

Ligand Binding Properties of Odorant-Binding Protein OBP5 from Mus musculus
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. mOBP5 Production and Purification
2.2. Recombinant mOBP5 Characterization
2.3. Fluorescence-Based Binding Assay
2.4. Isothermal Titration Microcalorimetry (ITC)
2.5. Molecular Modeling and Molecular Docking
3. Results
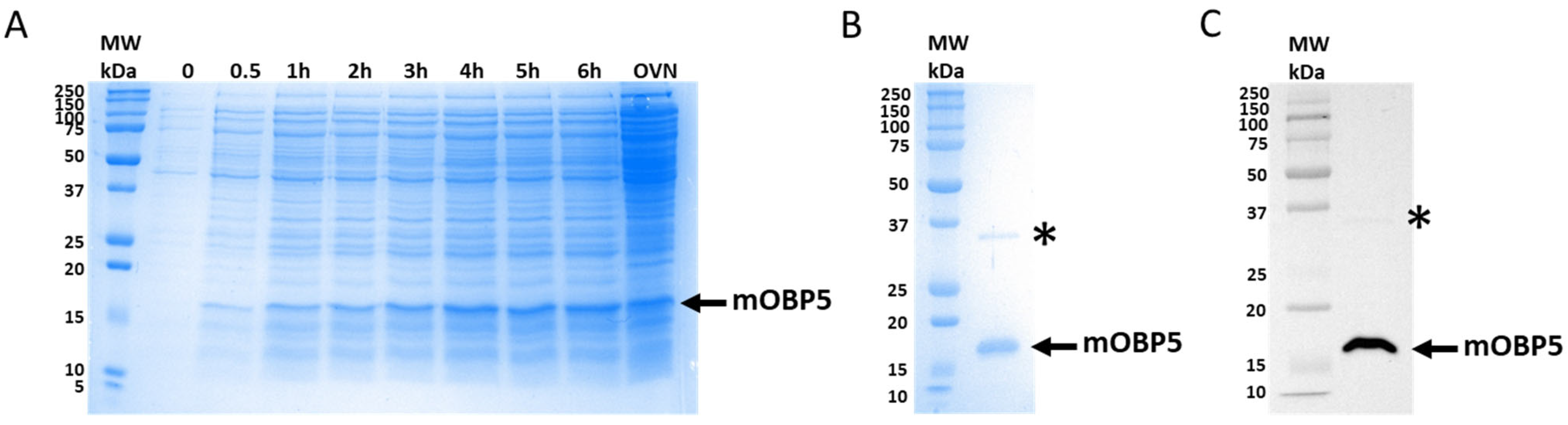
3.1. Expression and Purification of mOBP5
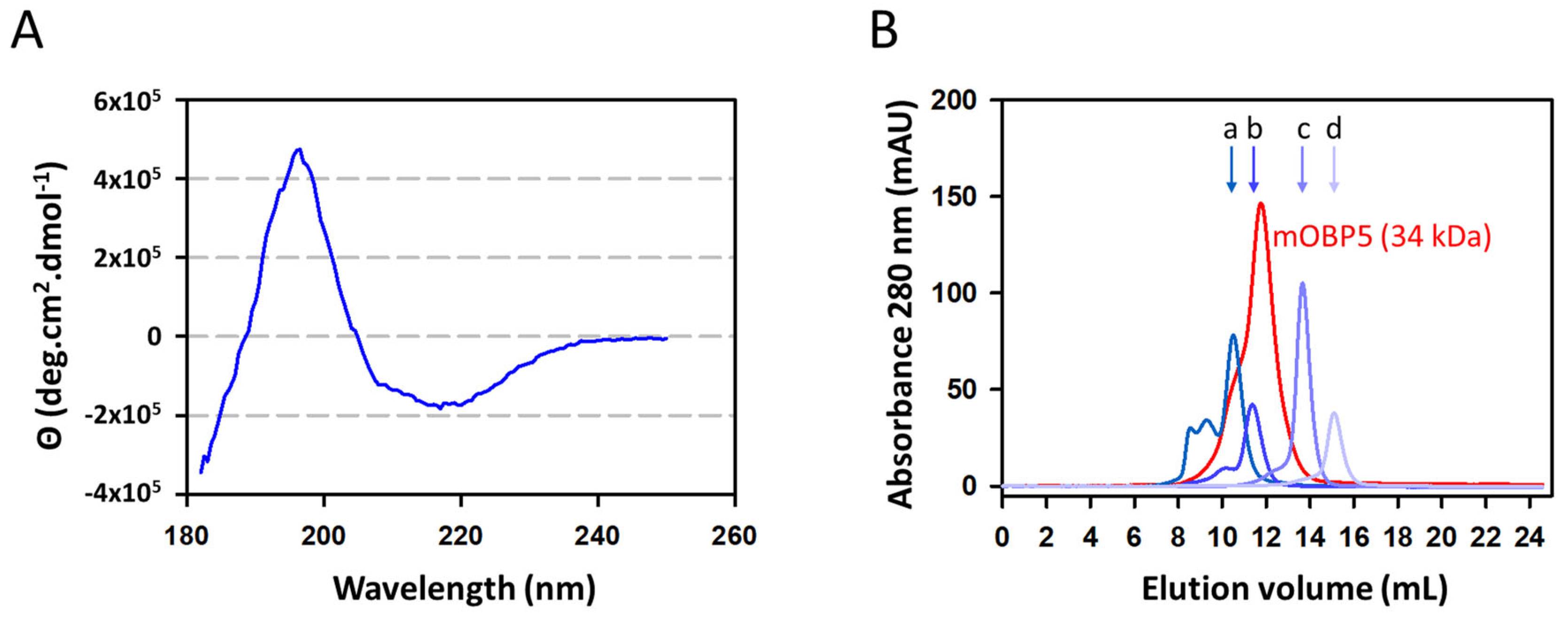
3.2. Characterization of mOBP5
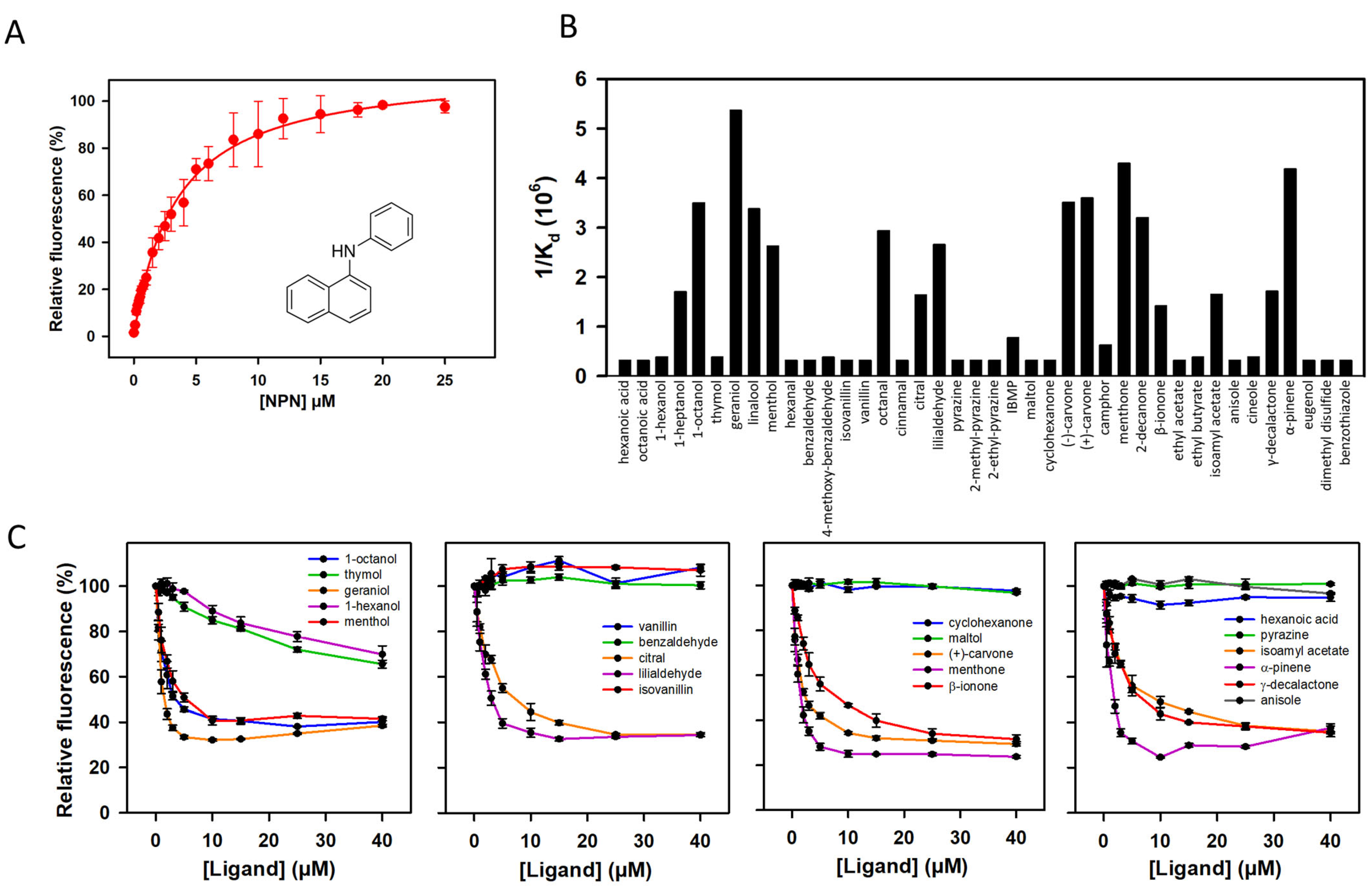
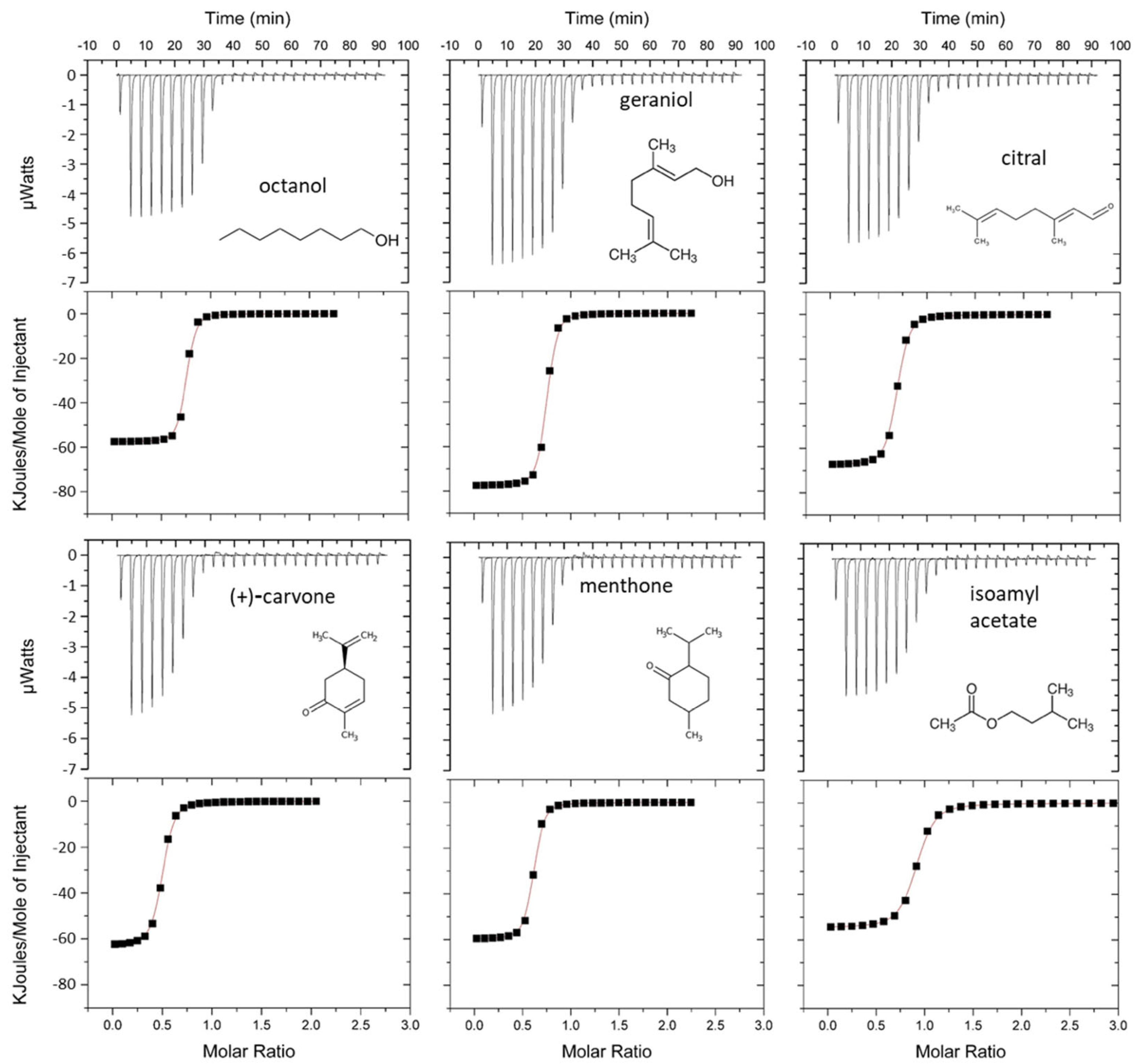
3.3. Ligand Binding Properties of mOBP5
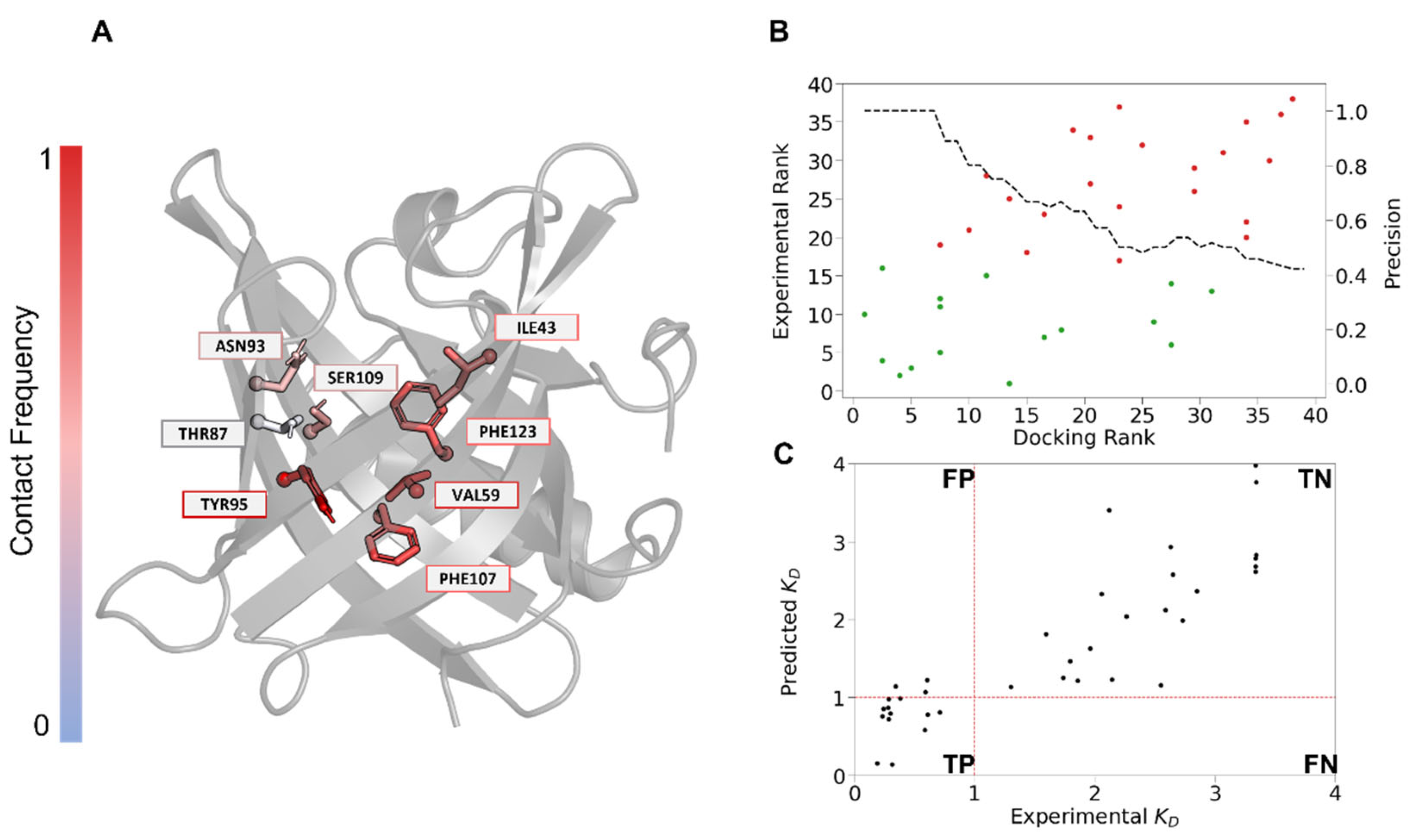
3.4. Docking Reveals Key Residues for mOBP5 Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heydel, J.M.; Coelho, A.; Thiebaud, N.; Legendre, A.; Le Bon, A.M.; Faure, P.; Neiers, F.; Artur, Y.; Golebiowski, J.; Briand, L. Odorant-binding proteins and xenobiotic metabolizing enzymes: Implications in olfactory perireceptor events. Anat. Rec. 2013, 296, 1333–1345. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P. The role of perireceptor events in vertebrate olfaction. Cell. Mol. Life Sci. CMLS 2001, 58, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Knoll, W. Odorant-binding proteins of mammals. Biol. Rev. 2022, 97, 20–44. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Flisi, S.; Careri, M.; Riboni, N.; Resimini, S.; Sala, A.; Conti, V.; Mattarozzi, M.; Taddei, S.; Spadini, C.; et al. Vertebrate odorant binding proteins as antimicrobial humoral components of innate immunity for pathogenic microorganisms. PLoS ONE 2019, 14, e0213545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flower, D.R. Beyond the superfamily: The lipocalin receptors. Biochim. Biophys. Acta 2000, 1482, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.; Spinelli, S.; Ramoni, R.; Grolli, S.; Pelosi, P.; Cambillau, C.; Tegoni, M. Complexes of porcine odorant binding protein with odorant molecules belonging to different chemical classes. J. Mol. Biol. 2000, 300, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Logan, J.G.; Birkett, M.A.; Clark, S.J.; Powers, S.; Seal, N.J.; Wadhams, L.J.; Mordue Luntz, A.J.; Pickett, J.A. Identification of human-derived volatile chemicals that interfere with attraction of Aedes aegypti mosquitoes. J. Chem. Ecol. 2008, 34, 308–322. [Google Scholar] [CrossRef]
