
DnFCA Isoforms Cooperatively Regulate Temperature-Related Flowering in Dendrobium nobile

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Isolation of Alternative Polyadenylation Isoforms
2.3. Generation of Arabidopsis Transgenic Lines and Phenotype Analysis
2.4. Generation of Transgenic D. nobile
2.5. Transcript Quantitation in Response to Temperature Treatment
2.6. Data Analysis and Statistical Tests
3. Results
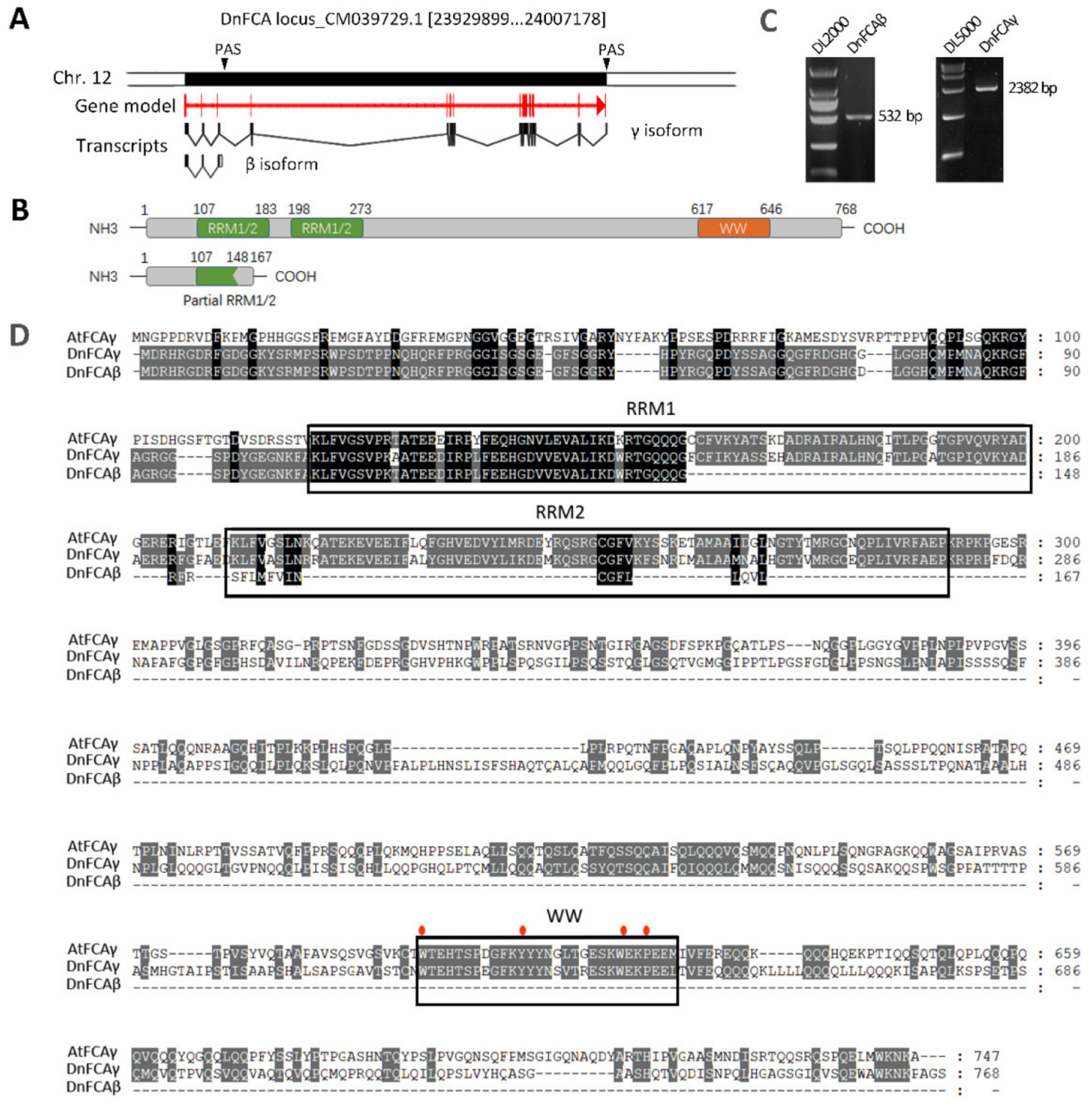
3.1. DnFCA Produces γ and β Isoforms by Alternative 3′ Polyadenylation
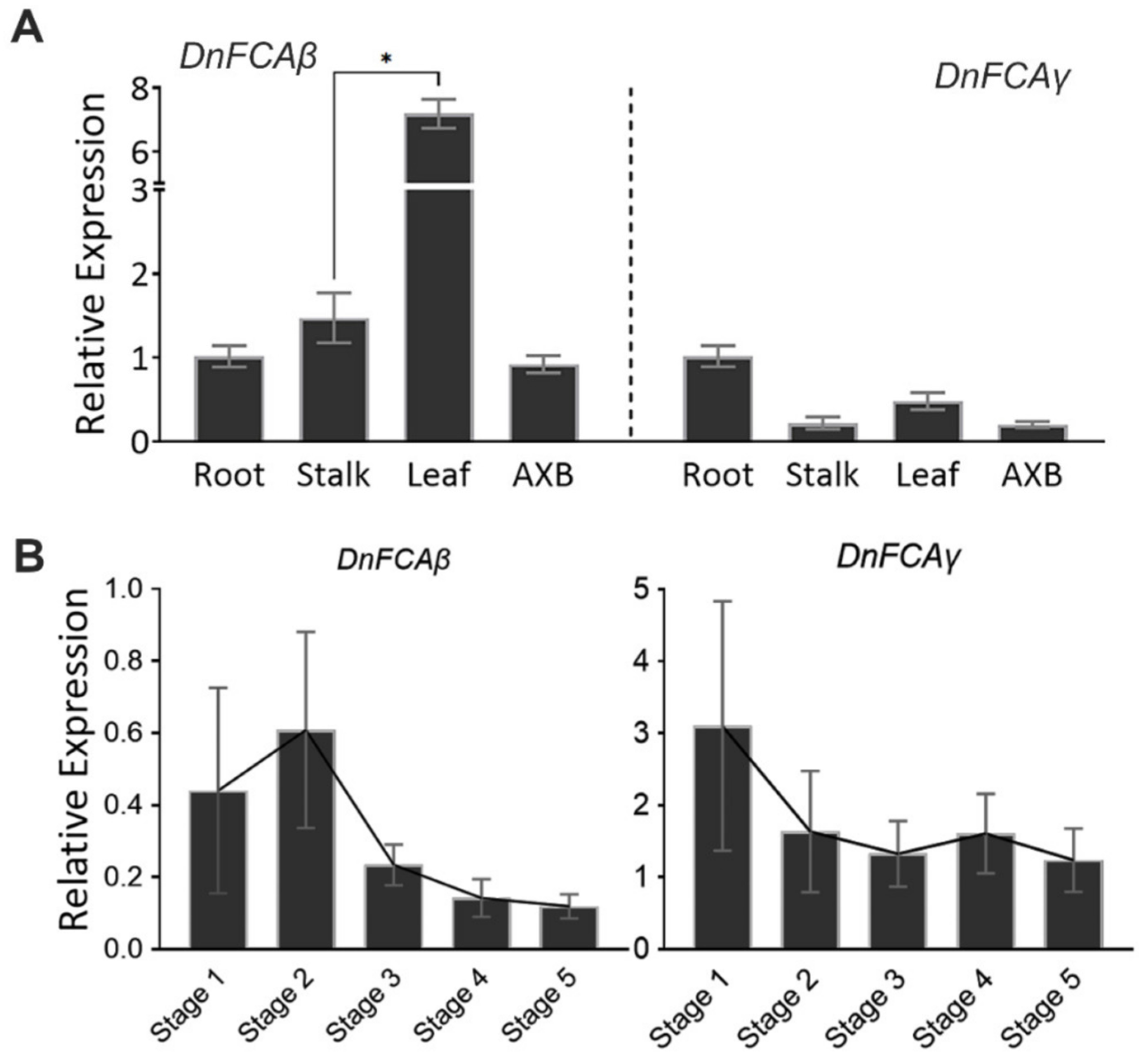
3.2. Expression of the DnFCAγ and β Isoforms
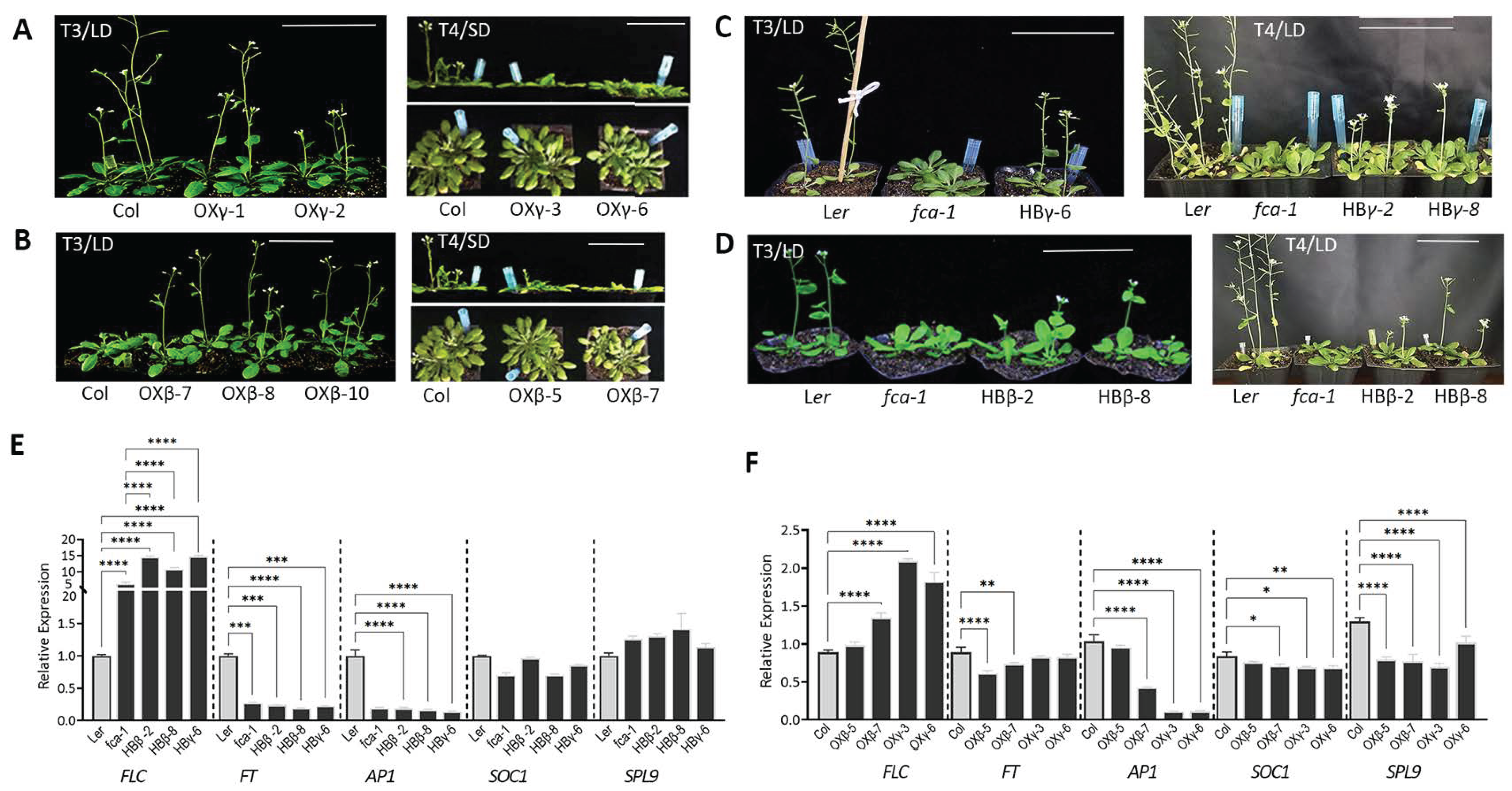
3.3. DnFCAγ and β Isoforms Regulate Flowering in Arabidopsis
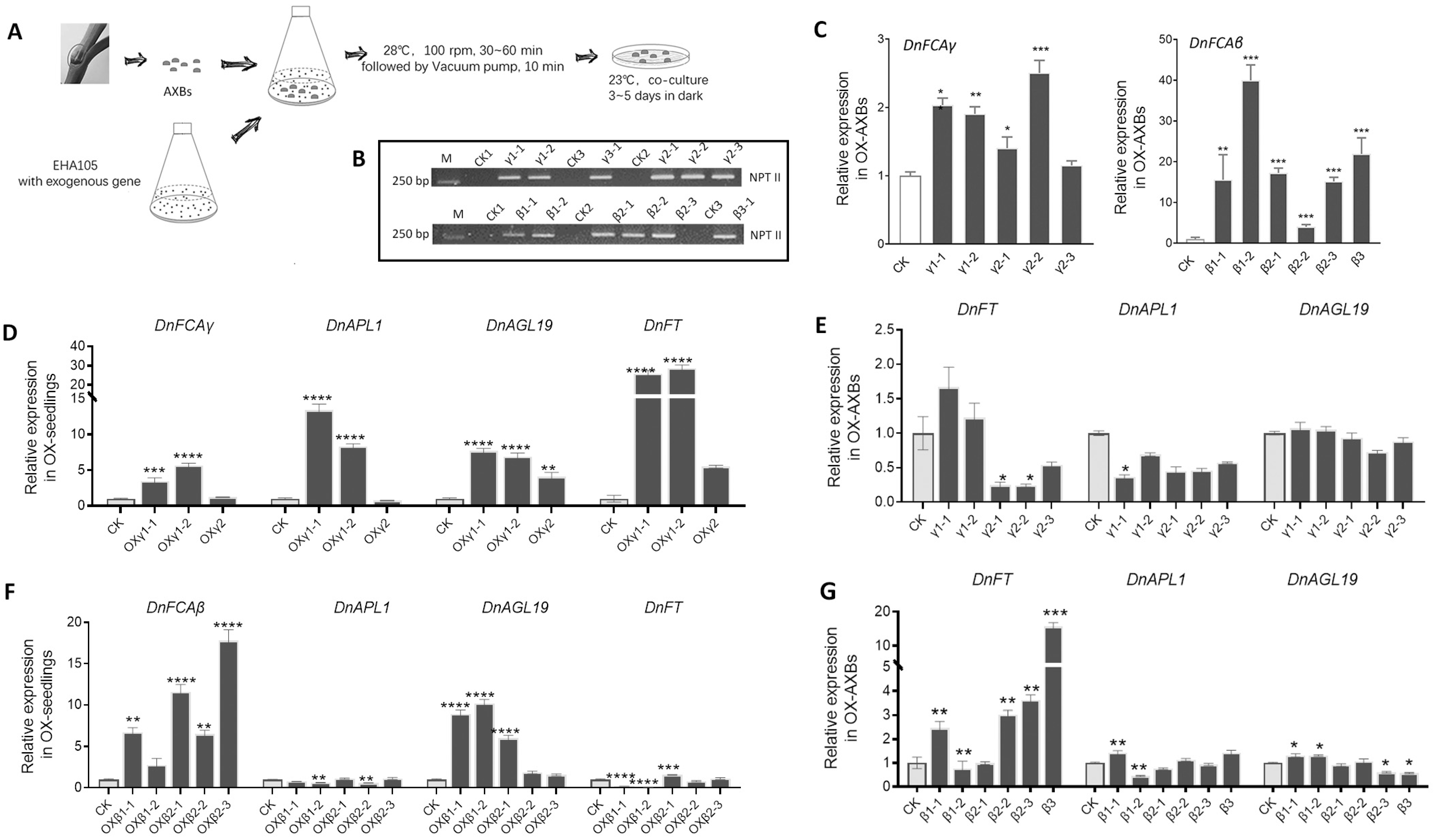
3.4. Overexpression of the DnFCA Isoform in D. nobile
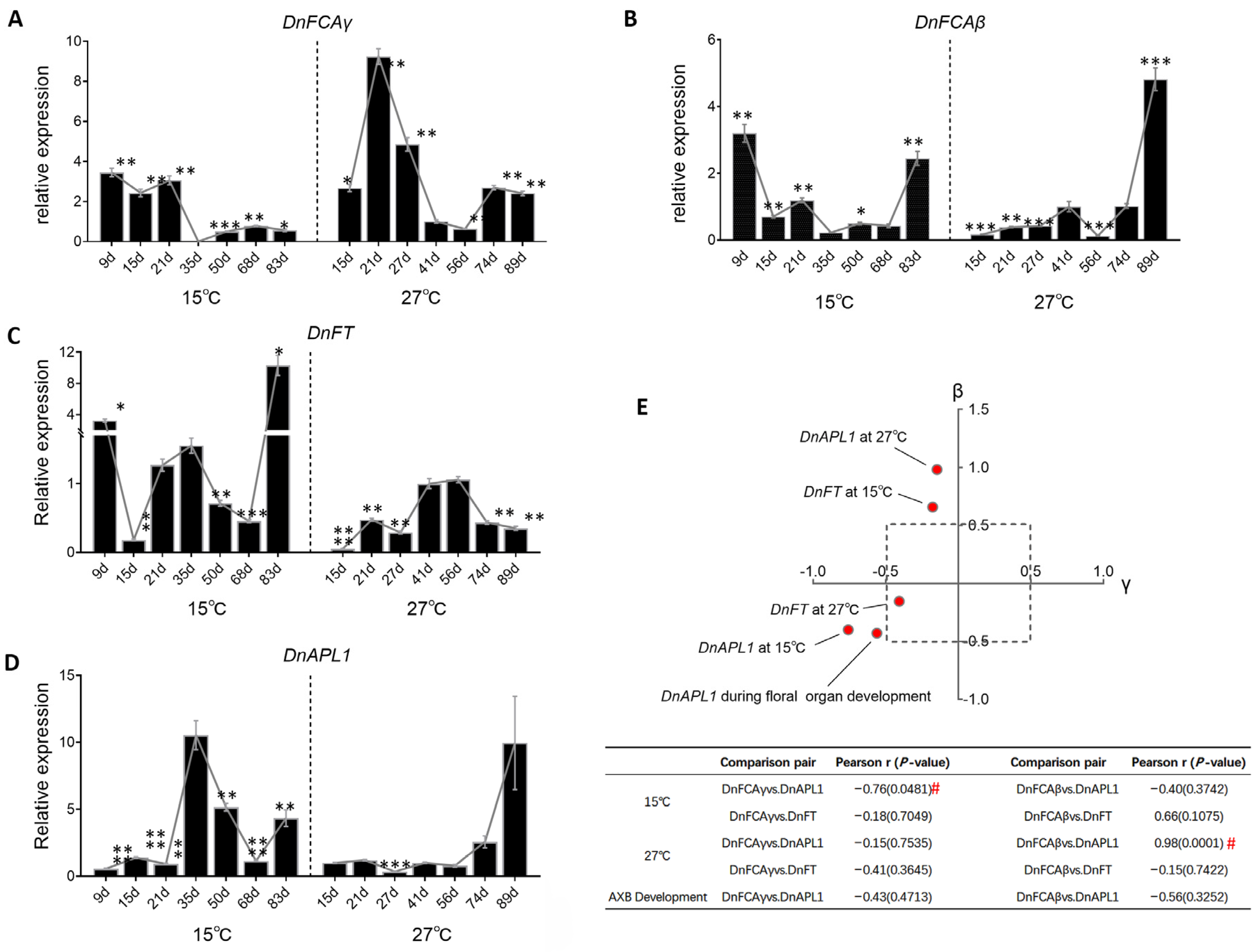
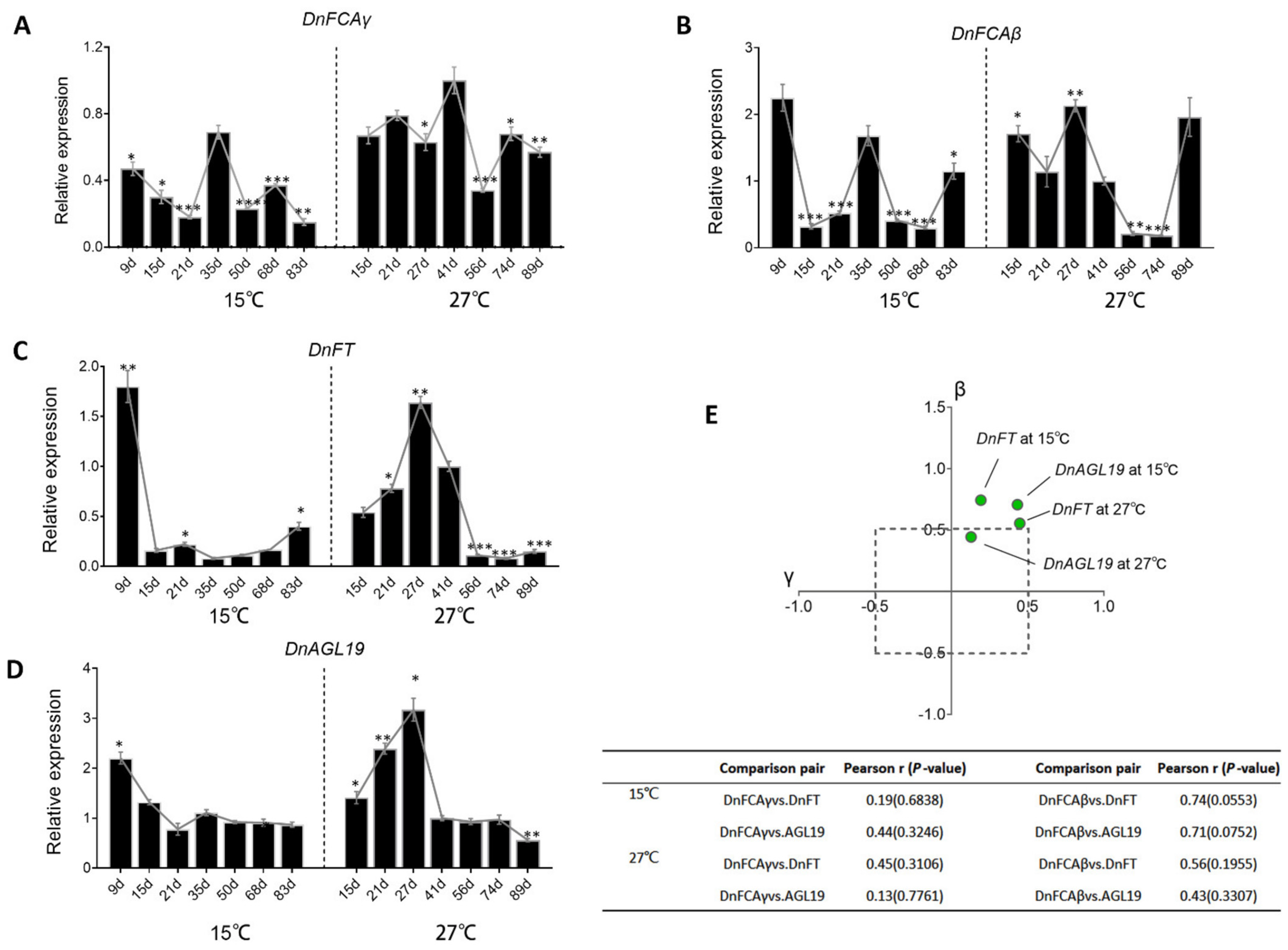
3.5. Temperature-Induced Accumulation of DnFCA Isoforms
4. Discussion
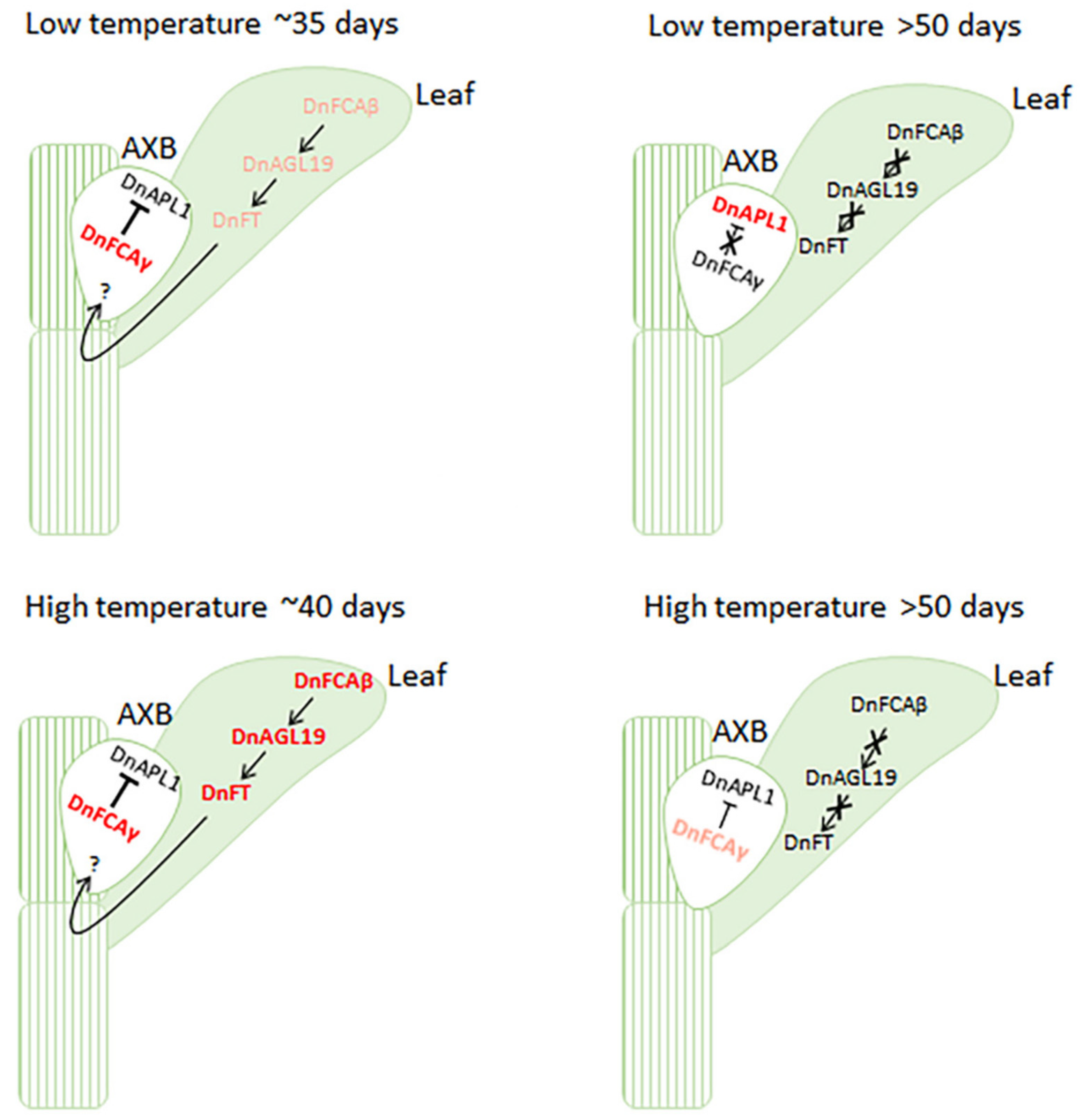
4.1. The DnFCAγ Protein Suppresses the Expression of DnAPL1 in AXBs
4.2. The DnFCAβ Protein Induces the Expression of DnAGL19 in Leaves
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yen, C.Y.; Starman, T.W.; Wang, Y.T.; Niu, G. Effects of cooling temperature and duration on flowering of the nobile Dendrobium orchid. HortScience 2008, 43, 1765–1769. [Google Scholar]
- Rotor, G.B. Daylength and temperature in relation to growth and flowering of orchids. Cornell Univ. Agric. Exp. Stn. Bull. 1952, 885, 3–47. [Google Scholar]
