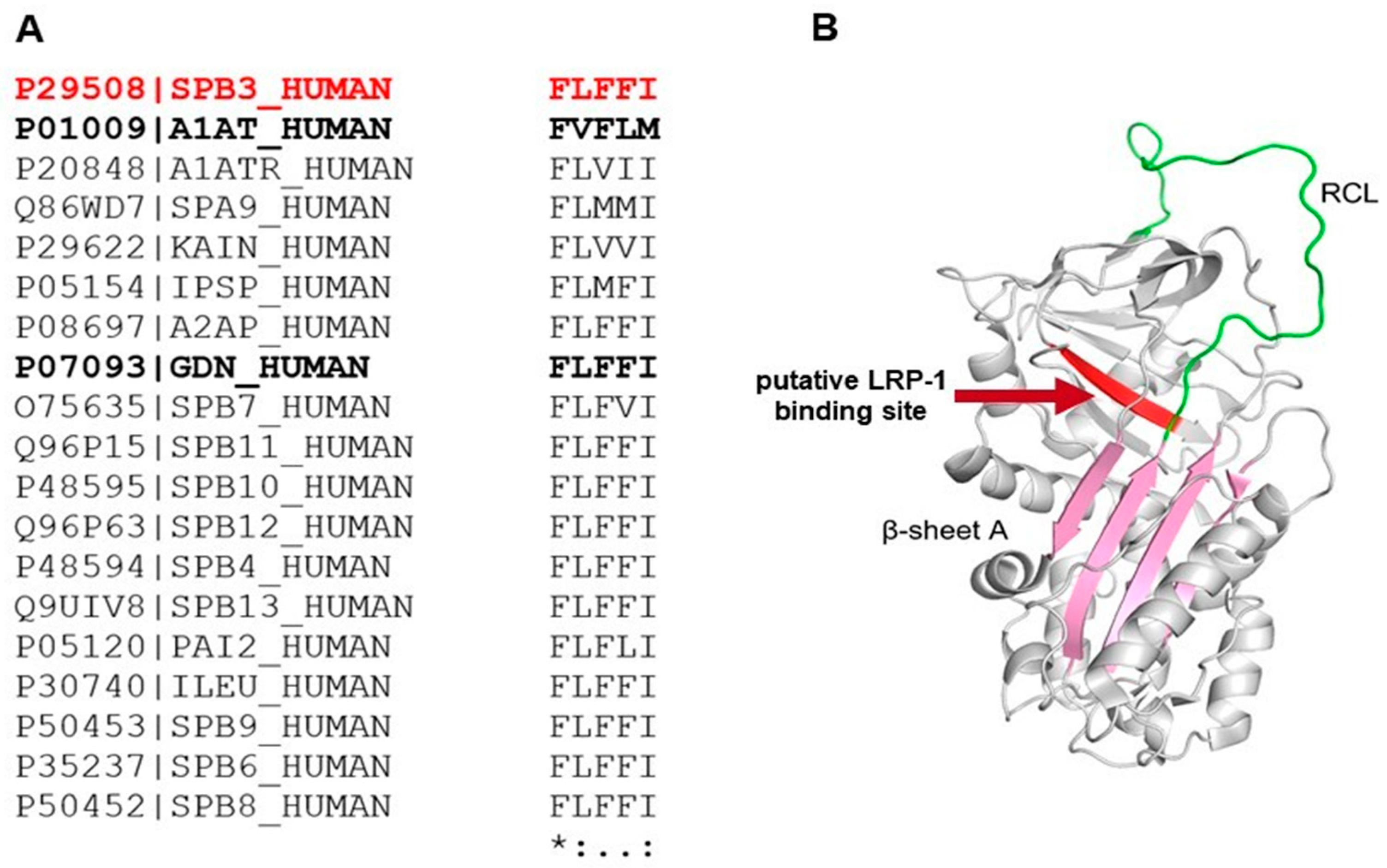
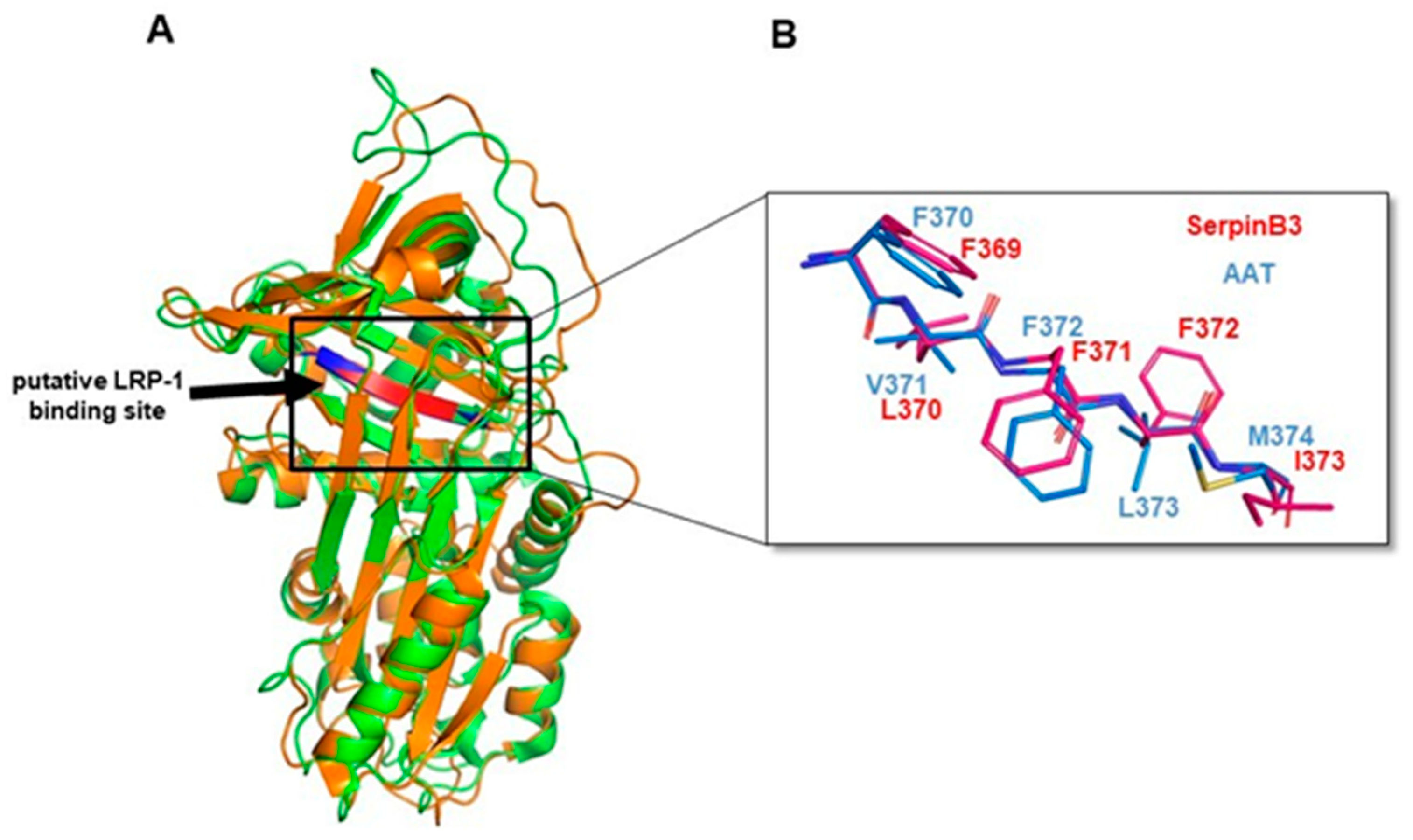
3.7. Amino Acid and Structure Alignment of Serpins
3.8. SerpinB3 Induces the Pro-Invasive Activity of LRP-1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perugorria, M.J.; Olaizola, P.; Labiano, I.; Esparza-Baquer, A.; Marzioni, M.; Marin, J.J.G.; Bujanda, L.; Banales, J.M. Wnt-β-catenin signalling in liver development, health and disease. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Boulter, L.; Govaere, O.; Bird, T.G.; Radulescu, S.; Ramachandran, P.; Pellicoro, A.; Ridgway, R.A.; Seo, S.S.; Spee, B.; Van Rooijen, N.; et al. Macrophage-derived Wnt opposes notch signaling to specify hepatic progenitor cell fate in chronic liver disease. Nat. Med. 2012, 18, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Debebe, A.; Medina, V.; Chen, C.-Y.; Mahajan, I.M.; Jia, C.; Fu, D.; He, L.; Zeng, N.; Stiles, B.W.; Chen, C.-L.; et al. Wnt/β-catenin activation and macrophage induction during liver cancer development following steatosis. Oncogene 2017, 36, 6020–6029. [Google Scholar] [CrossRef] [PubMed]
- Valenta, T.; Hausmann, G.; Basler, K. The many faces and functions of β-catenin: β-catenin: A life by, beyond, and against the Wnt canon. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [PubMed]
- Quarta, S.; Vidalino, L.; Turato, C.; Ruvoletto, M.; Calabrese, F.; Valente, M.; Cannito, S.; Fassina, G.; Parola, M.; Gatta, A.; et al. SERPINB3 induces Epithelial-Mesenchymal Transition. J. Pathol. 2010, 221, 343–356. [Google Scholar] [CrossRef]
- Terrin, L.; Agostini, M.; Ruvoletto, M.; Martini, A.; Pucciarelli, S.; Bedin, C.; Nitti, D.; Pontisso, P. SerpinB3 Upregulates the cyclooxygenase-2/β-Catenin positive loop in colorectal cancer. Oncotarget 2017, 8, 15732–15743. [Google Scholar] [CrossRef]