- Shahan, K.; Denaro, M.; Gilmartin, M.; Shi, Y.; Derman, E. Expression of six mouse major urinary protein genes in the mammary, parotid, sublingual, submaxillary, and lachrymal glands and in the liver. Mol. Cell. Biol. 1987, 7, 1947–1954. [Google Scholar] [CrossRef]
- Utsumi, M.; Ohno, K.; Kawasaki, Y.; Tamura, M.; Kubo, T.; Tohyama, M. Expression of major urinary protein genes in the nasal glands associated with general olfaction. J. Neurobiol. 1999, 39, 227–236. [Google Scholar] [CrossRef]
- Briand, L.; Eloit, C.; Nespoulous, C.; Bézirard, V.; Huet, J.C.; Henry, C.; Blon, F.; Trotier, D.; Pernollet, J.C. Evidence of an odorant-binding protein in the human olfactory mucus: Location, structural characterization, and odorant-binding properties. Biochemistry 2002, 41, 7241–7252. [Google Scholar] [CrossRef]
- Cann, P.; Chabi, M.; Delsart, A.; Le Danvic, C.; Saliou, J.M.; Chasles, M.; Keller, M.; Nagnan-Le Meillour, P. The olfactory secretome varies according to season in female sheep and goat. BMC Genom. 2019, 20, 794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazar, J.; Greenwood, D.R.; Rasmussen, L.E.; Prestwich, G.D. Molecular and functional characterization of an odorant binding protein of the Asian elephant, Elephas maximus: Implications for the role of lipocalins in mammalian olfaction. Biochemistry 2002, 41, 11786–11794. [Google Scholar] [CrossRef] [PubMed]
- Tegoni, M.; Pelosi, P.; Vincent, F.; Spinelli, S.; Campanacci, V.; Grolli, S.; Ramoni, R.; Cambillau, C. Mammalian odorant binding proteins. Biochim. Biophys. Acta 2000, 1482, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Arena, S.; Spinelli, S.; Liu, D. Reverse chemical ecology: Olfactory proteins from the giant panda and their interactions with putative pheromones and bamboo volatiles. Proc. Natl. Acad. Sci. USA 2017, 114, E9802–E9810. [Google Scholar] [CrossRef] [Green Version]