- Kosugi, K. Effects of soil moisture and low temperature upon the flower bud differentiation in the Dendrobium nobile. J. Jpn. Soc. Hortic. Sci. 1952, 21, 179–182. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Starman, T.W.; Wang, Y.T.; Niu, G. Vernalization Duration and Light Intensity Influence Flowering of Three Hybrid Nobile Dendrobium Cultivars. HortScience Horts 2011, 46, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Starman, T.W.; Wang, Y.T.; Niu, G.; Cothren, J.T. Deferring flowering of nobile Dendrobium hybrids by holding plants under low temperature after vernalization. Sci. Hortic. 2011, 130, 869–873. [Google Scholar] [CrossRef]
- Yeh, Y.T.; Shen, R.-S.; Huang, K.L.; Miyajima, I. Effect of Two-stage Vernalization and Temperature Treatment at the Stage of Floral Bud Development on Flowering in Nobile Dendrobium. J. Fac. Agr. Kyushu Univ. 2021, 66, 9. [Google Scholar]
- Boss, P.K.; Bastow, R.M.; Mylne, J.S.; Dean, C. Multiple pathways in the decision to flower: Enabling, promoting, and resetting. Plant Cell 2004, 16, S18–S31. [Google Scholar]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of Flowering Time: Interacting Pathways as a Basis for Diversity. Plant Cell 2002, 14, S111–S130. [Google Scholar] [CrossRef] [Green Version]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Ann. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar]
- Gregis, V.; Sessa, A.; Dorca-Fornell, C.; Kater, M.M. The Arabidopsis floral meristem identity genes AP1, AGL24 and SVP directly repress class B and C floral homeotic genes. Plant J. 2009, 60, 626–637. [Google Scholar]