- Turato, C.; Vitale, A.; Fasolato, S.; Ruvoletto, M.; Terrin, L.; Quarta, S.; Ramirez Morales, R.; Biasiolo, A.; Zanus, G.; Zali, N.; et al. SERPINB3 is associated with TGF-Β1 and cytoplasmic β-catenin expression in hepatocellular carcinomas with poor prognosis. Br. J. Cancer 2014, 110, 2708–2715. [Google Scholar] [CrossRef]
- Correnti, M.; Cappon, A.; Pastore, M.; Piombanti, B.; Lori, G.; Oliveira, D.V.P.N.; Munoz-Garrido, P.; Lewinska, M.; Andersen, J.B.; Coulouarn, C.; et al. The protease-inhibitor SerpinB3 as a critical modulator of the stem-like subset in human cholangiocarcinoma. Liver Int. 2022, 42, 233–248. [Google Scholar] [CrossRef]
- Ciscato, F.; Sciacovelli, M.; Villano, G.; Turato, C.; Bernardi, P.; Rasola, A.; Pontisso, P. SERPINB3 protects from oxidative damage by chemotherapeutics through inhibition of mitochondrial respiratory complex I. Oncotarget 2014, 5, 2418–2427. [Google Scholar] [CrossRef]
- Wang, S.; Luke, C.J.; Pak, S.C.; Shi, V.; Chen, L.; Moore, J.; Andress, A.P.; Jayachandran, K.; Zhang, J.; Huang, Y.; et al. SERPINB3 (SCCA1) inhibits cathepsin L and lysoptosis, protecting cervical cancer cells from chemoradiation. Commun. Biol. 2022, 5, 46. [Google Scholar] [CrossRef]
- Turato, C.; Buendia, M.A.; Fabre, M.; Redon, M.J.; Branchereau, S.; Quarta, S.; Ruvoletto, M.; Perilongo, G.; Grotzer, M.; Gatta, A. Over-expression of SERPINB3 in hepatoblastoma: A possible insight into the genesis of this tumour? Eur. J. Cancer 2012, 48, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Wang, J.; Liu, B.; Pan, W.; Farr, G.H.; Flynn, C.; Yuan, H.; Takada, S.; Kimelman, D.; Li, L.; et al. Low-Density Lipoprotein Receptor-Related Protein-5 binds to axin and regulates the canonical Wnt signaling pathway. Mol. Cell 2001, 7, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Strickland, D.K.; Muratoglu, S.C.; Antalis, T.M. Serpin–Enzyme Receptors. Meth. Enzymol 2011, 499, 17–31. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, C.; Chen, S.; Liu, J.; Fu, Y.; Luo, Y. Extracellular Hsp90α and clusterin synergistically promote breast cancer epithelial-to-mesenchymal transition and metastasis via LRP1. J. Cell Sci. 2019, 132, jcs228213. [Google Scholar] [CrossRef] [PubMed]