- Löbel, D.; Jacob, M.; Völkner, M.; Breer, H. Odorants of different chemical classes interact with distinct odorant binding protein subtypes. Chem. Senses 2002, 27, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Löbel, D.; Marchese, S.; Krieger, J.; Pelosi, P.; Heinz, B. Subtypes of odorant-binding proteins—Heterologous expression and ligand binding. Eur. J. Biochem. 1998, 254, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Löbel, D.; Strotmann, J.; Jacob, M.; Breer, H. Identification of a third rat odorant-binding protein (OBP3). Chem. Senses 2001, 26, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Nagnan-Le Meillour, P.; Vercoutter-Edouart, A.S.; Hilliou, F.; Le Danvic, C.; Lévy, F. Proteomic Analysis of Pig (Sus scrofa) Olfactory Soluble Proteome Reveals O-Linked-N-Acetylglucosaminylation of Secreted Odorant-Binding Proteins. Front. Endocrinol. 2014, 5, 202. [Google Scholar] [CrossRef] [Green Version]
- Scaloni, A.; Paolini, S.; Brandazza, A.; Fantacci, M.; Bottiglieri, C.; Marchese, S.; Navarrini, A.; Fini, C.; Ferrara, L.; Pelosi, P. Purification, cloning and characterisation of odorant- and pheromone-binding proteins from pig nasal epithelium. Cell. Mol. Life Sci. CMLS 2001, 58, 823–834. [Google Scholar] [CrossRef]
- Brimau, F.; Cornard, J.P.; Le Danvic, C.; Lagant, P.; Vergoten, G.; Grebert, D.; Pajot, E.; Nagnan-Le Meillour, P. Binding specificity of recombinant odorant-binding protein isoforms is driven by phosphorylation. J. Chem. Ecol. 2010, 36, 801–813. [Google Scholar] [CrossRef]
- Ibarra-Soria, X.; Levitin, M.O.; Saraiva, L.R.; Logan, D.W. The olfactory transcriptomes of mice. PLoS Genet. 2014, 10, e1004593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuntová, B.; Stopková, R.; Stopka, P. Transcriptomic and Proteomic Profiling Revealed High Proportions of Odorant Binding and Antimicrobial Defense Proteins in Olfactory Tissues of the House Mouse. Front. Genet. 2018, 9, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pes, D.; Mameli, M.; Andreini, I.; Krieger, J.; Weber, M.; Breer, H.; Pelosi, P. Cloning and expression of odorant-binding proteins Ia and Ib from mouse nasal tissue. Gene 1998, 212, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Pes, D.; Pelosi, P. Odorant-binding proteins of the mouse. