

When Corticospinal Inhibition Favors an Efficient Motor Response
 , , , and
, , , and 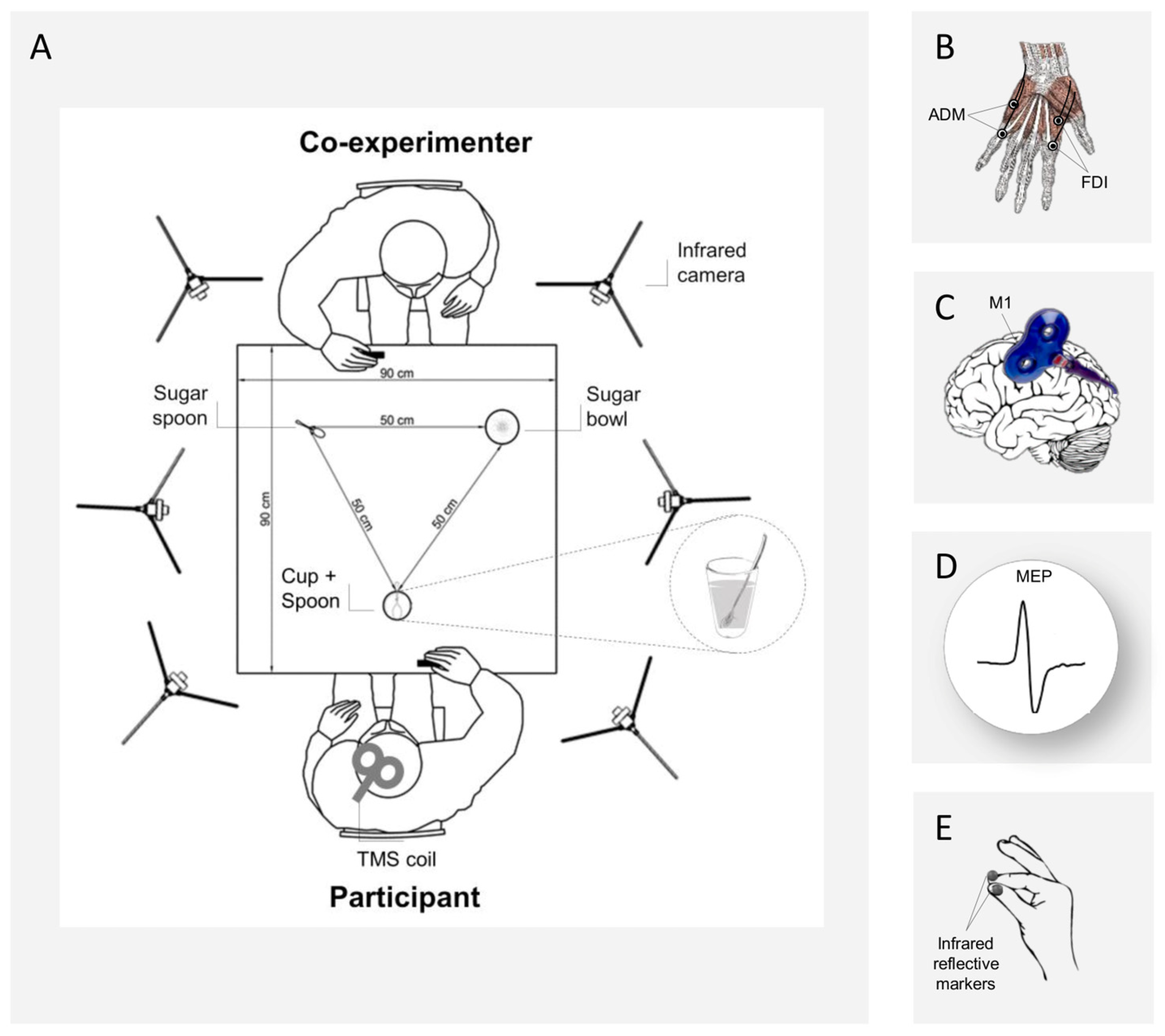{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Paradigm
- Interactive request, precision grip execution: the co-experimenter grasped the sugar spoon, took some sugar from a sugar bowl, and then stretched out her arm toward the participant’s cup, as if to pour the sugar in it. Participants had to reach and grasp their teaspoon with a PG, and to stir the coffee;
- Non-Interactive action, precision grip execution: the co-experimenter grasped the sugar spoon, took some sugar from the sugar bowl and then returned to the initial position. Participants had to reach and grasp their teaspoon with a PG, and to stir the coffee;
- Interactive request, whole-hand grasp execution: the co-experimenter grasped the sugar spoon, took some sugar from a sugar bowl, and then stretched out her arm toward the participant’s cup, as if to pour some sugar in it. Participants had to reach and grasp the cup with a WHG and to lift it up (i.e., bringing it closer to the teaspoon full of sugar);
- Non-Interactive action, whole-hand grasp execution: the co-experimenter grasped the sugar spoon, took some sugar from a sugar bowl and then returned to the initial position. Participants had to reach and grasp the cup with a WHG and to lift it up.