- Fasolato, S.; Ruvoletto, M.; Nardo, G.; Rasola, A.; Sciacovelli, M.; Zanus, G.; Turato, C.; Quarta, S.; Terrin, L.; Fadini, G.P.; et al. Low P66shc with high SerpinB3 levels favors necroptosis and better survival in hepatocellular carcinoma. Biology 2021, 10, 363. [Google Scholar] [CrossRef]
- Turato, C.; Biasiolo, A.; Pengo, P.; Frecer, V.; Quarta, S.; Fasolato, S.; Ruvoletto, M.; Beneduce, L.; Zuin, J.; Fassina, G.; et al. Increased antiprotease activity of the SERPINB3 polymorphic variant SCCA-PD. Exp. Biol. Med. 2011, 236, 281–290. [Google Scholar] [CrossRef]
- Albiero, M.; Fullin, A.; Villano, G.; Biasiolo, A.; Quarta, S.; Bernardotto, S.; Turato, C.; Ruvoletto, M.; Fadini, G.P.; Pontisso, P.; et al. Semisolid wet Sol–Gel Silica/Hydroxypropyl methyl cellulose formulation for slow release of Serpin B3 promotes wound healing in vivo. Pharmaceutics 2022, 14, 1944. [Google Scholar] [CrossRef]
- Turato, C.; Scarpa, M.; Kotsafti, A.; Cappon, A.; Quarta, S.; Biasiolo, A.; Cavallin, F.; Trevellin, E.; Guzzardo, V.; Fassan, M.; et al. Squamous Cell Carcinoma Antigen 1 is associated to poor prognosis in esophageal cancer through immune surveillance impairment and reduced chemosensitivity. Cancer Sci. 2019, 110, 1552–1563. [Google Scholar] [CrossRef]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.-L.; Oh, S.W.; Kim, H.Y.; et al. A small molecule inhibitor of β-catenin/cyclic AMP response element-binding protein transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef]
- Foglia, B.; Sutti, S.; Cannito, S.; Rosso, C.; Maggiora, M.; Autelli, R.; Novo, E.; Villano, M.; Ramavat, N.N.; Younes, R.; et al. Hepatocyte-specific deletion of HIF2α prevents NASH-related liver carcinogenesis by decreasing cancer cell proliferation. Cell Mol. Gastgroenterol. Hepatol. 2022, 13, 459–482. [Google Scholar] [CrossRef]