- Irish, V.F.; Sussex, I.M. Function of the apetala-1 gene during Arabidopsis floral development. Plant Cell 1990, 2, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Ito, T.; Wellmer, F.; Meyerowitz, E.M. Repression of AGAMOUS-LIKE 24 is a crucial step in promoting flower development. Nat. Genet. 2004, 36, 157–161. [Google Scholar]
- Alejandra Mandel, M.; Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 1992, 360, 273–277. [Google Scholar]
- Kaufmann, K.; Wellmer, F.; Muiño, J.M.; Ferrier, T.; Wuest, S.E.; Kumar, V.; Serrano-Mislata, A.; Madueño, F.; Krajewski, P.; Meyerowitz, E.M.; et al. Orchestration of Floral Initiation by APETALA1. Science 2010, 328, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Regulation of the Arabidopsis floral homeotic gene APETALA1. Cell 1994, 76, 131–143. [Google Scholar]
- Wagner, D.; Sablowski, R.W.; Meyerowitz, E.M. Transcriptional activation of APETALA1 by LEAFY. Science 1999, 285, 582–584. [Google Scholar]
- Liljegren, S.J.; Gustafson-Brown, C.; Pinyopich, A.; Ditta, G.S.; Yanofsky, M.F. Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 specify meristem fate. Plant Cell 1999, 11, 1007–1018. [Google Scholar]
- Wigge, P.A.; Kim, M.C.; Jaeger, K.E.; Busch, W.; Schmid, M.; Lohmann, J.U.; Weigel, D. Integration of spatial and temporal information during floral induction in Arabidopsis. Science 2005, 309, 1056–1059. [Google Scholar]
- Kaufmann, K.; Nagasaki, M.; Jauregui, R. Modelling the molecular interactions in the flower developmental network of Arabidopsis thaliana. Silico Biol. 2010, 10, 125–143. [Google Scholar] [CrossRef]