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 112, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Shiao, M.S.; Chang, A.Y.; Liao, B.Y.; Ching, Y.H.; Lu, M.Y.; Chen, S.M.; Li, W.H. Transcriptomes of mouse olfactory epithelium reveal sexual differences in odorant detection. Genome Biol. Evol. 2012, 4, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryche, B.; Frétaud, M.; Saint-Albin Deliot, A.; Galloux, M.; Sedano, L.; Langevin, C.; Descamps, D.; Rameix-Welti, M.A.; Eléouët, J.F.; Le Goffic, R. Respiratory syncytial virus tropism for olfactory sensory neurons in mice. J. Neurochem. 2020, 155, 137–153. [Google Scholar] [CrossRef]
- Strotmann, J.; Breer, H. Internalization of odorant-binding proteins into the mouse olfactory epithelium. Histochem. Cell Biol. 2011, 136, 357–369. [Google Scholar] [CrossRef]
- Stopková, R.; Otčenášková, T.; Matějková, T.; Kuntová, B.; Stopka, P. Biological Roles of Lipocalins in Chemical Communication, Reproduction, and Regulation of Microbiota. Front. Physiol. 2021, 12, 740006. [Google Scholar] [CrossRef]
- Stopkova, R.; Klempt, P.; Kuntova, B.; Stopka, P. On the tear proteome of the house mouse (Mus musculus musculus) in relation to chemical signalling. PeerJ 2017, 5, e3541. [Google Scholar] [CrossRef] [Green Version]
- Zaremska, V.; Tan, J.; Lim, S.; Knoll, W.; Pelosi, P. Isoleucine residues determine chiral discrimination of odorant-binding protein. Chem. Eur. J. 2020, 26, 8720–8724. [Google Scholar] [CrossRef]
- Zhu, J.; Zaremska, V.; D’Onofrio, C.; Knoll, W.; Pelosi, P. Site-directed mutagenesis of odorant-binding proteins. Methods Enzymol. 2020, 642, 301–324. [Google Scholar] [CrossRef]
- Gonçalves, F.; Ribeiro, A. Biotechnological applications of mammalian odorant-binding proteins. Crit. Rev. Biotechnol. 