- Morello, E.; Sutti, S.; Foglia, E.; Novo, E.; Cannito, S.; Bocca, C.; Rajsky, M.; Bruzzì, S.; Abate, M.L.; Rosso, C.; et al. Hypoxya-inducible factor 2α drives nonalcoholic fatty liver disease progression by triggering hepatocyte release of histidine-rich glycoprotein. Hepatology 2018, 67, 2196–2214. [Google Scholar] [CrossRef] [PubMed]
- Novo, E.; Cappon, A.; Villano, G.; Quarta, S.; Cannito, S.; Bocca, C.; Turato, C.; Guido, M.; Maggi, M.; Protopapa, F.; et al. SerpinB3 as a pro-inflammatory mediator in the progression of experimental non-alcoholic fatty liver disease. Front. Immunol. 2022, 13, 910526. [Google Scholar] [CrossRef] [PubMed]
- Juric, V.; Ruffell, B.; Evason, K.J.; Hu, J.; Che, L.; Wang, L.; Chen, X.; Bishop, M.J. Monocyte-dependent liver injury promotes carcinogenesis in an oncogene-specific manner. J. Hepatol. 2016, 64, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-catenin signalling: Function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Tamai, K.; Semenov, M.; Kato, Y.; Spokony, R.; Liu, C.; Katsuyama, Y.; Hess, F.; Saint-Jeannet, J.-P.; He, X. LDL-Receptor-Related Proteins in Wnt signal transduction. Nature 2000, 407, 530–535. [Google Scholar] [CrossRef]
- Cannito, S.; Turato, C.; Paternostro, P.; Biasiolo, A.; Colombatto, S.; Cambieri, I.; Quarta, S.; Novo, E.; Morello, E.; Rovida, E.; et al. Hypoxya up-regulates SERPINB3 through HIF-2a in human liver cancer cells. Oncotarget 2015, 6, 2206–2221. [Google Scholar] [CrossRef]
- Bu, G. The roles of Receptor-Associated Protein (RAP) as a molecular chaperone for members of the LDL receptor family. Int. Rev. Cytol. 2001, 209, 79–116. [Google Scholar] [CrossRef]
- Joslin, G.; Fallon, R.J.; Bullock, J.; Adams, S.P.; Perlmutter, D.H. The SEC receptor recognizes a pentapeptide neodomain of alpha 1-antitrypsin-protease complexes. J. Biol. Chem. 1991, 266, 11282–11288. [Google Scholar] [CrossRef]