2.3. Validation Study
2.4. Procedure
2.5. Kinematic Recording
2.6. Electromyography
2.7. Transcranial Magnetic Stimulation
2.8. Interpersonal Reactivity Index Questionnaire
2.9. Data Preparation
2.9.1. MEP Data
2.9.2. EMG Data
2.9.3. Reaction Times
2.9.4. Kinematic Data
2.9.5. Interpersonal Reactivity Index Questionnaire
2.10. Statistical Analysis
3. Results
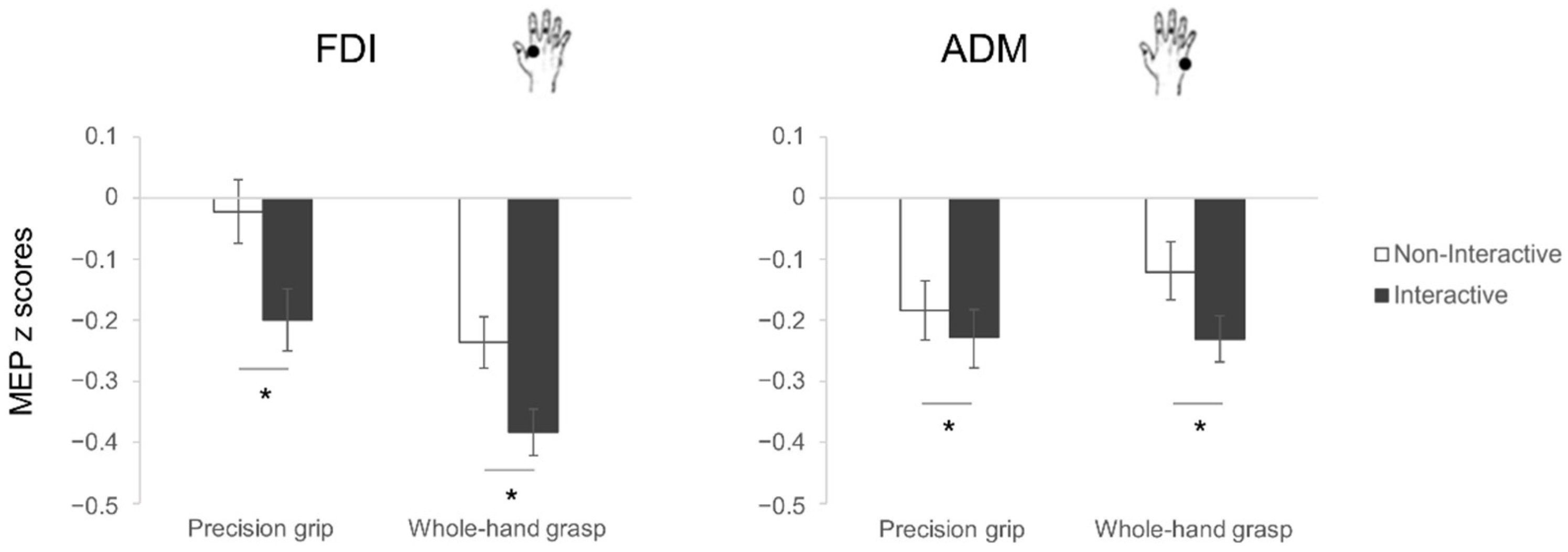
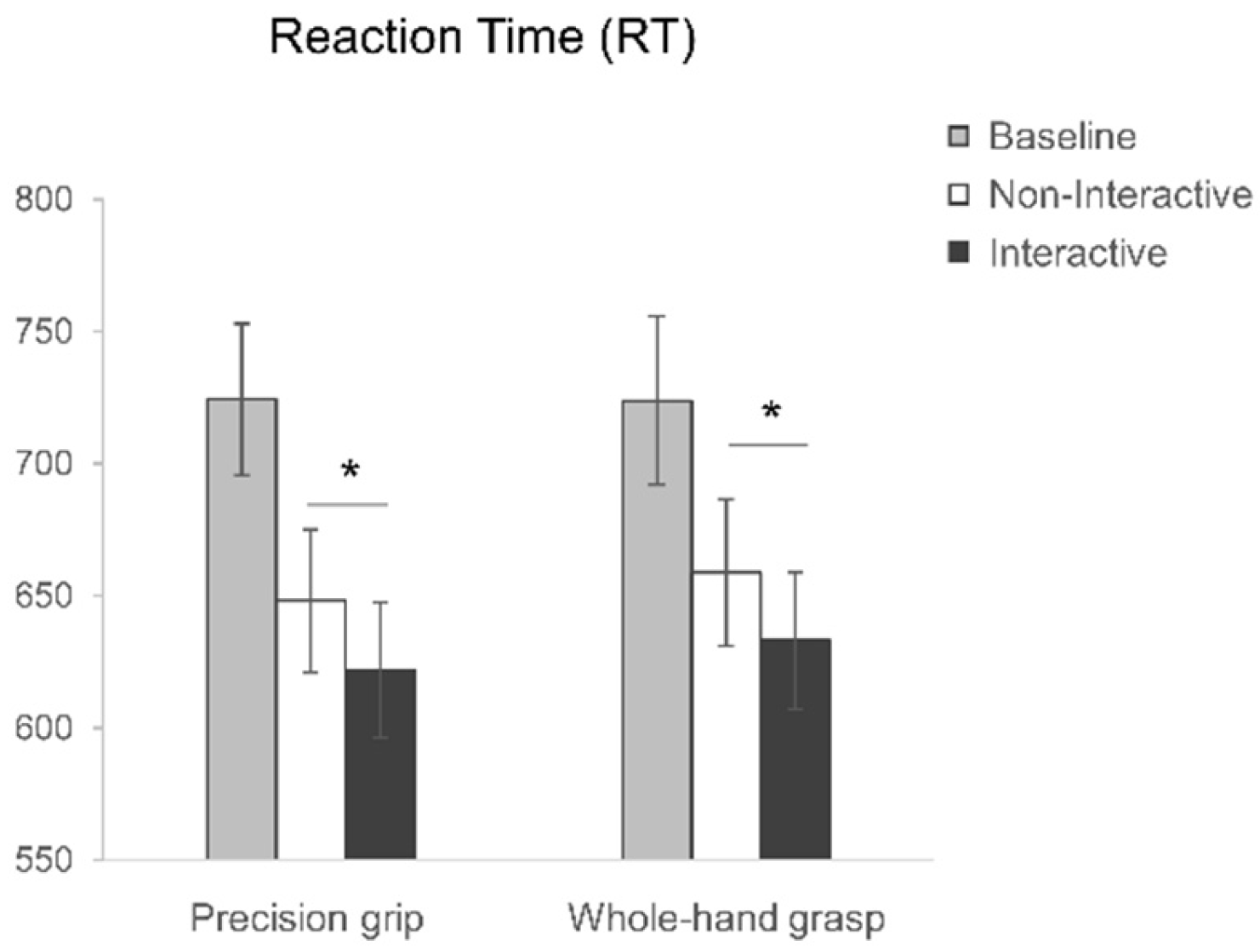
3.1. Motor Preparation
3.1.1. Motor-Evoked Potentials
3.1.2. Reaction Times
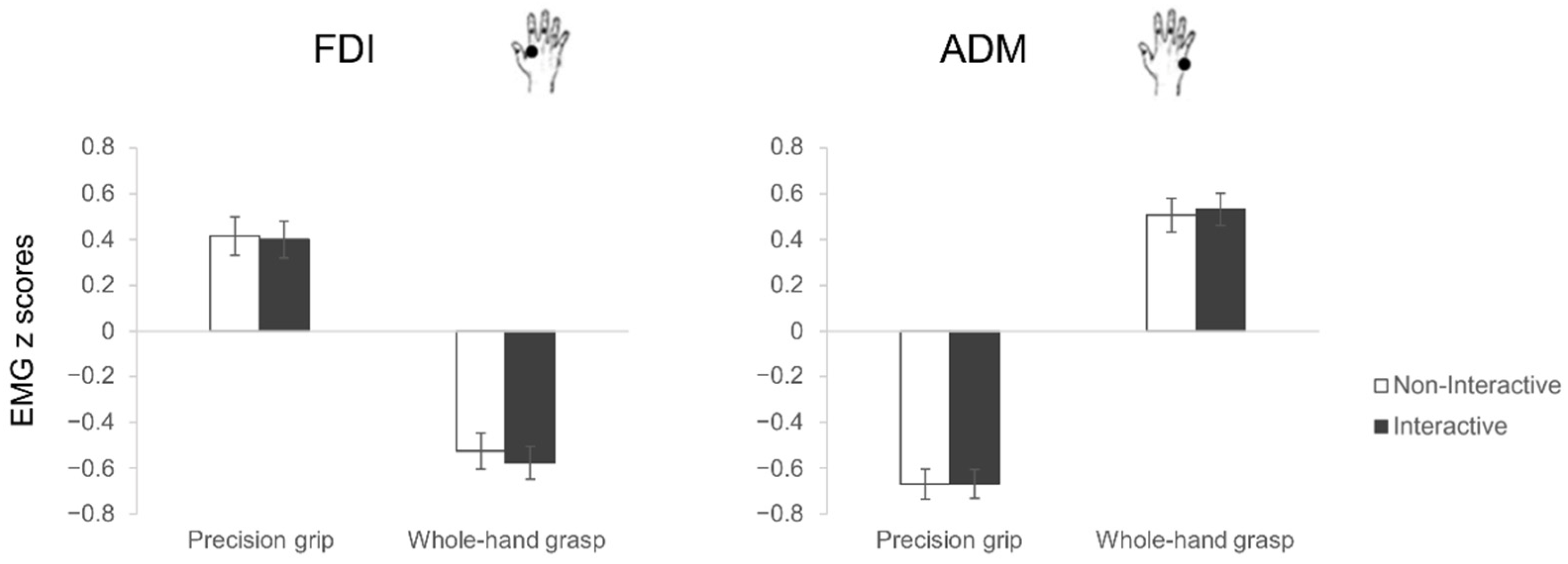
3.2. Motor Execution
3.2.1. Electromyography
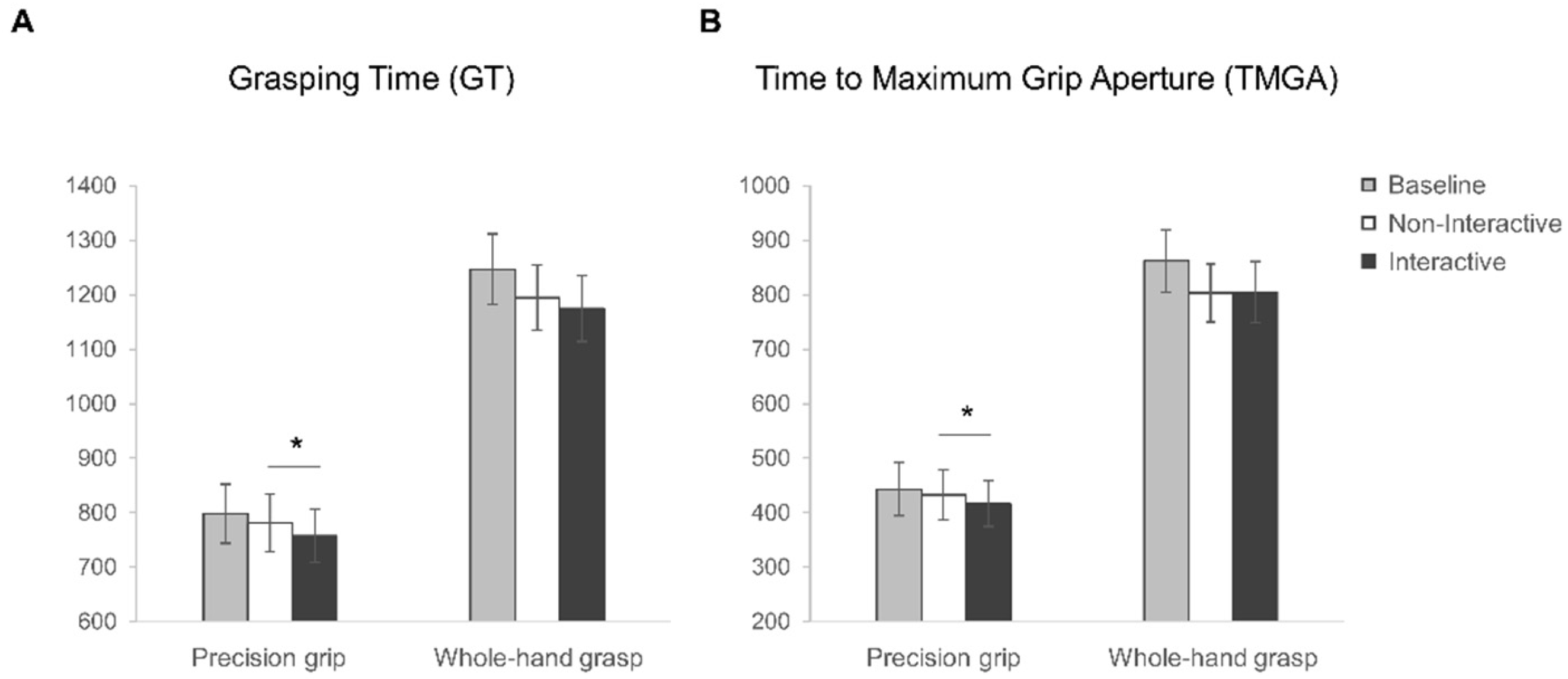
3.2.2. Kinematics
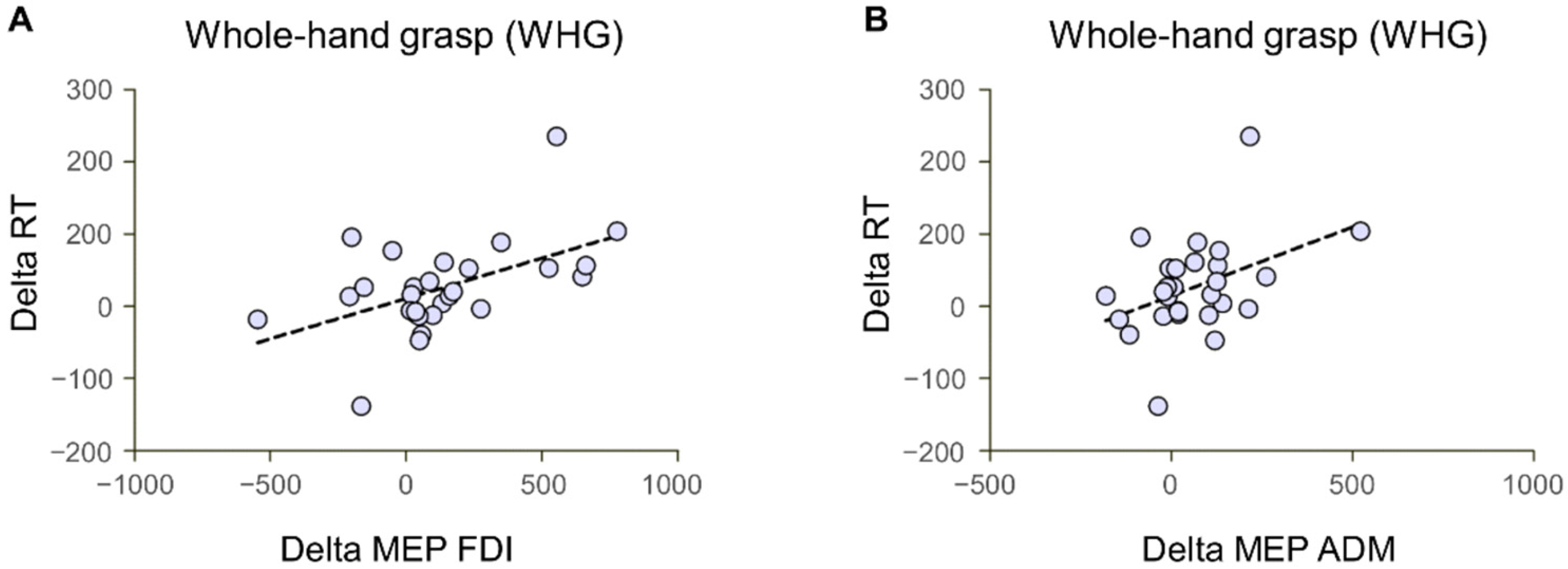
3.3. Correlations
4. Discussion
4.1. Social Motor Inhibition
4.2. Social Motor Priming
4.3. The Encoding of Incomplete Joint Actions
4.4. Future Applications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kilner, J.M.; Vargas, C.; Duval, S.; Blakemore, S.-J.; Sirigu, A. Motor Activation Prior to Observation of a Predicted Movement. Nat. Neurosci. 2004, 7, 1299–1301. [Google Scholar] [CrossRef] [PubMed]
- Lelonkiewicz, J.R.; Gambi, C.; Weller, L.; Pfister, R. Action–Effect Anticipation and Temporal Adaptation in Social Interactions. J. Exp. Psychol. Hum. Percept. Perform. 2020, 46, 335–349. [Google Scholar] [CrossRef]
- Sacheli, L.M.; Tidoni, E.; Pavone, E.F.; Aglioti, S.M.; Candidi, M. Kinematics Fingerprints of Leader and Follower Role-Taking during Cooperative Joint Actions. Exp. Brain Res. 2013, 226, 473–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacheli, L.M.; Arcangeli, E.; Carioti, D.; Butterfill, S.; Berlingeri, M. Taking Apart What Brings Us Together: The Role of Action Prediction, Perspective-Taking, and Theory of Mind in Joint Action. Q. J. Exp. Psychol. 2021, 75, 17470218211050198. [Google Scholar] [CrossRef]
- Sebanz, N.; Knoblich, G. Prediction in Joint Action: What, When, and Where. Top. Cogn. Sci. 2009, 1, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Gallotti, M.; Fairhurst, M.T.; Frith, C.D. Alignment in Social Interactions. Conscious. Cogn. 2017, 48, 253–261. [Google Scholar] [CrossRef]
- Sartori, L.; Becchio, C.; Castiello, U. Cues to Intention: The Role of Movement Information. Cognition 2011, 119, 242–252. [Google Scholar] [CrossRef]
- Sartori, L.; Bucchioni, G.; Castiello, U. When Emulation Becomes Reciprocity. Soc. Cogn. Affect. Neurosci. 2013, 8, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Sartori, L.; Betti, S.; Chinellato, E.; Castiello, U. The Multiform Motor Cortical Output: Kinematic, Predictive and Response Coding. Cortex 2015, 70, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Sartori, L.; Betti, S.; Perrone, C.; Castiello, U. Congruent and Incongruent Corticospinal Activations at the Level of Multiple Effectors. J. Cogn. Neurosci. 2015, 27, 2063–2070. [Google Scholar] [CrossRef] [Green Version]