2021, 41, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Barou, E.; Sigoillot, M.; Bouvet, M.; Briand, L.; Meunier-Prest, R. Electrochemical detection of the 2-isobutyl-3-methoxypyrazine model odorant based on odorant-binding proteins: The proof of concept. Bioelectrochemistry 2015, 101, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Capo, A.; Pennacchio, A.; Varriale, A.; D’Auria, S.; Staiano, M. The porcine odorant-binding protein as molecular probe for benzene detection. PLoS ONE 2018, 13, e0202630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurot, C.; Brenet, S.; Buhot, A.; Barou, E.; Belloir, C.; Briand, L.; Hou, Y. Highly sensitive olfactory biosensors for the detection of volatile organic compounds by surface plasmon resonance imaging. Biosens. Bioelectron. 2019, 123, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Larisika, M.; Kotlowski, C.; Steininger, C.; Mastrogiacomo, R.; Pelosi, P.; Schütz, S.; Peteu, S.F.; Kleber, C.; Reiner-Rozman, C.; Nowak, C.; et al. Electronic olfactory sensor based on A. mellifera Odorant-Binding Protein 14 on a reduced graphene Oxide Field-Effect Transistor. Angew. Chem. Int. Ed. 2015, 54, 13245–13248. [Google Scholar] [CrossRef]
- Mulla, M.Y.; Tuccori, E.; Magliulo, M.; Lattanzi, G.; Palazzo, G.; Persaud, K.; Torsi, L. Capacitance-modulated transistor detects odorant binding protein chiral interactions. Nat. Commun. 2015, 6, 6010. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Zhu, J.; Knoll, W. Odorant-Binding Proteins as sensing elements for odour monitoring. Sensors 2018, 18, 3248. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.; Matamá, T.; Azoia, N.G.; Mansilha, C.; Casal, M.; Cavaco-Paulo, A. Odorant binding proteins: A biotechnological tool for odour control. Appl. Microbiol. Biotechnol. 2014, 98, 3629–3638. [Google Scholar] [CrossRef]
- Saraiva, L.R.; Kondoh, K.; Ye, X.; Yoon, K.H.; Hernandez, M.; Buck, L.B. Combinatorial effects of odorants on mouse behavior. Proc. Natl. Acad. Sci. USA 2016, 113, E3300–E3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brulé, M.; Glaz, M.; Belloir, C.; Poirier, N.; Moitrier, L.; Neiers, F.; Briand, L. Bacterial expression and purification of vertebrate odorant-binding proteins. Methods Enzymol. 2020, 642, 125–150. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Olsson, M.H.M.; Søndergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent treatment of internal and surface residues in empirical pKa predictions. J. Chem. Theory Comput. 