- Toldo, S.; Austin, D.; Mauro, A.G.; Mezzaroma, E.; Van Tassell, B.W.; Marchetti, C.; Carbone, S.; Mogelsvang, S.; Gelber, C.; Abbate, A. Low-Density Lipoprotein Receptor–Related Protein-1 is a therapeutic target in acute myocardial infarction. JACC Basic Transl. Sci. 2017, 2, 561–574. [Google Scholar] [CrossRef]
- Chiu, I.-J.; Hsu, Y.-H.; Chang, J.-S.; Yang, J.-C.; Chiu, H.-W.; Lin, Y.-F. Lactotransferrin Downregulation Drives the Metastatic Progression in Clear Cell Renal Cell Carcinoma. Cancers 2020, 12, 847. [Google Scholar] [CrossRef]
- Fayard, B.; Bianchi, F.; Dey, J.; Moreno, E.; Djaffer, S.; Hynes, N.E.; Monard, D. The Serine Protease Inhibitor Protease Nexin-1 controls mammary cancer metastasis through LRP-1–mediated MMP-9 expression. Cancer Res. 2009, 69, 5690–5698. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quarta, S.; Cappon, A.; Turato, C.; Ruvoletto, M.; Cannito, S.; Villano, G.; Biasiolo, A.; Maggi, M.; Protopapa, F.; Bertazza, L.; et al. SerpinB3 Upregulates Low-Density Lipoprotein Receptor-Related Protein (LRP) Family Members, Leading to Wnt Signaling Activation and Increased Cell Survival and Invasiveness. Biology 2023, 12, 771. https://doi.org/10.3390/biology12060771
Quarta S, Cappon A, Turato C, Ruvoletto M, Cannito S, Villano G, Biasiolo A, Maggi M, Protopapa F, Bertazza L, et al. SerpinB3 Upregulates Low-Density Lipoprotein Receptor-Related Protein (LRP) Family Members, Leading to Wnt Signaling Activation and Increased Cell Survival and Invasiveness. Biology. 2023; 12(6):771. https://doi.org/10.3390/biology12060771
Chicago/Turabian StyleQuarta, Santina, Andrea Cappon, Cristian Turato, Mariagrazia Ruvoletto, Stefania Cannito, Gianmarco Villano, Alessandra Biasiolo, Maristella Maggi, Francesca Protopapa, Loris Bertazza, and et al. 2023. "SerpinB3 Upregulates Low-Density Lipoprotein Receptor-Related Protein (LRP) Family Members, Leading to Wnt Signaling Activation and Increased Cell Survival and Invasiveness" Biology 12, no. 6: 771. https://doi.org/10.3390/biology12060771