- Pesquita, A.; Bernardet, U.; Richards, B.E.; Jensen, O.; Shapiro, K. Isolating Action Prediction from Action Integration in the Perception of Social Interactions. Brain Sci. 2022, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Sartori, L.; Betti, S. Complementary Actions. Front. Psychol. 2015, 6, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naish, K.R.; Houston-Price, C.; Bremner, A.J.; Holmes, N.P. Effects of Action Observation on Corticospinal Excitability: Muscle Specificity, Direction, and Timing of the Mirror Response. Neuropsychologia 2014, 64, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Cardellicchio, P.; Dolfini, E.; Fadiga, L.; D’Ausilio, A. Parallel Fast and Slow Motor Inhibition Processes in Joint Action Coordination. Cortex 2020, 133, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Cardellicchio, P.; Dolfini, E.; Hilt, P.M.; Fadiga, L.; D’Ausilio, A. Motor Cortical Inhibition during Concurrent Action Execution and Action Observation. NeuroImage 2020, 208, 116445. [Google Scholar] [CrossRef]
- Sartori, L.; Cavallo, A.; Bucchioni, G.; Castiello, U. Corticospinal Excitability Is Specifically Modulated by the Social Dimension of Observed Actions. Exp. Brain Res. 2011, 211, 557. [Google Scholar] [CrossRef] [PubMed]
- Sartori, L.; Cavallo, A.; Bucchioni, G.; Castiello, U. From Simulation to Reciprocity: The Case of Complementary Actions. Soc. Neurosci. 2012, 7, 146–158. [Google Scholar] [CrossRef]
- Betti, S.; Zani, G.; Granziol, U.; Guerra, S.; Castiello, U.; Sartori, L. Look at Me: Early Gaze Engagement Enhances Corticospinal Excitability during Action Observation. Front. Psychol. 2018, 9, 1408. [Google Scholar] [CrossRef]
- Betti, S.; Castiello, U.; Guerra, S.; Granziol, U.; Zani, G.; Sartori, L. Gaze and Body Cues Interplay during Interactive Requests. PLoS ONE 2019, 14, e0223591. [Google Scholar] [CrossRef] [Green Version]
- Dezecache, G.; Conty, L.; Grèzes, J. Social Affordances: Is the Mirror Neuron System Involved? Behav. Brain Sci. 2013, 36, 417–418. [Google Scholar] [CrossRef]
- Gallagher, H.L.; Frith, C.D. Dissociable Neural Pathways for the Perception and Recognition of Expressive and Instrumental Gestures. Neuropsychologia 2004, 42, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Betti, S.; Zani, G.; Guerra, S.; Castiello, U.; Sartori, L. Reach-to-Grasp Movements: A Multimodal Techniques Study. Front. Psychol. 2018, 9, 990. [Google Scholar] [CrossRef] [PubMed]
- Albiero, P.; Ingoglia, S.; Lo Coco, A. Contributo All’adattamento Italiano Dell’Interpersonal Reactivity Index. TPM–Test. Psychom. Methodol. Appl. Psychol. 2006, 13, 107–125. [Google Scholar]