- Page, T.; Macknight, R.; Yang, C.H.; Dean, C. Genetic interactions of the Arabidopsis flowering time gene FCA, with genes regulating floral initiation. Plant J. 1999, 17, 231–239. [Google Scholar]
- Murai, K.; Miyamae, M.; Kato, H.; Takumi, S.; Ogihara, Y. WAP1, a Wheat APETALA1 Homolog, Plays a Central Role in the Phase Transition from Vegetative to Reproductive Growth. Plant Cell Physiol. 2003, 44, 1255–1265. [Google Scholar] [CrossRef]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar]
- Fu, D.; Szűcs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Greenup, A.; Peacock, W.J.; Dennis, E.S.; Trevaskis, B. The molecular biology of seasonal flowering-responses in Arabidopsis and the cereals. Ann. Bot. 2009, 103, 1165–1172. [Google Scholar]
- Shimada, S.; Ogawa, T.; Kitagawa, S.; Suzuki, T.; Ikari, C.; Shitsukawa, N.; Abe, T.; Kawahigashi, H.; Kikuchi, R.; Handa, H. A genetic network of flowering-time genes in wheat leaves, in which an APETALA1/FRUITFULL-like gene, VRN1, is upstream of FLOWERING LOCUS T. Plant J. 2009, 58, 668–681. [Google Scholar]
- Macknight, R.; Bancroft, I.; Page, T.; Lister, C.; Schmidt, R.; Love, K.; Westphal, L.; Murphy, G.; Sherson, S.; Cobbett, C.; et al. FCA, a gene controlling flowering time in Arabidopsis, encodes a protein containing RNA-binding domains. Cell 1997, 89, 737–745. [Google Scholar]
- Lee, J.H.; Cho, Y.S.; Yoon, H.S.; Suh, M.C.; Moon, J.; Lee, I.; Weigel, D.; Yun, C.H.; Kim, J.K. Conservation and divergence of FCA function between Arabidopsis and rice. Plant Mol. Biol. 2005, 58, 823–838. [Google Scholar]
- Kumar, S.; Jiang, S.; Jami, S.K.; Hill, R.D. Cloning and characterization of barley caryopsis FCA. Physiol. Plant. 2011, 143, 93–106. [Google Scholar] [CrossRef]