2011, 7, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Søndergaard, C.R.; Olsson, M.H.M.; Rostkowski, M.; Jensen, J.H. Improved treatment of ligands and coupling effects in empirical calculation and rationalization of pKa values. J. Chem. Theory Comput. 2011, 7, 2284–2295. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ropp, P.J.; Spiegel, J.O.; Walker, J.L.; Green, H.; Morales, G.A.; Milliken, K.A.; Ringe, J.J.; Durrant, J.D. Gypsum-DL: An open-source program for preparing small-molecule libraries for structure-based virtual screening. J. Cheminform. 2019, 11, 34. [Google Scholar] [CrossRef]
- Greg, L. rdkit/rdkit: 2022_09_1 (Q3 2022) Release (Release_2022_09_1). Zenodo. 2022. Available online: https://zenodo.org/record/7235579#.Y5h-PXbMJPY (accessed on 1 July 2022).
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Nagy, G.; Igaev, M. SESCA: Predicting circular dichroism spectra from protein molecular structures. J. Chem. Theory Comput. 2019, 15, 5087–5102. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Q.; Huang, H.; Huang, W. A crowdsourcing open platform for literature curation in UniProt. PLoS Biol. 2021, 19, e3001464. [Google Scholar] [CrossRef] [PubMed]
- Rozewicki, J.; Li, S.; Amada, K.M.; Standley, D.M.; Katoh, K. MAFFT-DASH: Integrated protein sequence and structural alignment. Nucleic Acids Res. 2019, 47, W5–W10. [Google Scholar] [CrossRef] [PubMed]
- Ramoni, R.; Spinelli, S.; Grolli, S.; Conti, V.; Merli, E.; Cambillau, C.; Tegoni, M. Deswapping bovine odorant binding protein. Biochim. Biophys. Acta 2008, 1784, 651–657. [Google Scholar] [CrossRef]
- Ramoni, R.; Vincent, F.; Grolli, S.; Conti, V.; Malosse, C.; Boyer, F.D.; Nagnan-Le Meillour, P.; Spinelli, S.; Cambillau, C.; Tegoni, M. The insect attractant 1-octen-3-ol is the natural ligand of bovine odorant-binding protein. J. Biol. Chem. 2001, 276, 7150–7155. [Google Scholar] [CrossRef] [Green Version]
- Borysik, A.J.; Briand, L.; Taylor, A.J.; Scott, D.J. Rapid odorant release in mammalian odour binding proteins facilitates their temporal coupling to odorant signals. J. Mol. Biol. 2010, 404, 372–380. [Google Scholar] [CrossRef]