- Davis, M.H. Measuring Individual Differences in Empathy: Evidence for a Multidimensional Approach. J. Personal. Soc. Psychol. 1983, 44, 113–126. [Google Scholar] [CrossRef]
- Coxon, J.P.; Stinear, C.M.; Byblow, W.D. Intracortical Inhibition during Volitional Inhibition of Prepared Action. J. Neurophysiol. 2006, 95, 3371–3383. [Google Scholar] [CrossRef]
- Duque, J.; Greenhouse, I.; Labruna, L.; Ivry, R.B. Physiological Markers of Motor Inhibition during Human Behavior. Trends Neurosci. 2017, 40, 219–236. [Google Scholar] [CrossRef] [Green Version]
- Betti, S.; Chinellato, E.; Guerra, S.; Castiello, U.; Sartori, L. Social Motor Priming: When Offline Interference Facilitates Motor Execution. PeerJ 2019, 7, e7796. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety of TMS Consensus Group Safety, Ethical Considerations, and Application Guidelines for the Use of Transcranial Magnetic Stimulation in Clinical Practice and Research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Wassermann, E.M. Risk and Safety of Repetitive Transcranial Magnetic Stimulation: Report and Suggested Guidelines from the International Workshop on the Safety of Repetitive Transcranial Magnetic Stimulation, June 5–7, 1996. Electroencephalogr. Clin. Neurophysiol./Evoked Potentials Sect. 1998, 108, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kemmerer, D. What Modulates the Mirror Neuron System during Action Observation?: Multiple Factors Involving the Action, the Actor, the Observer, the Relationship between Actor and Observer, and the Context. Prog. Neurobiol. 2021, 205, 102128. [Google Scholar] [CrossRef]
- Naish, K.R.; Obhi, S.S. Self-Selected Conscious Strategies Do Not Modulate Motor Cortical Output during Action Observation. J. Neurophysiol. 2015, 114, 2278–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becchio, C.; Sartori, L.; Bulgheroni, M.; Castiello, U. The Case of Dr. Jekyll and Mr. Hyde: A Kinematic Study on Social Intention. Conscious. Cogn. 2008, 17, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Jeannerod, M. Intersegmental Coordination during Reaching at Natural Visual Objects. In Attention and Performance IX; Long, J., Baddeley, A., Eds.; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1981; pp. 153–168. [Google Scholar]
- Jeannerod, M. The Timing of Natural Prehension Movements. J. Mot. Behav. 1984, 16, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, G.; De Stefani, E.; Gentilucci, M.; De Marco, D. Vitality Forms Expressed by Others Modulate Our Own Motor Response: A Kinematic Study. Front. Hum. Neurosci. 2017, 11, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasil-Neto, J.P.; Cohen, L.G.; Panizza, M.; Nilsson, J.; Roth, B.J.; Hallett, M. Optimal Focal Transcranial Magnetic Activation of the Human Motor Cortex: Effects of Coil Orientation, Shape of the Induced Current Pulse, and Stimulus Intensity. J. Clin. Neurophysiol. 1992, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.R.; Boniface, S.J.; Schubert, M. Magnetic Brain Stimulation with a Double Coil: The Importance of Coil Orientation. Electroencephalogr. Clin. Neurophysiol./Evoked Potentials Sect. 1992, 85, 17–21. [Google Scholar] [CrossRef]