- Macknight, R.; Duroux, M.; Laurie, R.; Dijkwel, P.; Simpson, G.; Dean, C. Functional significance of the alternative transcript processing of the Arabidopsis floral promoter FCA. Plant Cell 2002, 14, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Seo, P.J.; Ahn, J.H.; Park, C.M. Arabidopsis RNA-binding protein FCA regulates microRNA172 processing in thermosensory flowering. J. Biol. Chem. 2012, 287, 16007–16016. [Google Scholar]
- Rao, S.; Li, Y.; Chen, J. Combined Analysis of MicroRNAs and Target Genes Revealed miR156-SPLs and miR172-AP2 Are Involved in a Delayed Flowering Phenomenon after Chromosome Doubling in Black Goji (Lycium Ruthencium). Front. Genet. 2021, 12, 706930. [Google Scholar]
- Mathieu, J.; Yant, L.J.; Mürdter, F.; Küttner, F.; Schmid, M. Repression of flowering by the miR172 target SMZ. PLoS Biol. 2009, 7, e1000148. [Google Scholar]
- Swiezewski, S.; Liu, F.; Magusin, A.; Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature 2009, 462, 799–802. [Google Scholar]
- Liu, F.; Marquardt, S.; Lister, C.; Swiezewski, S.; Dean, C. Targeted 3′ Processing of Antisense Transcripts Triggers Arabidopsis FLC Chromatin Silencing. Science 2010, 327, 94–97. [Google Scholar] [CrossRef]
- Sawettalake, N.; Bunnag, S.; Wang, Y.; Shen, L.; Yu, H. DOAP1 promotes flowering in the orchid Dendrobium Chao Praya Smile. Front. Plant Sci. 2017, 8, 400. [Google Scholar]
- Chen, D.; Guo, B.; Hexige, S.; Zhang, T.; Shen, D.; Ming, F. SQUA-like genes in the orchid Phalaenopsis are expressed in both vegetative and reproductive tissues. Planta 2007, 226, 369–380. [Google Scholar]