- Burova, T.V.; Choiset, Y.; Jankowski, C.K.; Haertlé, T. Conformational stability and binding properties of porcine odorant binding protein. Biochemistry 1999, 38, 15043–15051. [Google Scholar] [CrossRef]
- Nespoulous, C.; Briand, L.; Delage, M.M.; Tran, V.; Pernollet, J.C. Odorant binding and conformational changes of a rat odorant-binding protein. Chem. Senses 2004, 29, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Yabuki, M.; Scott, D.J.; Briand, L.; Taylor, A.J. Dynamics of odorant binding to thin aqueous films of rat-OBP3. Chem. Senses 2011, 36, 659–671. [Google Scholar] [CrossRef] [Green Version]
- Slater, O.; Kontoyianni, M. The compromise of virtual screening and its impact on drug discovery. Expert Opin. Drug Discov. 2019, 14, 619–637. [Google Scholar] [CrossRef]
- Hoffmann, W.; Langenhan, J.; Huhmann, S.; Moschner, J.; Chang, R.; Accorsi, M.; Seo, J.; Rademann, J.; Meijer, G.; Koksch, B.; et al. An intrinsic hydrophobicity scale for amino acids and its application to fluorinated compounds. Angew. Chem. Int. Ed. 2019, 58, 8216–8220. [Google Scholar] [CrossRef] [PubMed]
- Portman, K.L.; Long, J.; Carr, S.; Briand, L.; Winzor, D.J.; Searle, M.S.; Scott, D.J. Enthalpy/Entropy compensation effects from cavity desolvation underpin broad ligand binding sSelectivity for rat Odorant Binding Protein 3. Biochemistry 2014, 53, 2371–2379. [Google Scholar] [CrossRef] [PubMed]
- Bianchet, M.A.; Bains, G.; Pelosi, P.; Pevsner, J.; Snyder, S.H.; Monaco, H.L.; Amzel, L.M. The three-dimensional structure of bovine odorant binding protein and its mechanism of odor recognition. Nat. Struct. Biol. 1996, 3, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Pevsner, J.; Trifiletti, R.R.; Strittmatter, S.M.; Snyder, S.H. Isolation and characterization of an olfactory receptor protein for odorant pyrazines. Proc. Natl. Acad. Sci. USA 1985, 82, 3050–3054. [Google Scholar] [CrossRef] [Green Version]
- Osada, K.; Kurihara, K.; Izumi, H.; Kashiwayanagi, M. Pyrazine analogues are active components of wolf urine that induce avoidance and freezing behaviours in mice. PLoS ONE 2013, 8, e61753. [Google Scholar] [CrossRef] [Green Version]