- Rossini, P.M.; Barker, A.T.; Berardelli, A.; Caramia, M.D.; Caruso, G.; Cracco, R.Q.; Dimitrijević, M.R.; Hallett, M.; Katayama, Y.; Lücking, C.H.; et al. Non-Invasive Electrical and Magnetic Stimulation of the Brain, Spinal Cord and Roots: Basic Principles and Procedures for Routine Clinical Application. Report of an IFCN Committee. Electroencephalogr. Clin. Neurophysiol. 1994, 91, 79–92. [Google Scholar] [CrossRef]
- Gentilucci, M.; Castiello, U.; Corradini, M.L.; Scarpa, M.; Umiltà, C.; Rizzolatti, G. Influence of Different Types of Grasping on the Transport Component of Prehension Movements. Neuropsychologia 1991, 29, 361–378. [Google Scholar] [CrossRef]
- Davis, M. A Multidimensional Approach to Individual Differences in Empathy. JSAS Cat. Sel. Doc. Psychol. 1980, 10, 85. [Google Scholar]
- Sartori, L.; Betti, S.; Castiello, U. Corticospinal Excitability Modulation during Action Observation. J. Vis. Exp. 2013, 82, e51001. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, E.; De Jaegher, H. The Interactive Brain Hypothesis. Front. Hum. Neurosci. 2012, 6, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aglioti, S.M.; Cesari, P.; Romani, M.; Urgesi, C. Action Anticipation and Motor Resonance in Elite Basketball Players. Nat. Neurosci. 2008, 11, 1109. [Google Scholar] [CrossRef] [PubMed]
- Gangitano, M.; Mottaghy, F.M.; Pascual-Leone, A. Phase-Specific Modulation of Cortical Motor Output during Movement Observation. NeuroReport 2001, 12, 1489. [Google Scholar] [CrossRef]
- Gangitano, M.; Mottaghy, F.M.; Pascual-Leone, A. Modulation of Premotor Mirror Neuron Activity during Observation of Unpredictable Grasping Movements. Eur. J. Neurosci. 2004, 20, 2193–2202. [Google Scholar] [CrossRef] [Green Version]
- Urgesi, C.; Maieron, M.; Avenanti, A.; Tidoni, E.; Fabbro, F.; Aglioti, S.M. Simulating the Future of Actions in the Human Corticospinal System. Cereb. Cortex 2010, 20, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- Sartori, L.; Becchio, C.; Bulgheroni, M.; Castiello, U. Modulation of the Action Control System by Social Intention: Unexpected Social Requests Override Preplanned Action. J. Exp. Psychol. Hum. Percept. Perform. 2009, 35, 1490–1500. [Google Scholar] [CrossRef]