- Chang, Y.Y.; Chiu, Y.F.; Wu, J.W.; Yang, C.H. Four orchid (Oncidium Gower Ramsey) AP1/AGL9-like MADS box genes show novel expression patterns and cause different effects on floral transition and formation in Arabidopsis thaliana. Plant Cell Physiol. 2009, 50, 1425–1438. [Google Scholar]
- Skipper, M.; Pedersen, K.B.; Johansen, L.B.; Frederiksen, S.; Irish, V.F.; Johansen, B.B. Identification and quantification of expression levels of three FRUITFULL-like MADS-box genes from the orchid Dendrobium thyrsiflorum (Reichb. f.). Plant Sci. 2005, 169, 579–586. [Google Scholar]
- Yu, H.; Goh, C.J. Identification and characterization of three orchid MADS-box genes of the AP1/AGL9 subfamily during floral transition. Plant Physiol. 2000, 123, 1325–1336. [Google Scholar]
- Liang, S.; Ye, Q.S.; Li, R.H.; Leng, J.Y.; Li, M.R.; Wang, X.J.; Li, H.Q. Transcriptional Regulations on the Low-Temperature-Induced Floral Transition in an Orchidaceae Species, Dendrobium nobile: An Expressed Sequence Tags Analysis. Comp. Funct. Genom. 2012, 2012, 757801. [Google Scholar] [CrossRef] [Green Version]
- Deng, N.M.; Gao, L.; Wang, C.; Deng, S.L.; Shen, W.J.; Gao, C.; Liang, S. Isolation and functional characterization of DnAPL1, an AP1/FUL subfamily member from Dendrobium nobile. Plant Physiol. J. 2020, 56, 9. [Google Scholar]