- Tegoni, M.; Ramoni, R.; Bignetti, E.; Spinelli, S.; Cambillau, C. Domain swapping creates a third putative combining site in bovine odorant binding protein dimer. Nat. Struct. Biol. 1996, 3, 863–867. [Google Scholar] [CrossRef]
- White, S.A.; Briand, L.; Scott, D.J.; Borysik, A.J. Structure of rat odorant-binding protein OBP1 at 1.6 A resolution. Acta Crystallogr. Sect. D 2009, 65, 403–410. [Google Scholar] [CrossRef]
- Spinelli, S.; Ramoni, R.; Grolli, S.; Bonicel, J.; Cambillau, C.; Tegoni, M. The Structure of the monomeric porcine Odorant Binding Protein sheds light on the domain swapping mechanism. Biochemistry 1998, 37, 7913–7918. [Google Scholar] [CrossRef]
- Schiefner, A.; Freier, R.; Eichinger, A.; Skerra, A. Crystal structure of the human odorant binding protein, OBPIIa. Proteins Struct. Funct. Bioinform. 2015, 83, 1180–1184. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | Interaction | Frequency | Conservation | Conservation Type |
|---|---|---|---|---|
| TYR95 | Hydrophobic | 99% | 3% | Hydrophobic |
| VAL59 | Hydrophobic | 91% | 11% | Hydrophobic |
| PHE107 | Hydrophobic | 85% | 61% | Aromatic |
| PHE123 | Hydrophobic | 80% | 3% | Polar |
| ILE43 | Hydrophobic | 77% | 5% | - |
| ASN93 | H-Bond | 67% | 11% | Polar |
| SER109 | H-Bond | 65% | 5% | Small |
| THR87 | H-Bond | 48% | 34% | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moitrier, L.; Belloir, C.; Lalis, M.; Hou, Y.; Topin, J.; Briand, L. Ligand Binding Properties of Odorant-Binding Protein OBP5 from Mus musculus. Biology 2023, 12, 2. https://doi.org/10.3390/biology12010002
Moitrier L, Belloir C, Lalis M, Hou Y, Topin J, Briand L. Ligand Binding Properties of Odorant-Binding Protein OBP5 from Mus musculus. Biology. 2023; 12(1):2. https://doi.org/10.3390/biology12010002
Chicago/Turabian StyleMoitrier, Lucie, Christine Belloir, Maxence Lalis, Yanxia Hou, Jérémie Topin, and Loïc Briand. 2023. "Ligand Binding Properties of Odorant-Binding Protein OBP5 from Mus musculus" Biology 12, no. 1: 2. https://doi.org/10.3390/biology12010002