- Schuch, S.; Bayliss, A.P.; Klein, C.; Tipper, S.P. Attention Modulates Motor System Activation during Action Observation: Evidence for Inhibitory Rebound. Exp. Brain Res. 2010, 205, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Cisek, P. Cortical Mechanisms of Action Selection: The Affordance Competition Hypothesis. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1585–1599. [Google Scholar] [CrossRef]
- Wijnen, J.G.; Ridderinkhof, K.R. Response Inhibition in Motor and Oculomotor Conflict Tasks: Different Mechanisms, Different Dynamics? Brain Cogn. 2007, 63, 260–270. [Google Scholar] [CrossRef]
- Duque, J.; Mazzocchio, R.; Dambrosia, J.; Murase, N.; Olivier, E.; Cohen, L.G. Kinematically Specific Interhemispheric Inhibition Operating in the Process of Generation of a Voluntary Movement. Cereb. Cortex 2005, 15, 588–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leocani, L.; Cohen, L.G.; Wassermann, E.M.; Ikoma, K.; Hallett, M. Human Corticospinal Excitability Evaluated with Transcranial Magnetic Stimulation during Different Reaction Time Paradigms. Brain 2000, 123, 1161–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liepert, J.; Dettmers, C.; Terborg, C.; Weiller, C. Inhibition of Ipsilateral Motor Cortex during Phasic Generation of Low Force. Clin. Neurophysiol. 2001, 112, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Sohn, Y.H.; Jung, H.Y.; Kaelin-Lang, A.; Hallett, M. Excitability of the Ipsilateral Motor Cortex during Phasic Voluntary Hand Movement. Exp. Brain Res. 2003, 148, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.C.; Weiller, C.; Liepert, J. Pre-Movement Motor Excitability Is Reduced Ipsilateral to Low Force Pinch Grips. J. Neural. Transm. 2003, 110, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Duque, J.; Lew, D.; Mazzocchio, R.; Olivier, E.; Ivry, R.B. Evidence for Two Concurrent Inhibitory Mechanisms during Response Preparation. J. Neurosci. 2010, 30, 3793–3802. [Google Scholar] [CrossRef] [Green Version]
- Duque, J.; Labruna, L.; Verset, S.; Olivier, E.; Ivry, R.B. Dissociating the Role of Prefrontal and Premotor Cortices in Controlling Inhibitory Mechanisms during Motor Preparation. J. Neurosci. 2012, 32, 806–816. [Google Scholar] [CrossRef] [Green Version]
- Boulinguez, P.; Jaffard, M.; Granjon, L.; Benraiss, A. Warning Signals Induce Automatic EMG Activations and Proactive Volitional Inhibition: Evidence from Analysis of Error Distribution in Simple RT. J. Neurophysiol. 2008, 99, 1572–1578. [Google Scholar] [CrossRef] [Green Version]
- Duque, J.; Ivry, R.B. Role of Corticospinal Suppression during Motor Preparation. Cereb. Cortex 2009, 19, 2013–2024. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, C.; Hammond, G.R. Excitatory and Inhibitory Processes in Primary Motor Cortex during the Foreperiod of a Warned Reaction Time Task Are Unrelated to Response Expectancy. Exp. Brain Res. 2009, 194, 103–113. [Google Scholar] [CrossRef]