- Liu, X.R.; Pan, T.; Liang, W.Q.; Gao, L.; Wang, X.J.; Li, H.Q.; Liang, S. Overexpression of an orchid (Dendrobium nobile) SOC1/TM3-like ortholog, DnAGL19, in Arabidopsis regulates HOS1-FT expression. Front. Plant Sci. 2016, 7, 99. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Logemann, E.; Birkenbihl, R.P.; Ülker, B.; Somssich, I.E. An improved method for preparing Agrobacterium cells that simplifies the Arabidopsis transformation protocol. Plant Methods 2006, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Görlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Vervliet, G.; Holsters, M.; Teuchy, H.; Van Montagu, M.; Schell, J. Characterization of different plaque-forming and defective temperate phages in Agrobacterium. J. Gen. Virol. 1975, 26, 33–48. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Xu, Q.; Niu, S.C.; Li, K.L.; Zheng, P.J.; Zhang, X.J.; Jia, Y.; Liu, Y.; Niu, Y.X.; Yu, L.H.; Chen, D.F.; et al. Chromosome-Scale Assembly of the Dendrobium nobile Genome Provides Insights Into the Molecular Mechanism of the Biosynthesis of the Medicinal Active Ingredient of Dendrobium. Front. Genet. 2022, 13, 844622. [Google Scholar] [CrossRef]
- Li, R.; Wang, A.; Sun, S.; Liang, S.; Wang, X.; Ye, Q.; Li, H. Functional characterization of FT and MFT ortholog genes in orchid (Dendrobium nobile Lindl) that regulate the vegetative to reproductive transition in Arabidopsis. Plant Cell Tissue Organ Cult. (PCTOC) 2012, 111, 143–151. [Google Scholar]
- Goh, C.J.; Arditti, J. Orchidaceae. In Handbook of Flowering; Halevy, A.H., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 309–336. [Google Scholar]
- Chuck, G.; Hake, S. Regulation of developmental transitions. Curr. Opin. Plant Biol. 2005, 8, 67–70. [Google Scholar] [CrossRef]