- Greenhouse, I.; Sias, A.; Labruna, L.; Ivry, R.B. Nonspecific Inhibition of the Motor System during Response Preparation. J. Neurosci. 2015, 35, 10675–10684. [Google Scholar] [CrossRef] [Green Version]
- Davranche, K.; Tandonnet, C.; Burle, B.; Meynier, C.; Vidal, F.; Hasbroucq, T. The Dual Nature of Time Preparation: Neural Activation and Suppression Revealed by Transcranial Magnetic Stimulation of the Motor Cortex. Eur. J. Neurosci. 2007, 25, 3766–3774. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, C.; Hammond, G.R. Reduced Intracortical Inhibition during the Foreperiod of a Warned Reaction Time Task. Exp. Brain Res. 2008, 186, 385–392. [Google Scholar] [CrossRef]
- Bruno, V.; Fossataro, C.; Garbarini, F. Inhibition or Facilitation? Modulation of Corticospinal Excitability during Motor Imagery. Neuropsychologia 2018, 111, 360–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craighero, L.; Fadiga, L.; Umiltà, C.A.; Rizzolatti, G. Evidence for Visuomotor Priming Effect. NeuroReport 1996, 8, 347–349. [Google Scholar] [CrossRef]
- Brass, M.; Bekkering, H.; Wohlschläger, A.; Prinz, W. Compatibility between Observed and Executed Finger Movements: Comparing Symbolic, Spatial, and Imitative Cues. Brain Cogn. 2000, 44, 124–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinellato, E.; Castiello, U.; Sartori, L. Motor Interference in Interactive Contexts. Front. Psychol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becchio, C.; Sartori, L.; Bulgheroni, M.; Castiello, U. Both Your Intention and Mine Are Reflected in the Kinematics of My Reach-to-Grasp Movement. Cognition 2008, 106, 894–912. [Google Scholar] [CrossRef]
- Georgiou, I.; Becchio, C.; Glover, S.; Castiello, U. Different Action Patterns for Cooperative and Competitive Behaviour. Cognition 2007, 102, 415–433. [Google Scholar] [CrossRef]
- Heyes, C. Automatic Imitation. Psychol. Bull. 2011, 137, 463–483. [Google Scholar] [CrossRef]
- Flach, R.; Press, C.; Badets, A.; Heyes, C. Shaking Hands: Priming by Social Action Effects. Br. J. Psychol. 2010, 101, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Liepelt, R.; Prinz, W.; Brass, M. When Do We Simulate Non-Human Agents? Dissociating Communicative and Non-Communicative Actions. Cognition 2010, 115, 426–434. [Google Scholar] [CrossRef] [Green Version]
- de C Hamilton, A.F. The Mirror Neuron System Contributes to Social Responding. Cortex 2013, 49, 2957–2959. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; de C Hamilton, A.F. Understanding the Role of the ‘Self’ in the Social Priming of Mimicry. PLoS ONE 2013, 8, e60249. [Google Scholar] [CrossRef] [Green Version]