- Lai, X.; Blanc-Mathieu, R.; Grandvuillemin, L.; Huang, Y.; Stigliani, A.; Lucas, J.; Thévenon, E.; Loue-Manifel, J.; Turchi, L.; Daher, H. The LEAFY floral regulator displays pioneer transcription factor properties. Mol. Plant 2021, 14, 829–837. [Google Scholar]
- Wang, R.; Albani, M.C.; Vincent, C.; Bergonzi, S.; Luan, M.; Bai, Y.; Kiefer, C.; Castillo, R.; Coupland, G. Aa TFL1 Confers an Age-Dependent Response to Vernalization in Perennial Arabis alpina. Plant Cell 2011, 23, 1307–1321. [Google Scholar] [CrossRef] [Green Version]
- Bergonzi, S.; Albani, M.C.; Ver Loren van Themaat, E.; Nordström, K.J.; Wang, R.; Schneeberger, K.; Moerland, P.D.; Coupland, G. Mechanisms of age-dependent response to winter temperature in perennial flowering of Arabis alpina. Science 2013, 340, 1094–1097. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, T.; Deng, N.-M.; Guo, W.-X.; Wan, M.-Z.; Zheng, Y.-T.; Chen, S.-Y.; Liu, C.-L.; Li, H.-B.; Liang, S. DnFCA Isoforms Cooperatively Regulate Temperature-Related Flowering in Dendrobium nobile. Biology 2023, 12, 331. https://doi.org/10.3390/biology12020331
Pan T, Deng N-M, Guo W-X, Wan M-Z, Zheng Y-T, Chen S-Y, Liu C-L, Li H-B, Liang S. DnFCA Isoforms Cooperatively Regulate Temperature-Related Flowering in Dendrobium nobile. Biology. 2023; 12(2):331. https://doi.org/10.3390/biology12020331
Chicago/Turabian StylePan, Ting, Ning-Meng Deng, Wu-Xia Guo, Min-Zhen Wan, Yan-Tong Zheng, Song-Yi Chen, Chuan-Liang Liu, Hong-Bo Li, and Shan Liang. 2023. "DnFCA Isoforms Cooperatively Regulate Temperature-Related Flowering in Dendrobium nobile" Biology 12, no. 2: 331. https://doi.org/10.3390/biology12020331