- Sartori, L.; Straulino, E.; Castiello, U. How Objects Are Grasped: The Interplay between Affordances and End-Goals. PLoS ONE 2011, 6, e25203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crajé, C.; Lukos, J.R.; Ansuini, C.; Gordon, A.M.; Santello, M. The Effects of Task and Content on Digit Placement on a Bottle. Exp. Brain Res. 2011, 212, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Kawai, S.; Ikuta, K. Contributions and Co-Ordination of Individual Fingers in Multiple Finger Prehension. Ergonomics 1995, 38, 1212–1230. [Google Scholar] [CrossRef] [PubMed]
- Santello, M.; Soechting, J.F. Force Synergies for Multifingered Grasping. Exp. Brain Res. 2000, 133, 457–467. [Google Scholar] [CrossRef]
- Stern, D.N. Forms of Vitality Exploring Dynamic Experience in Psychology, Arts, Psychotherapy, and Development; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Pfister, R.; Dignath, D.; Hommel, B.; Kunde, W. It Takes Two to Imitate: Anticipation and Imitation in Social Interaction. Psychol. Sci. 2013, 24, 2117–2121. [Google Scholar] [CrossRef] [Green Version]
- Kilner, J.M.; Friston, K.J.; Frith, C.D. Predictive Coding: An Account of the Mirror Neuron System. Cogn. Process. 2007, 8, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Betti, S.; Finisguerra, A.; Amoruso, L.; Urgesi, C. Contextual Priors Guide Perception and Motor Responses to Observed Actions. Cereb. Cortex 2021, 32, 608–625. [Google Scholar] [CrossRef]
- Ding, X.; Gao, Z.; Shen, M. Two Equals One: Two Human Actions during Social Interaction Are Grouped as One Unit in Working Memory. Psychol. Sci. 2017, 28, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Papeo, L. Twos in Human Visual Perception. Cortex 2020, 132, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Vestner, T.; Tipper, S.P.; Hartley, T.; Over, H.; Rueschemeyer, S.-A. Bound Together: Social Binding Leads to Faster Processing, Spatial Distortion, and Enhanced Memory of Interacting Partners. J. Exp. Psychol. Gen. 2019, 148, 1251–1268. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.L.; Wagemans, J. Gestalts at Threshold Could Reveal Gestalts as Predictions. Sci. Rep. 2021, 11, 18308. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betti, S.; Zani, G.; Guerra, S.; Granziol, U.; Castiello, U.; Begliomini, C.; Sartori, L. When Corticospinal Inhibition Favors an Efficient Motor Response. Biology 2023, 12, 332. https://doi.org/10.3390/biology12020332
Betti S, Zani G, Guerra S, Granziol U, Castiello U, Begliomini C, Sartori L. When Corticospinal Inhibition Favors an Efficient Motor Response. Biology. 2023; 12(2):332. https://doi.org/10.3390/biology12020332
Chicago/Turabian StyleBetti, Sonia, Giovanni Zani, Silvia Guerra, Umberto Granziol, Umberto Castiello, Chiara Begliomini, and Luisa Sartori. 2023. "When Corticospinal Inhibition Favors an Efficient Motor Response" Biology 12, no. 2: 332. https://doi.org/10.3390/biology12020332